
Geographical Structuring of Quercus robur (L.) Chloroplast DNA Haplotypes in Lithuania: Recolonization, Adaptation, or Overexploitation Effects?


Abstract
:1. Introduction
2. Material and Methods
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Eaton, E.; Caudullo, G.; Oliveira, S.; de Rigo, D. Quercus robur and Quercus petraea in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Karazija, S. (Ed.) Historical Review of Oaks Forests in Lithuania. In Oak Stands in Lithuania: Protection and Reforestation Problems; Lithuanian Forest Research Institute: Kaunas, Lithuania, 1997; pp. 13–17. (In Lithuanian) [Google Scholar]
- Ducousso, A.; Bordacs, S. EUFORGEN Technical Guidelines for Genetic Conservation and Use for Pedunculate and Sessile Oaks (Quercus robur and Q. petraea); International Plant Genetic Resources Institute: Rome, Italy, 2004. [Google Scholar]
- Kremer, A.; Petit, R.J. Gene diversity in natural populations of oak species. Ann. Sci. For. 1993, 50, 186–202. [Google Scholar] [CrossRef] [Green Version]
- Tomiczek, C. Oak decline in Austria and Europe. J. Arboric. 1993, 19, 71–73. [Google Scholar]
- Gustiene, A.; Vasiliauskas, A. Oak decline in Lithuania: Preliminary estimating of extend of the process and role of fungi. Miskininkyste 2006, 2, 5–11, (In Lithuanian with English summary, Figure, Table headings). [Google Scholar]
- Finch, J.P.; Brown, N.; Beckmann, M.; Denman, S.; Draper, J. Index measures for oak decline severity using phenotypic descriptors. For. Ecol. Manag. 2021, 485, 118948. [Google Scholar] [CrossRef]
- Reed, K.; Brown, N.; Vanguelova, E.; Finch, J.; Denman, S. Current Understanding of Acute Oak Decline: A summary of recent research with implications for woodland management. Q. J. For. 2021, 115, 38–45. [Google Scholar]
- Aldrich, P.R.; Glaubitz, J.C.; Parker, G.R.; Rhodes, O.E.; Michler, C.H. Genetic Structure Inside a Declining Red Oak Community in Old-Growth Forest. J. Hered. 2005, 96, 627–634. [Google Scholar] [CrossRef] [Green Version]
- Csaikl, U.M.; Glaz, I.; Baliuckas, V.; Petit, R.J.; Jensenen, J.S. Chloroplast DNA variation of white oak in the Baltic countries of Poland. Chloroplast DNA variation of white oak in the Baltic countries of Poland. For. Ecol. Manag. 2002, 156, 211–222. [Google Scholar] [CrossRef]
- Dumolin, S.; Demesure, B.; Petit, R.J. Inheritance of chloroplast and mitochondrial genomes in pedunculate oak investigated with an efficient PCR method. Theor. Appl. Genet. 1995, 91, 1253–1256. [Google Scholar] [CrossRef]
- Clegg, M.T.; Brandon, S.G.; Learn, G.H.; Morton, B. Rates and patterns of chloroplast DNA evolution. Proc. Natl. Acad. Sci. USA 1994, 91, 6795–6801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petit, R.J.; Kremer, A.; Wagner, D.B. Geographic structure of chloroplast DNA polymorphisms in European oaks. Theor. Appl. Genet. 1993, 87, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Dumolin-Lapegue, S.; Demesure, B.; Le Corre, V.; Fineschi, S.; Petit, R.J. Phylogeographic structure of white oaks throughout the European continent. Genetics 1997, 146, 1475–1487. [Google Scholar] [CrossRef]
- Huntley, B.; Birks, H.J.B. An Atlas of Past and Present Pollen Maps for Europe 0–13,000 Years Ago; Cambridge University Press: Cambridge, UK, 1983. [Google Scholar]
- Tollefsrud, M.M.; Sønstebø, J.H.; Brochmann, C.; Johnsen, Ø.; Skrøppa, T.; Vendramin, G.G. Combined analysis of nuclear and mitochondrial markers provide new insight into the genetic structure of North European Picea abies. Heredity 2009, 102, 549–562. [Google Scholar] [CrossRef] [Green Version]
- Pigott, C.D. Lime-Trees and Basswoods: A Biological Monograph of the Genus Tilia; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Danusevičius, D.; Kembrytė, R.; Buchovska, J.; Baliuckas, V.; Kavaliauskas, D. Genetic signature of the natural genepool of Tilia cordata Mill. in Lithuania: Compound evolutionary and anthropogenic effects. Ecol. Evol. 2021, (in press). [Google Scholar] [CrossRef]
- Petit, R.J.; Csaikl, U.M.; Bordács, S.; Burg, K.; Coart, E.; Cottrell, J.; van Dam, B.; Deans, J.D.; Dumolin-Lapègue, S.; Fineschi, S.; et al. Chloroplast DNA variation in European white oaks phylogeography and patterns of diversity based on data from over 2600 populations. For. Ecol. Manag. 2002, 156, 5–26. [Google Scholar] [CrossRef]
- Gailing, O.; Wachter, H.; Heyder, J.; Schmitt, H.P.; Finkeldey, R. Chloroplast DNA analysis in oak stands (Quercus robur L.) in North Rhine-Westphalia with presumably Slavonian origin: Is there an association between geographic origin and bud phenology? J. Appl. Bot. Food Qual. 2007, 81, 165–171. [Google Scholar]
- Semerikova, S.A.; Isakov, I.Y.; Semerikov, V.L. Chloroplast DNA Variation and Phylogeography of Pedunculate Oak Quercus robur L. in the Eastern Part of the Range. Russ. J. Genet. 2021, 57, 47–60. [Google Scholar] [CrossRef]
- Pliura, A.; Rungis, D.; Baliuckas, V. Population structure of pedunculate oak in Lithuania based on analysis of chloroplast DNA haplotypes and adaptive traits. Balt. For. 2009, 15, 212. [Google Scholar]
- Neophytou, C.; Michiels, H.G. Upper Rhine Valley: A migration crossroads of middle European oaks. For. Ecol. Manag. 2013, 304, 89–98. [Google Scholar]
- Huntley, B. European post-glacial forests: Compositional changes in response to climatic change. J. Veg. Sci. 1990, 1, 507–518. [Google Scholar] [CrossRef]
- Kvist, L. Phylogeny and Phylogeography of European Parids. Ph.D. Thesis, University of Oulu, Oulu, Finland, 2000. Available online: http://urn.fi/urn:isbn:9514255364 (accessed on 13 March 2021).
- Giesecke, T.; Brewer, S.; Finsinger, W.; Leydet, M.; Bradshaw, R.H.W. Patterns and dynamics of European vegetation change over the last 15,000 years. J. Biogeogr. 2017, 44, 1441–1456. [Google Scholar] [CrossRef] [Green Version]
- Buchovska, J.; Danusevicius, D.; Stanys, V.; Šikšnianienė, J.B.; Kavaliauskas, D. The location of the northern glacial refugium of Scots pine based on mitochondrial DNA markers. Balt. For. 2013, 19, 2–12. [Google Scholar]
- Birks, H.J.B.; Tinner, W. Past forests of Europe. In European Atlas of Forest Tree Species; San Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016; p. 1045. [Google Scholar]
- Balakauskas, L. Forest Vegetation During Late Glaciation Holocene in Lithuania Mased on LRA Modeling. Ph.D. Thesis, Vilnius University, Vilnius, Lithuania, 2012. (In Lithuanian with English summary). [Google Scholar]
- Žulkus, V.; Girininkas, A. The eastern shores of the Baltic Sea in the Early Holocene according to natural and cultural relict data. Geo: Geogr. Environ. 2020, 7, e00087. [Google Scholar] [CrossRef]
- Kremer, A.J.; Kleinschmit, J.; Cottrell, E.P.; Cundall, J.D.; Deans, A.; Ducousso, A.O.; Konig, A.J.; Lowe, R.C.; Munro, R.J.; Petit, B.R.S. Is there a correlation between chloroplastic and nuclear divergence, or what are the roles of history and selection on genetic diversity in European oaks? For. Ecol. Manag. 2002, 156, 75–87. [Google Scholar] [CrossRef]
- Petit, R.J.; Bodenes, C.; Ducousso, A.; Roussel, G.; Kremer, A. Hybridization as a mechanism of invasion in oaks. New Phytol. 2004, 161, 151–164. [Google Scholar] [CrossRef] [Green Version]
- Bussotti, F.; Grossoni, P. European and Mediterranean oaks (Quercus, L.; Fagaceae): SEM Characterization of the Micromorphology of the Abaxial Leaf Surface. Bot. J. Linnaean Soc. 1997, 124, 183–199. [Google Scholar] [CrossRef]
- Dupouey, J.L.; Badeau, V. Morphological Variability of Oaks (Quercus robur L., Quercus petraea (Matt) Liebl, Quercus pubescens Willd.) in Northeastern France: Preliminary Results. Ann. For. Sci. 1993, 50, 35s–40s. [Google Scholar] [CrossRef]
- del Río, S.; Álvarez, R.; Candelas, A.; González-Sierra, S.; Herrero, L.; Penas, A. Preliminary Study on Taxonomic Review Using Histological Sections of Some Iberian Species from the Genus Quercus L. (Fagaceae). Am. J. Plant Sci. 2014, 5, 2773–2784. [Google Scholar]
- Enescu, C.M. A Dichotomous Determination Key for Autochthonous Oak Species from Romania. J. Hortic. For. Biotechnol. 2017, 21, 58–62. [Google Scholar]
- Rushton, B.S. Natural Hybridization within the Genus Quercus L. Ann. For. Sci. 1993, 50, 73–90. [Google Scholar] [CrossRef] [Green Version]
- Schicchi, R.; Mazzola, P.; Raimondo, F.M. Eco-Morphologic and Taxonomic Studies of Quercus Hybrids (Fagaceae) in Sicily. Bocconea 2001, 13, 485–490. [Google Scholar]
- Tamošaitis, S.; Jurkšienė, G.; Petrokas, R.; Buchovska, J.; Kavaliauskienė, I.; Danusevičius, D.; Baliuckas, V. Dissecting Taxonomic Variants within Ulmus spp. Complex in Natural Forests with the Aid of Microsatellite and Morphometric Markers. Forests 2021, 12, 653. [Google Scholar] [CrossRef]
- Danusevicius, D.; Marozas, V.; Brazaitis, G.; Petrokas, R.; Christensen, K.I. Spontaneous hybridization between Pinus mugo (Turra) and Pinus sylvestris (L.) at the Lithuanian sea-side: A morphological survey. Sci. World J. 2012, 2012, 172407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danusevičius, D.; Buchovska, J.; Stanys, V.; Šikšnianiene, J.B.; Marozas, V.; Bendokas, V. DNA marker based identification of spontaneous hybrids between Pinus mugo and Pinus sylvestris at the Lithuanian sea-side. Nord. J. Bot. 2013, 31, 001–009. [Google Scholar] [CrossRef]
- Lithuanian Forest Statistics Yearbook for 2019; State Forest Service: Kaunas, Lithuania, 2020.
- Dumolin-Lapègue, S.; Pemonge, M.H.; Gielly, L.; Taberlet, P.; Petit, R.J. Amplification of oak DNA from ancient and modern wood. Mol. Ecol. 1999, 8, 2137–2140. [Google Scholar] [CrossRef] [PubMed]
- Deguilloux, M.F.; Dumolin-Lapègue, S.; Gielly, L.; Grivet, D.; Petit, R.J. A set of primers for the amplification of chloroplast microsatellites in Quercus. Mol. Ecol. Notes 2003, 3, 24–27. [Google Scholar] [CrossRef]
- User Bulletin GeneMapper® Software; Version 4.0; Applied Biosystems: Foster City, CA, USA, 2006.
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Manni, F.; Guerard, E.; Heyer, E. Geographic Patterns of (Genetic, Morphologic, Linguistic) Variation: How Barriers Can Be Detected by Using Monmonier’s Algorithm. Hum. Biol. 2004, 76, 173–190. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Tajima, F.; Tateno, Y. Accuracy of estimated phylogenetic trees from molecular data. II. Gene frequency data. J. Mol. Evol. 1983, 19, 153–170. [Google Scholar] [CrossRef]
- Kembrytė, R.; Danusevičius, D.; Buchovska, J.; Baliuckas, V.; Kavaliauskas, D.; Fussi, B.; Kempf, M. DNA-based tracking of historical introductions of forest trees: The case of European beech (Fagus sylvatica L.) in Lithuania. Eur. J. For. Res. 2021, 140, 435–449. [Google Scholar] [CrossRef]
- Deltuvas, R. Forestry and hunting in Little Lithuania. Monography; Klaipeda University Press: Klaipėda, Lithuania, 2019. [Google Scholar]
- König, A.O.; Ziegenhagen, B.; van Dam, B.C.; Csaikl, U.M.; Coartd, E.; Degena, B.; Burg, K.; de Vries, S.M.G.; Petita, R.J. Chloroplast DNA variation of oaks in western Central Europe and genetic consequences of human influences. For. Ecol. Manag. 2002, 156, 147166. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
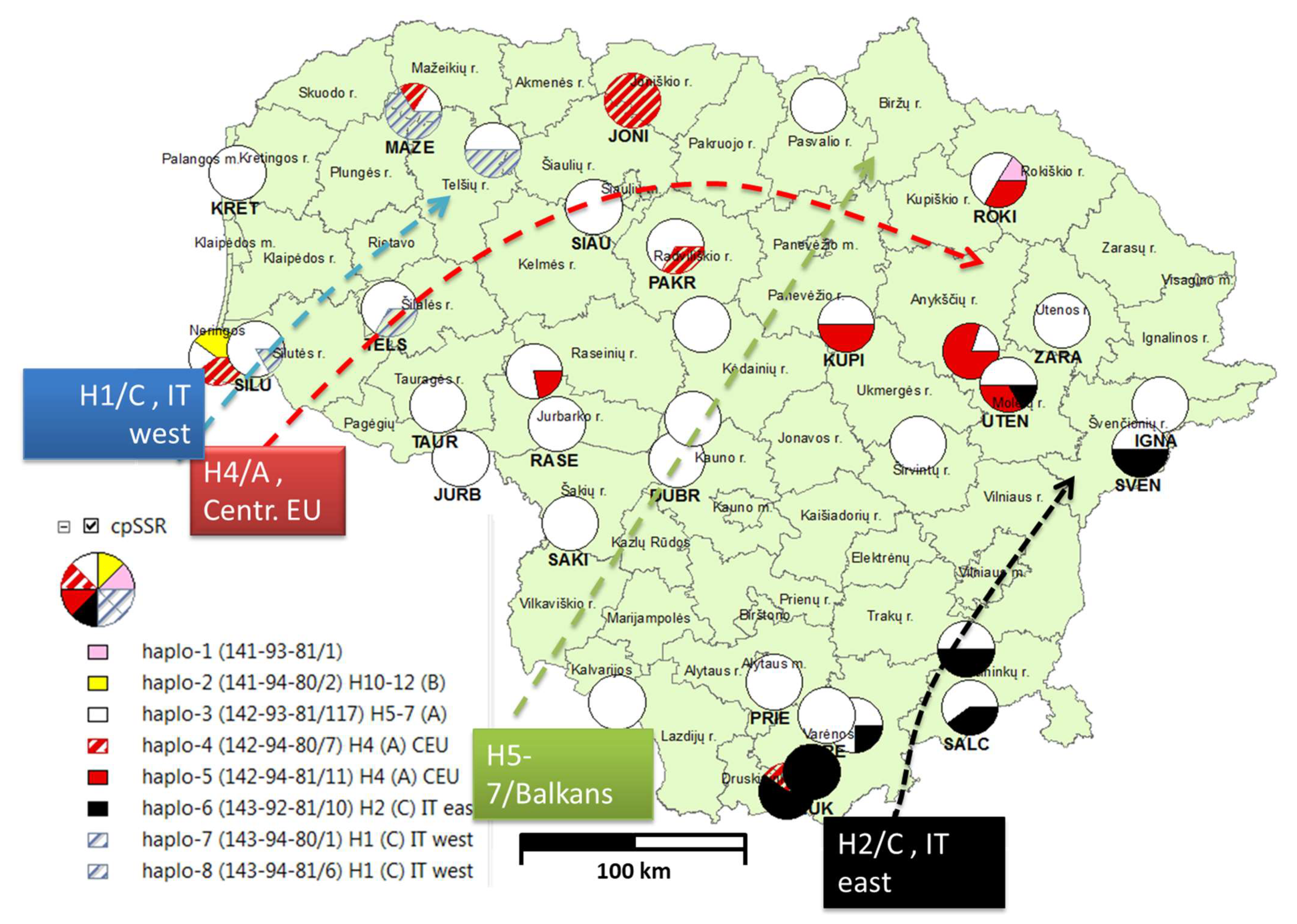
| Pop | Lat | Long | N | Haplo-1 | Haplo-2 | Haplo-3 | Haplo-4 | Haplo-5 | Haplo-6 | Haplo-7 | Haplo-8 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| NF | B Iberia H10-12 | A Balcan H5-7 | A CEU H4 | A CEU H4 | C ITeast H2 | C ITwest H1 | C ITwest H1 | ||||
| 141-93-81 | 141-94-80 | 142-93-81 | 142-94-80 | 142-94-81 | 143-92-81 | 143-94-80 | 143-94-81 | ||||
| Total | 157 | 1 | 4 | 117 | 7 | 11 | 10 | 1 | 6 | ||
| ANYK | 55.39 | 25.18 | 5 | 0 | 0 | 1 | 0 | 4 | 0 | 0 | 0 |
| BIRZ | 56.17 | 24.35 | 5 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 |
| DRUS | 54.00 | 24.17 | 5 | 0 | 0 | 0 | 2 | 0 | 3 | 0 | 0 |
| DUBR | 54.54 | 23.48 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| DZUK | 54.06 | 24.30 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| IGNA | 55.21 | 26.20 | 2 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 |
| JONI | 56.19 | 23.32 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| JURB | 55.04 | 22.41 | 5 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 |
| KEDA | 55.18 | 23.66 | 3 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 |
| KRET | 55.93 | 21.14 | 10 | 0 | 0 | 10 | 0 | 0 | 0 | 0 | 0 |
| KUPI | 55.48 | 24.50 | 2 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 |
| KURS | 56.03 | 22.55 | 2 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| MARI | 54.43 | 22.58 | 5 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 |
| MAZE | 56.14 | 22.10 | 6 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 4 |
| NERI | 55.34 | 21.07 | 7 | 0 | 4 | 1 | 2 | 0 | 0 | 0 | 0 |
| PAKR | 55.73 | 23.56 | 3 | 0 | 0 | 2 | 1 | 0 | 0 | 0 | 0 |
| PRIE | 54.35 | 24.10 | 6 | 0 | 0 | 6 | 0 | 0 | 0 | 0 | 0 |
| RADV | 55.38 | 23.52 | 7 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 |
| RASE | 55.16 | 22.93 | 6 | 0 | 0 | 6 | 0 | 0 | 0 | 0 | 0 |
| ROKI | 55.93 | 25.35 | 6 | 1 | 0 | 3 | 0 | 2 | 0 | 0 | 0 |
| SAKI | 54.84 | 23.01 | 5 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 |
| SALC | 54.26 | 25.14 | 5 | 0 | 0 | 3 | 0 | 0 | 2 | 0 | 0 |
| SIAU | 55.85 | 23.11 | 3 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 |
| SILU | 55.37 | 21.28 | 6 | 0 | 0 | 5 | 0 | 0 | 0 | 1 | 0 |
| SVEN | 55.07 | 26.09 | 2 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 |
| TAUR | 55.21 | 22.28 | 2 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 |
| TELS | 55.51 | 22.00 | 3 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 1 |
| TYTU | 55.33 | 22.80 | 9 | 0 | 0 | 7 | 0 | 2 | 0 | 0 | 0 |
| UKME | 55.10 | 24.88 | 8 | 0 | 0 | 8 | 0 | 0 | 0 | 0 | 0 |
| UTEN | 55.28 | 25.38 | 6 | 0 | 0 | 3 | 0 | 2 | 1 | 0 | 0 |
| VALK | 54.21 | 24.53 | 4 | 0 | 0 | 3 | 0 | 0 | 1 | 0 | 0 |
| VARE | 54.24 | 24.38 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| VEIS | 54.05 | 23.39 | 6 | 0 | 0 | 6 | 0 | 0 | 0 | 0 | 0 |
| VILN | 54.45 | 25.13 | 2 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 |
| ZARA | 55.48 | 25.68 | 7 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 |
| Locus | Primer Sequence (5′–3′) | Repeat | Size, bp | Na | Ta (°C) | GeneBank Accession id, Genomic Location |
|---|---|---|---|---|---|---|
| udt1 | NED-ATCTTACACTAAGCTCGGAA TTCAATAACTTGTTGATCCC | (A)11 | 80–81 | 2 | 48 | AJ489829, intergenic trnE-trnT |
| udt4 | FAM-GATAATATAAAGAGTCAAAT CCGAAAGGTCCTATACCTCG | (A)9 | 141–143 | 3 | 44 TD | AJ489831, Intergenic trnE-trnT |
| ucd4 | FAM-TTATTTGTTTTTGGTTTCACC TTTCCCATAGAGAGTCTGTAT | (T)12 | 92–95 | 4 | 45 TD | AJ489837, Intergenic ycf6-psbM |
| Reference (Origin of the Material) | Alleles at Locus ucd4, bp | Size Shift 1 | Alleles at Locus udt4, bp | Size Shift 1 | |||||
|---|---|---|---|---|---|---|---|---|---|
| Our study (Lithuania) | 92 | 93 * | 94 | 95 | Reference. | 141 | 142 * | 143 | Reference |
| Gailing et al. 2007, (north-western Germany) | 93 | 94 * | 95 | 96 | +1 bp | 143 | 144 * | 145 | +2 bp |
| Neophytou and Michiels 2013 (south-western Germany) | 92 | 93 * | 94 | 95 | 0 bp | 142 | 143 * | 144 | +1 bp |
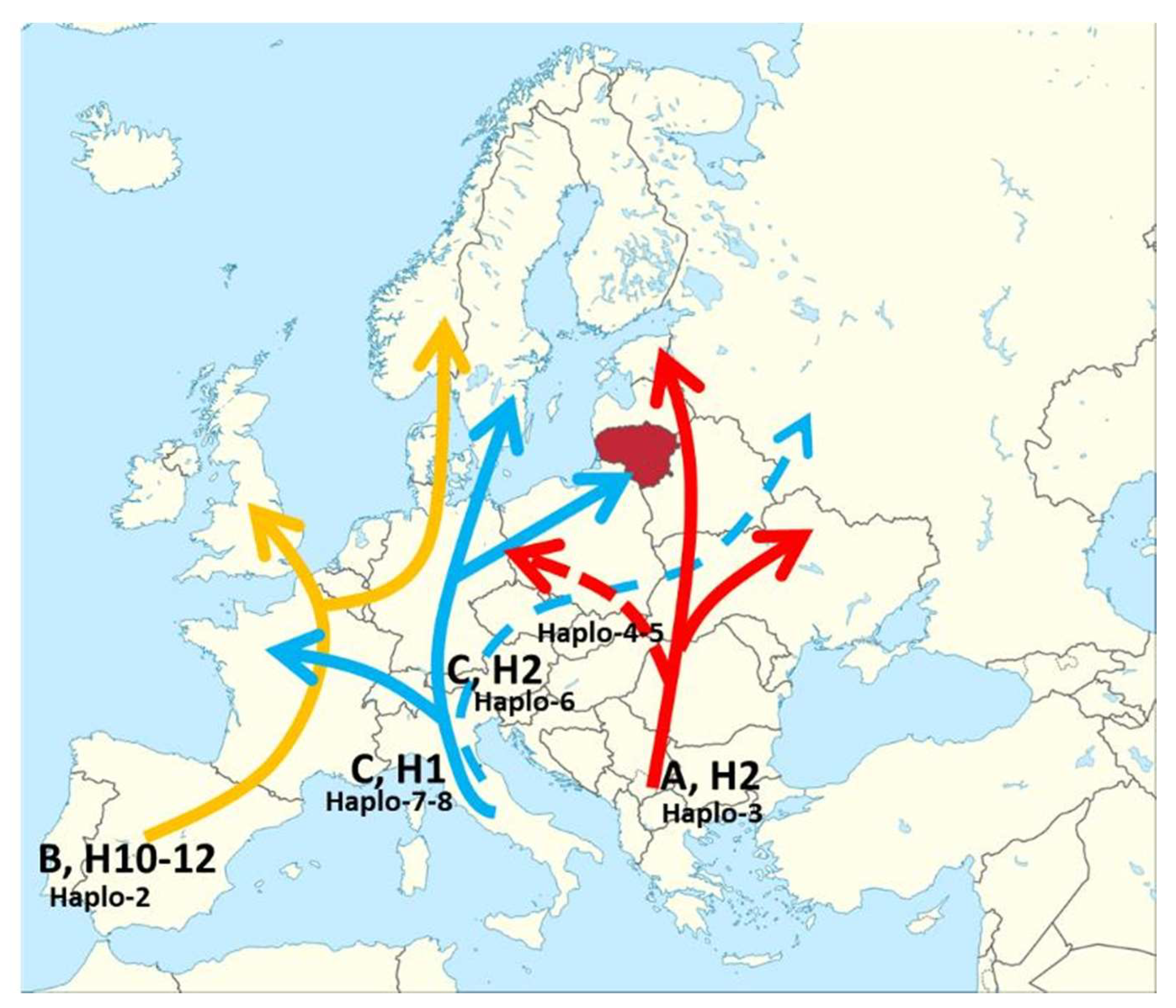
| Haplotype | Frequency | ucd_4 Allele Size, bp | udt_4 Allele Size, bp | udt_1 Allele Size, bp | Postglacial Migration Lineages and Haplotypes Based on [14,19,20,23] |
|---|---|---|---|---|---|
| Haplo-1 | 1 | 93 | 141 | 81 | [20,23] not found |
| Haplo-2 | 4 | 94 | 141 | 80 | H10-11-12 (lineage B) common SP, FR found in north DE and eastern SE |
| Haplo-3 | 117 | 93 | 142 | 81 | H5 ir H7 (lineage A) north-western Balkans |
| Haplo-4 | 7 | 94 | 142 | 80 | H4 (line A) post glacial mutation common in DE PL, HU, RO |
| Haplo-5 | 11 | 94 | 142 | 81 | H4 (lineage A) post glacial mutation common in DE PL, HU, RO |
| Haplo-6 | 10 | 92 | 143 | 81 | H2 (lineage C) eastern branch of Italian refugium |
| Haplo-7 | 1 | 94 | 143 | 80 | H1 (lineage C) western branch from Italian refugium |
| Haplo-8 | 6 | 94 | 143 | 81 | H1 (lineage C) western branch from Italian refugium |
| Source | df | SS | MS | Est. Var. | % |
|---|---|---|---|---|---|
| Among Pops | 30 | 27.0 | 0.90 | 0.123 | 30% |
| Within Pops | 127 | 36.1 | 0.29 | 0.292 | 70% |
| Total | 157 | 63.2 | 0.414 | 100% | |
| Stat | Value | p (rand ≥ data) | |||
| PhiPT | 0.30 | 0.0001 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Danusevičius, D.; Baliuckas, V.; Buchovska, J.; Kembrytė, R. Geographical Structuring of Quercus robur (L.) Chloroplast DNA Haplotypes in Lithuania: Recolonization, Adaptation, or Overexploitation Effects? Forests 2021, 12, 831. https://doi.org/10.3390/f12070831
Danusevičius D, Baliuckas V, Buchovska J, Kembrytė R. Geographical Structuring of Quercus robur (L.) Chloroplast DNA Haplotypes in Lithuania: Recolonization, Adaptation, or Overexploitation Effects? Forests. 2021; 12(7):831. https://doi.org/10.3390/f12070831
Chicago/Turabian StyleDanusevičius, Darius, Virgilijus Baliuckas, Jurata Buchovska, and Rūta Kembrytė. 2021. "Geographical Structuring of Quercus robur (L.) Chloroplast DNA Haplotypes in Lithuania: Recolonization, Adaptation, or Overexploitation Effects?" Forests 12, no. 7: 831. https://doi.org/10.3390/f12070831
APA StyleDanusevičius, D., Baliuckas, V., Buchovska, J., & Kembrytė, R. (2021). Geographical Structuring of Quercus robur (L.) Chloroplast DNA Haplotypes in Lithuania: Recolonization, Adaptation, or Overexploitation Effects? Forests, 12(7), 831. https://doi.org/10.3390/f12070831