
Effect of Long-Term Nitrogen and Phosphorus Additions on Understory Plant Nutrients in a Primary Tropical Forest



Abstract
:1. Introduction
2. Methods
2.1. Study Site
2.2. Experimental Design
2.3. Sampling and Analysis of Plants
2.4. Statistical Analyses
3. Results
3.1. Responses of Soil Properties to N or P Addition
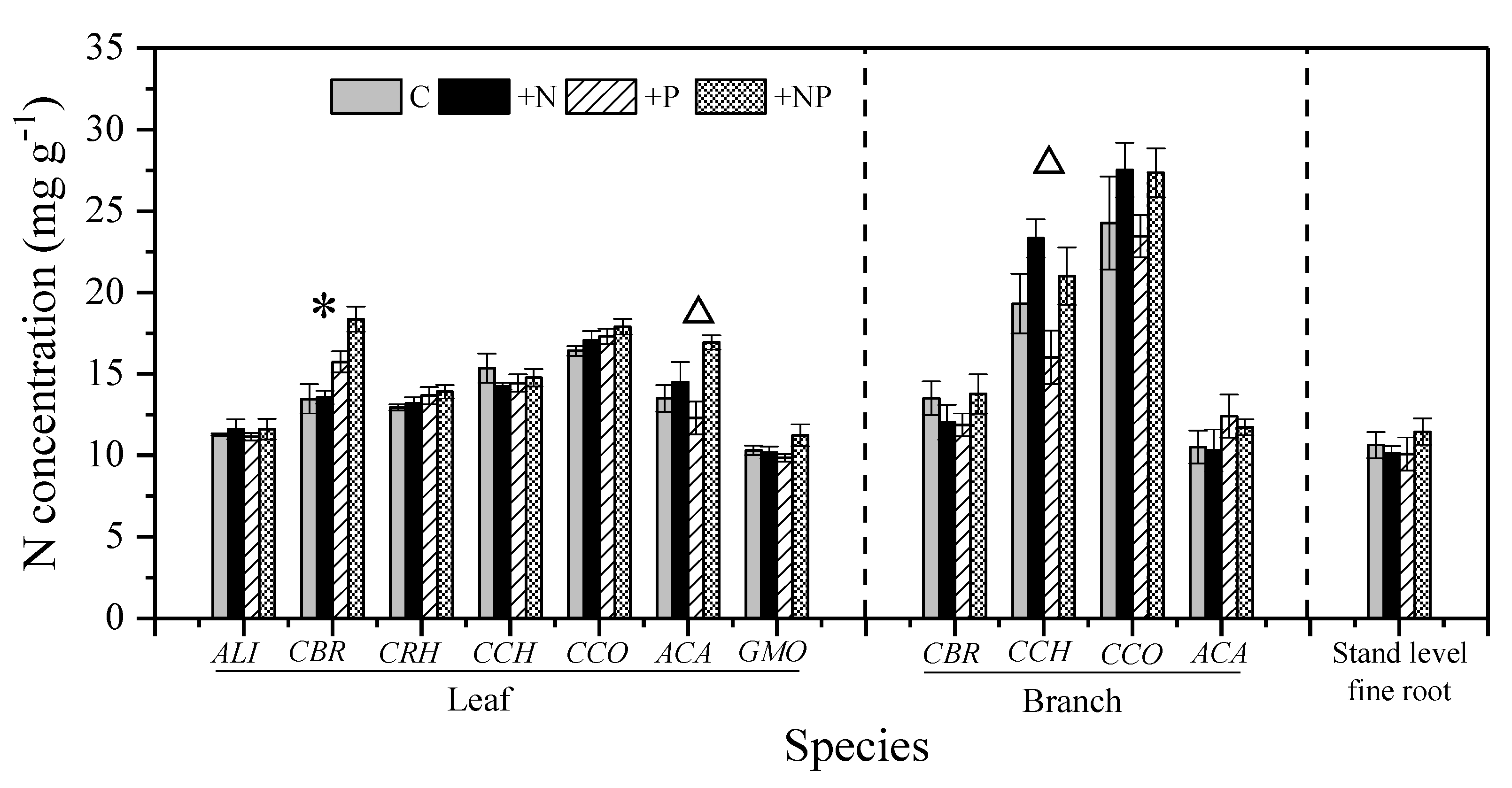
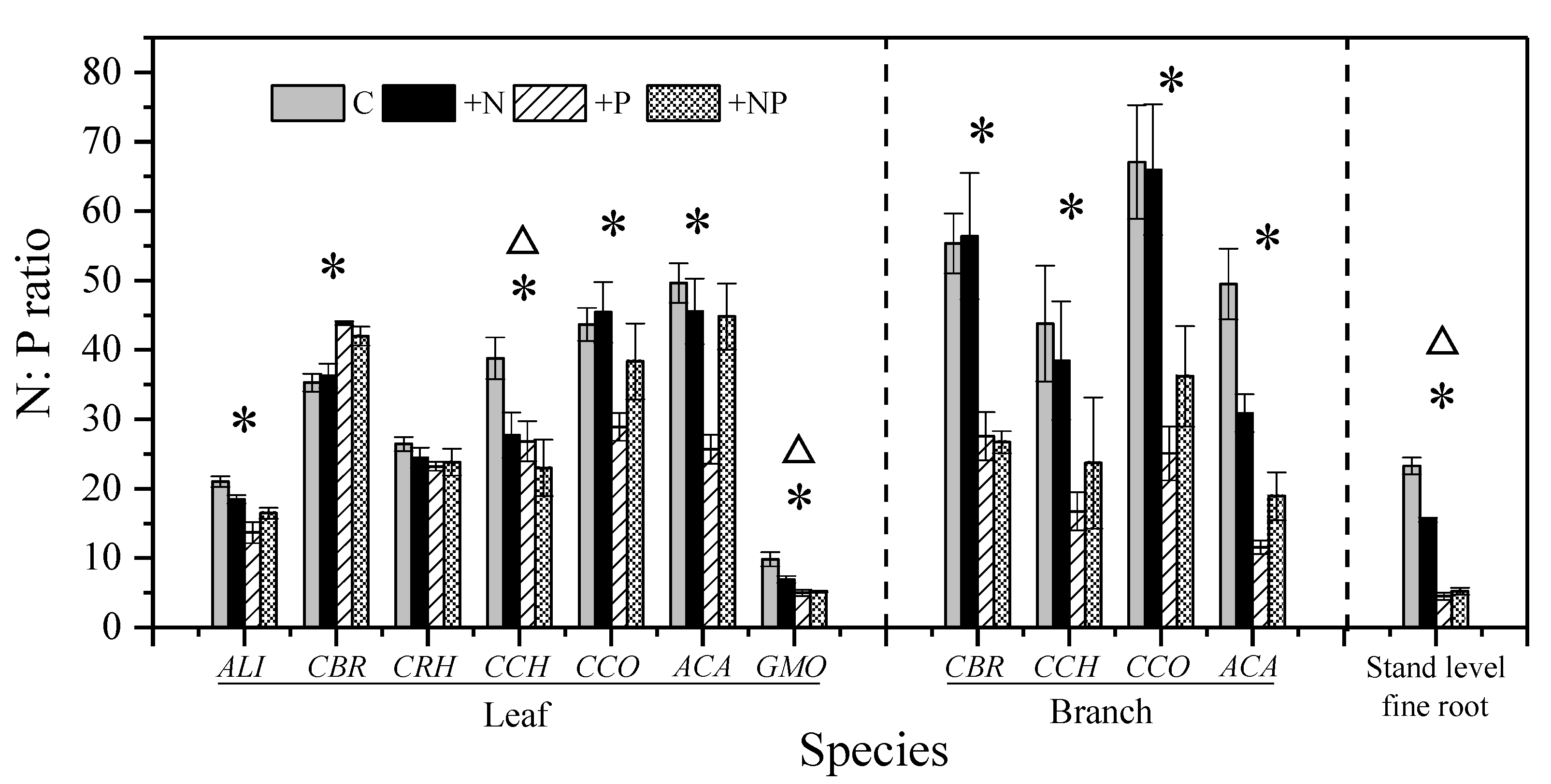
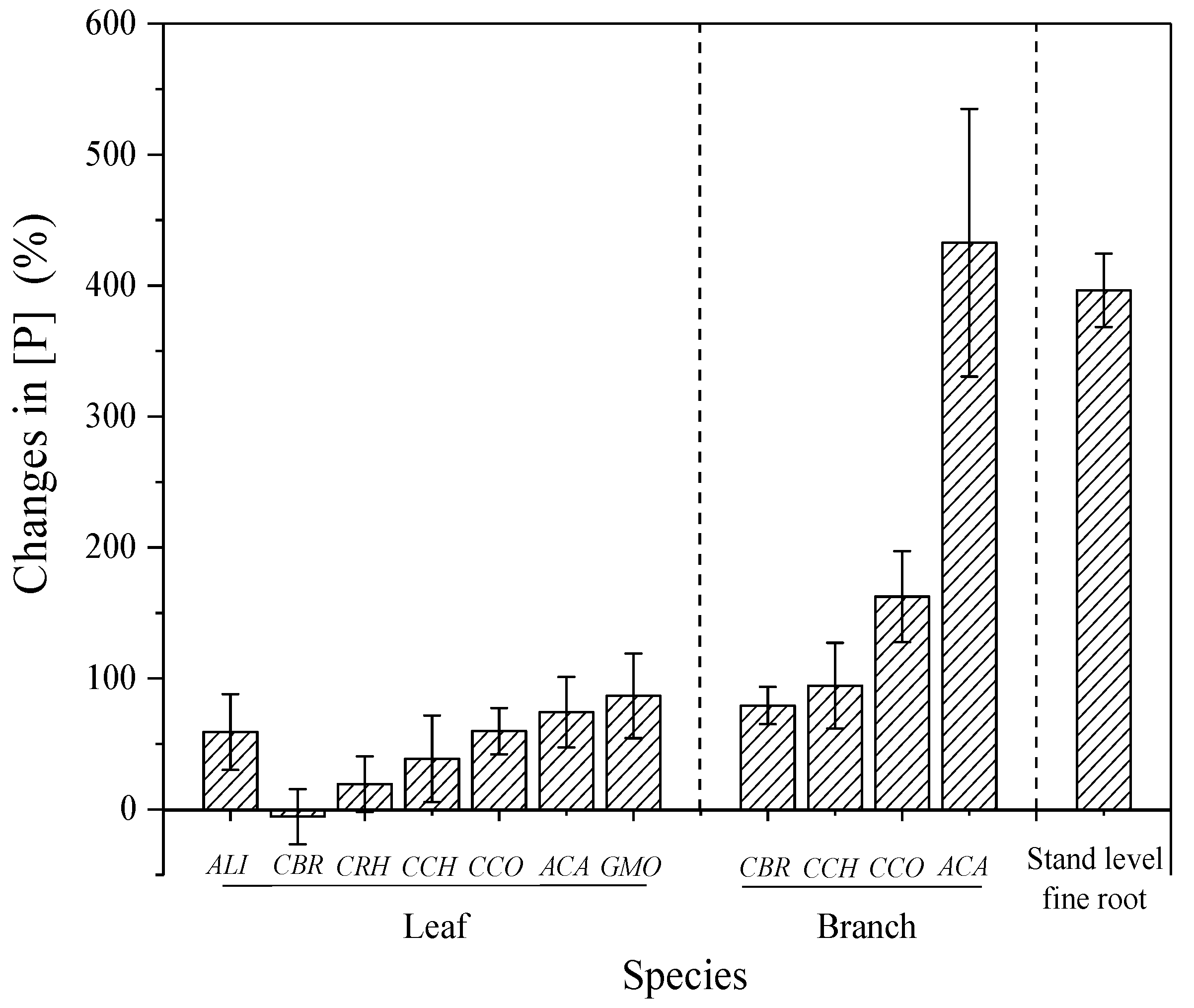
3.2. Effect of N and P Addition on the Nutrient Status of Understory Vegetation
4. Discussion
4.1. Effects of N Addition
4.2. Effects of P Addition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raven, P.H.; Gereau, R.E.; Phillipson, P.B.; Chatelain, C.; Jenkins, C.N.; Ulloa, C.U. The distribution of biodiversity richness in the tropics. Sci. Adv. 2020, 6, eabc6228. [Google Scholar] [CrossRef]
- Vitousek, P.; Porder, S.; Houlton, B.; Chadwick, O. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen- phosphorous interactions. Ecol. Appl. 2010, 20, 10. [Google Scholar] [CrossRef] [Green Version]
- Galloway, J.N.; Dentener, F.J.; Capone, D.G.; Boyer, E.W.; Howarth, R.W.; Seitzinger, S.P.; Asner, G.P.; Cleveland, C.C.; Green, P.A.; Holland, E.A.; et al. Nitrogen Cycles: Past, Present, and Future. Biogeochemistry 2004, 70, 153–226. [Google Scholar] [CrossRef]
- Yuan, Z.; Jiang, S.; Sheng, H.; Liu, X.; Hua, H.; Liu, X.; Zhang, Y. Human perturbation of the global phosphorus cycle: Changes and consequences. Environ. Sci. Technol. 2018, 52, 2438–2450. [Google Scholar] [CrossRef]
- Swaine, M.D. Rainfall and Soil Fertility as Factors Limiting Forest Species Distributions in Ghana. J. Ecol. 1996, 84, 419. [Google Scholar] [CrossRef]
- Zalamea, P.; Turner, B.L.; Winter, K.; Jones, F.A.; Sarmiento, C.; Dalling, J.W. Seedling growth responses to phosphorus reflect adult distribution patterns of tropical trees. New Phytol. 2016, 212, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Aber, J.; McDowell, W.; Nadelhoffer, K.; Magill, A.; Berntson, G.; Kamakea, M.; McNulty, S.; Currie, W.; Rustad, L.; Fernandez, I. Nitrogen Saturation in Temperate Forest Ecosystems. BioScience 1998, 48, 921–934. [Google Scholar] [CrossRef]
- Bleeker, A.; Hicks, W.; Dentener, F.; Galloway, J.; Erisman, J. N deposition as a threat to the World’s protected areas under the Convention on Biological Diversity. Environ. Pollut. 2011, 159, 2280–2288. [Google Scholar] [CrossRef]
- Mayor, J.R.; Wright, S.J.; Turner, B.L. Species-specific responses of foliar nutrients to long-term nitrogen and phosphorus additions in a lowland tropical forest. J. Ecol. 2013, 102, 36–44. [Google Scholar] [CrossRef]
- Mao, Q.; Lu, X.; Mo, H.; Gundersen, P.; Mo, J. Effects of simulated N deposition on foliar nutrient status, N metabolism and photosynthetic capacity of three dominant understory plant species in a mature tropical forest. Sci. Total Environ. 2018, 610-611, 555–562. [Google Scholar] [CrossRef]
- Fang, Y.; Gundersen, P.; Mo, J.M.; Zhu, W.X. Input and output of dissolved organic and inorganic nitrogen in subtropical forests of South China under high air pollution. Biogeosciences 2008, 5, 339–352. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Gurmesa, G.A.; Zhang, W.; Zhu, X.; Zheng, M.; Mao, Q.; Zhang, T.; Mo, J. Nitrogen saturation in humid tropical forests after 6 years of nitrogen and phosphorus addition: Hypothesis testing. Funct. Ecol. 2015, 30, 305–313. [Google Scholar] [CrossRef]
- Deng, Q.; Hui, D.F.; Dennis, S.; Reddy, K.C. Responses of terrestrial ecosystem phosphorus cycling to nitrogen addition: A meta-analysis. Glob. Ecol. Biogeogr. 2017, 26, 713–728. [Google Scholar] [CrossRef]
- Wang, C.; Mori, T.; Mao, Q.; Zhou, K.; Wang, Z.; Zhang, Y.; Mo, H.; Lu, X.; Mo, J. Responses of soil microbial community to continuous experimental nitrogen additions for 13 years in a nitrogen-rich tropical forest. Soil Biol. Biochem. 2018, 121, 103–112. [Google Scholar] [CrossRef]
- Deng, M.; Liu, L.; Sun, Z.; Piao, S.; Ma, Y.; Chen, Y.; Wang, J.; Qiao, C.; Wang, X.; Li, P. Increased phosphate uptake but not resorption alleviates phosphorus deficiency induced by nitrogen deposition in temperate Larix principisrupprechtii plantations. New Phytol. 2016, 212, 1019–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Van Groenigen, K.J.; Hungate, B.A.; Terrer, C.; Van Groenigen, J.; Maestre, F.T.; Ying, S.C.; Luo, Y.; Jørgensen, U.; Sinsabaugh, R.L.; et al. Long-term nitrogen loading alleviates phosphorus limitation in terrestrial ecosystems. Glob. Chang. Biol. 2020, 26, 5077–5086. [Google Scholar] [CrossRef]
- Pant, H.K.; Warman, P.R. Enzymatic hydrolysis of soil organic phosphorus by immobilized phosphatases. Biol. Fertil. Soils 2000, 30, 306–311. [Google Scholar] [CrossRef]
- Zhou, K.; Lu, X.; Mori, T.; Mao, Q.; Wang, C.; Zheng, M.; Mo, H.; Hou, E.; Mo, J. Effects of long-term nitrogen deposition on phosphorus leaching dynamics in a mature tropical forest. Biogeochemistry 2018, 138, 215–224. [Google Scholar] [CrossRef]
- You, C.; Wu, F.; Yang, W.; Xu, Z.; Tan, B.; Zhang, L.; Yue, K.; Ni, X.; Li, H.; Chang, C.; et al. Does foliar nutrient resorption regulate the coupled relationship between nitrogen and phosphorus in plant leaves in response to nitrogen deposition? Sci. Total Environ. 2018, 645, 733–742. [Google Scholar] [CrossRef]
- Lawrence, D. The response of tropical tree seedlings to nutrient supply: Meta-analysis for understanding a changing tropical landscape. J. Trop. Ecol. 2003, 19, 239–250. [Google Scholar] [CrossRef]
- Ostertag, R. Foliar nitrogen and phosphorus accumulation responses after fertilization: An example from nutrient-limited Hawaiian forests. Plant Soil 2010, 334, 85–98. [Google Scholar] [CrossRef]
- Cárate-Tandalla, D.; Camenzind, T.; Leuschner, C.; Homeier, J. Contrasting species responses to continued nitrogen and phosphorus addition in tropical montane forest tree seedlings. Biotropica 2018, 50, 234–245. [Google Scholar] [CrossRef]
- Brearley, F.Q.; Scholes, J.; Press, M.C.; Palfner, G. How does light and phosphorus fertilisation affect the growth and ectomycorrhizal community of two contrasting dipterocarp species? Plant Ecol. 2007, 192, 237–249. [Google Scholar] [CrossRef]
- Burslem, D.F.R.P.; Grubb, P.J.; Turner, I.M. Responses to Nutrient Addition among Shade-Tolerant Tree Seedlings of Lowland Tropical Rain Forest in Singapore. J. Ecol. 1995, 83, 113. [Google Scholar] [CrossRef]
- Schreeg, L.A.; Santiago, L.S.; Wright, S.J.; Turner, B.L. Stem, root, and older leaf N: P ratios are more responsive indicators of soil nutrient availability than new foliage. Ecology 2014, 95, 2062–2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Gundersen, P.; Zhang, T.; Mo, J. Effects of phosphorus addition on soil microbial biomass and community composition in three forest types in tropical China. Soil Biol. Biochem. 2012, 44, 31–38. [Google Scholar] [CrossRef]
- Zhang, T.; Zhu, W.; Mo, J.; Liu, L.; Dong, S. Increased phosphorus availability mitigates the inhibition of nitrogen deposition on CH4 uptake in an old-growth tropical forest, southern China. Biogeosciences 2011, 8, 2805–2813. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Wang, Y.; Zhang, X.; Dörsch, P.; Mulder, J. Phosphorus addition mitigates N2O and CH4 emissions in N-saturated subtropical forest, SW China. Biogeosciences 2017, 14, 3097–3109. [Google Scholar] [CrossRef] [Green Version]
- Neufeld, H.S.; Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology. Ecology 1999, 80, 1785–1787. [Google Scholar] [CrossRef]
- Gilliam, F.S. The Ecological Significance of the Herbaceous Layer in Temperate Forest Ecosystems. Bioscience 2007, 57, 845–858. [Google Scholar] [CrossRef]
- Lu, X.; Mo, J.; Gilliam, F.S.; Zhou, G.; Fang, Y. Effects of experimental nitrogen additions on plant diversity in an old-growth tropical forest. Glob. Chang. Biol. 2010, 16, 2688–2700. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.F.; Fan, Z.G. The Climate of Dinghushan, Tropical and Subtropical Forest Ecosystem; Science Press: Beijing, China, 1983. [Google Scholar]
- Mo, J.; Brown, S.; Peng, S.; Kong, G. Nitrogen availability in disturbed, rehabilitated and mature forests of tropical China. For. Ecol. Manag. 2003, 175, 573–583. [Google Scholar] [CrossRef]
- Lu, X.K.; Vitousek, P.M.; Mao, Q.G.; Gilliam, F.S.; Luo, Y.Q.; Zhou, G.Y.; Zou, X.; Bai, E.; Scanlon, T.M.; Hou, E.; et al. Plant acclimation to long-term high nitrogen deposition in an N-rich tropical forest. Proc. Natl. Acad. Sci. USA 2018, 115, 5187–5192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, Q.; Lu, X.; Zhou, K.; Chen, H.; Zhu, X.; Mori, T.; Mo, J. Effects of long-term nitrogen and phosphorus additions on soil acidification in an N-rich tropical forest. Geoderma 2017, 285, 57–63. [Google Scholar] [CrossRef]
- Shen, C.; Liu, D.; Peng, S.; Sun, Y.; Jiang, M.; Yi, W.; Xing, C.; Gao, Q.; Li, Z.; Zhou, G. 14C measurement of forest soils in Dinghushan Biosphere Reserve. Chin. Sci. Bull. 1999, 44, 251–256. [Google Scholar] [CrossRef]
- Mirabello, M.J.; Yavitt, J.B.; García, M.; Harms, K.E.; Turner, B.L.; Wright, S.J. Soil phosphorus responses to chronic nutrient fertilisation and seasonal drought in a humid lowland forest, Panama. Soil Res. 2013, 51, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.M.; Ingram, J.S.I. Tropical Soil Biology and Fertility. Soil Sci. 1994, 157, 265. [Google Scholar] [CrossRef]
- Hedin, L.O.; Brookshire, E.N.J.; Menge, D.N.L.; Barron, A.R. The Nitrogen Paradox in Tropical Forest Ecosystems. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 613–635. [Google Scholar] [CrossRef] [Green Version]
- Gurmesa, G.A.; Lu, X.; Gundersen, P.; Mao, Q.; Zhou, K.; Fang, Y.; Mo, J. High retention of 15N-labeled nitrogen deposition in a nitrogen saturated old-growth tropical forest. Glob. Chang. Biol. 2016, 22, 3608–3620. [Google Scholar] [CrossRef]
- Wright, S.J.; Turner, B.L.; Sheldrake, M.; Garcia, M.N.; Yavitt, J.B.; Harms, K.E.; Kaspari, M.; Tanner, E.V.J.; Bujan, J.; Griffin, E.A.; et al. Plant responses to fertilization experiments in lowland, species-rich, tropical forests. Ecology 2018, 99, 1129–1138. [Google Scholar] [CrossRef]
- Throop, H.L.; Lerdau, M.T. Effects of nitrogen deposition on insect herbivory: Implications for community and ecosystem processes. Ecosystems 2004, 7, 109–133. [Google Scholar] [CrossRef]
- Lu, X.; Mao, Q.; Gilliam, F.S.; Luo, Y.; Mo, J. Nitrogen deposition contributes to soil acidification in tropical ecosystems. Glob. Chang. Biol. 2014, 20, 3790–3801. [Google Scholar] [CrossRef]
- Treseder, K.K.; Vitousek, P.M. Effects of soil nutrient availability on investment in acquisition of N and P in Hawaiian rain forests. Ecology 2001, 82, 946–954. [Google Scholar] [CrossRef]
- Zheng, M.; Huang, J.; Chen, H.; Wang, H.; Mo, J. Responses of soil acid phosphatase and beta-glucosidase to nitrogen and phosphorus addition in two subtropical forests in southern China. Eur. J. Soil Biol. 2015, 68, 77–84. [Google Scholar] [CrossRef]
- Mo, Q.; Li, Z.; Sayer, E.J.; Lambers, H.; Li, Y.; Zou, B.; Tang, J.; Heskel, M.; Ding, Y.; Wang, F. Foliar phosphorus fractions reveal how tropical plants maintain photosynthetic rates despite low soil phosphorus availability. Funct. Ecol. 2019, 33, 503–513. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.J. Plant responses to nutrient addition experiments conducted in tropical forests. Ecol. Monogr. 2019, 89. [Google Scholar] [CrossRef]
- Sardans, J.; Vallicrosa, H.; Zuccarini, P.; Farré-Armengol, G.; Fernández-Martínez, M.; Peguero, G.; Gargallo-Garriga, A.; Ciais, P.; Janssens, I.A.; Obersteiner, M.; et al. Empirical support for the biogeochemical niche hypothesis in forest trees. Nat. Ecol. Evol. 2021, 5, 184–194. [Google Scholar] [CrossRef]
- McGroddy, M.E.; Daufresne, T.; Hedin, L.O. Scaling of C:N:P stoichiometry in forests worldwide: Implications of terrestrial redfield-type ratios. Ecology 2004, 85, 2390–2401. [Google Scholar] [CrossRef]
- Townsend, A.R.; Cleveland, C.C.; Asner, G.; Bustamante, M.M.C. Controls over foliar N: P ratios in tropical rain forests. Ecology 2007, 88, 107–118. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Abbreviation | Family | Growth Form |
|---|---|---|---|
| Ardisia lindleyana D. Dietrich | ALI | Myrsinaceae | shrub |
| Carallia brachiata Merr. | CBR | Rhizophoraceae | subcanopy tree seedling |
| Calamus rhabdocladus Burret | CRH | Arecaceae | palm liana |
| Cryptocarya chinensis Hemsl | CCH | Lauraceae | canopy tree seedling |
| Cryptocarya concinna Hance | CCO | Lauraceae | canopy tree seedling |
| Aidia canthioides Masam | ACA | Rubiaceae | subcanopy tree seedling |
| Gnetum montanum Markgr. | GMO | Gnetaceae | woody liana |
| Control | +N | +P | +NP | |
|---|---|---|---|---|
| SOM (g kg−1) | 31.15 (0.74) | 29.28 (0.39) | 30.50 (2.75) | 30.28 (1.66) |
| TN (g kg−1) | 1.88 (0.09) | 1.82 (0.06) | 1.79 (0.10) | 1.76 (0.09) |
| TP (g kg−1) | 0.20 (0.02) b | 0.22 (0.02) b | 0.62 (0.04) a | 0.47 (0.06) a |
| Extractable N (mg kg−1) | 12.70 (0.91) ab | 14.93 (0.88) ab | 11.51 (0.82) b | 16.40 (1.57) a |
| Extractable P (mg kg−1) | 3.26 (0.30) b | 5.28 (1.25) b | 111.76 (20.64) a | 73.09 (10.83) a |
| pH(H2O) | 3.88(0.04) ab | 3.70 (0.02) c | 3.95 (0.04) a | 3.77 (0.04) bc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, Q.; Chen, H.; Wang, C.; Pang, Z.; Mo, J.; Lu, X. Effect of Long-Term Nitrogen and Phosphorus Additions on Understory Plant Nutrients in a Primary Tropical Forest. Forests 2021, 12, 803. https://doi.org/10.3390/f12060803
Mao Q, Chen H, Wang C, Pang Z, Mo J, Lu X. Effect of Long-Term Nitrogen and Phosphorus Additions on Understory Plant Nutrients in a Primary Tropical Forest. Forests. 2021; 12(6):803. https://doi.org/10.3390/f12060803
Chicago/Turabian StyleMao, Qinggong, Hao Chen, Cong Wang, Zongqing Pang, Jiangming Mo, and Xiankai Lu. 2021. "Effect of Long-Term Nitrogen and Phosphorus Additions on Understory Plant Nutrients in a Primary Tropical Forest" Forests 12, no. 6: 803. https://doi.org/10.3390/f12060803
APA StyleMao, Q., Chen, H., Wang, C., Pang, Z., Mo, J., & Lu, X. (2021). Effect of Long-Term Nitrogen and Phosphorus Additions on Understory Plant Nutrients in a Primary Tropical Forest. Forests, 12(6), 803. https://doi.org/10.3390/f12060803