1. Introduction
Coffee is one of the most popular beverages, consumed by about one-third of the world’s population; it is also one of the most commonly traded agricultural commodities worldwide [
1]. The world coffee trade is based on two species,
Coffea arabica L. (Arabica coffee) and
C. canephora (Robusta coffee), which account for 99% of coffee production worldwide. The crop is grown in approximately 80 tropical countries and it is estimated that about 25 million families worldwide are coffee producers, mostly smallholders and families whose livelihoods largely depend on this crop [
2,
3]. Coffee production is a significant contributor to agricultural gross domestic product (GDP) and export earnings in Latin America [
4]. In Costa Rica, coffee is recognized for its high quality and, as a “social crop”, it has a direct impact on the socioeconomic development of the country, representing 0.29% of the national GDP [
5]. Coffee production covers more than 25,000 ha [
6] and impacts more than 40,000 families directly, while thousands of people participate in associated processes, such as harvesting, processing, and marketing.
The intensification of extreme events under global warming scenarios threatens the sustainability of agricultural production on a global scale, with consequences on the amount and quality of harvestable crops for the current production areas, with coffee being no exception [
7]. Coffee is considered a crop that is sensitive to climate change [
8,
9] and particularly to the impact of temperatures (as its optimum temperature range is 18–25 °C) [
10]. The impact of climate change on coffee production has been analyzed for the main producing regions [
11,
12]. General projections suggest reductions in productive areas of up to 88% by 2050 [
13] and losses (and drift) of suitable areas in most coffee-producing countries [
3,
4]. Yield reductions, lower bean quality, and even higher pressures for disease development (e.g., coffee rust) [
14] are already being cited as causes of major economic losses [
15,
16]. The impacts described so far are often attributed to the inherent sensitivity of coffee physiology to temperature, rainfall, and CO
2 changes [
3]. As coffee originated in the shady understory of North African forests, it is considered a shade plant and such sensitivity to temperature is expected.
Common strategies that producers have adopted to adapt coffee crops to changing climate conditions include the introduction of new varieties, modifications of the field layout, and the use of overstory vegetation for shading [
17]. The use of new genotypes is challenging, as they must be productive without neglecting quality parameters [
18] and must show stability across environments. Shading, on the other hand, an ancient practice, is likely a more suitable practice, as shade buffers the microclimate and temperature fluctuations [
19]. Shading improves WUE because it modulates stomatal conductance and protects the soil from direct solar radiation, reducing soil temperature and water evaporation [
20]. Despite its potential impact on fruit load, shading is considered beneficial when kept around 30% (leaf area index of shade < 1), [
21,
22], which adds value to the production given the associated additional ecosystem services [
21,
23].
To better understand the effects of climate change and variability on coffee production systems, we need to face the limitations imposed by data scarcity. At this point, meteorological data to link environmental dynamism and climatic changes to crop productivity are scant, and it is crucial to upscale the analysis to include the physiological responses to environmental changes. To fill these gaps, a multi-scale approach is necessary so that the soil–plant–atmosphere continuum (SPAC) can be used as a framework to understand the impacts of climate and the feedback at the crop scale, and thus to enhance our capacity to deliver an integral model of climate-smart agriculture to producers. In this prospective article, we use a case study of a sun-grown coffee field to set up a basis for the design and study of resilient coffee agroecosystems, with an emphasis on the SPAC, the decoupling mechanisms between plants and the atmosphere, and their impact on ecosystem performance and yield stability.
Here we present the theoretical framework behind our study (
Section 2) and use preliminary results of the first year of measurements from the experimental field setup (
Section 3) to establish important considerations for this long-term experiment and the potential uses of our data. Although the current perspective was only focused on a sun-grown coffee field, the same setup will be implemented in a shade-grown coffee field in the near future so that the impact of shade on WUE and the coupling between plants and the atmosphere can be addressed (
Section 4).
3. Preliminary Results
In the field, we used the eddy covariance (EC) method and focused on the quantification of the water and carbon balance, as well as the estimation of evapotranspiration, in order to determine the general components of the energy balance and their associated fluxes, RN, H (latent Heat), E, S (Soil heat flux), and F
C (Carbon assimilation), in this case, E is estimated based on the surface fluxes balance approximation. For the partitioning of canopy and soil surfaces, the EC was based on high (temporal) resolution micrometeorological data at the crop scale. In such a context, it is well-known that this kind of coupling can modulate processes ranging from surface environmental conditions (e.g., temperature and precipitation) to plant growth and development [
31].
Partitioning the contributions of crop canopy and soil surface to H, E, and FCO2 requires the deployment of instrumentation at strategic sampling points. A variety of eco-physiological techniques (water relations, sap flow, and gas exchange) are available to assess each of the contribution to the E of the whole field. Observations of this kind are important to understand the effects of soil conditions on crop microclimate, water use, biomass production, and yield. Thus, the observational setup described here was composed of an arrangement of instruments to measure key variables in the canopy, leaves, and soil components of the SPAC.
3.1. The Study Site
We used a 30-year-old coffee plantation (renewed by full pruning 5 years ago) to assess the effect of coffee management on decoupling from the atmosphere. Different aspects were assessed: quantifying the annual cycle of mass, energy and water balances, theoretical considerations, and the practical deployment of instruments at several scales of observation. This entailed simultaneously measuring the system using a combination of micrometeorological and eco-physiological techniques across the SPAC.
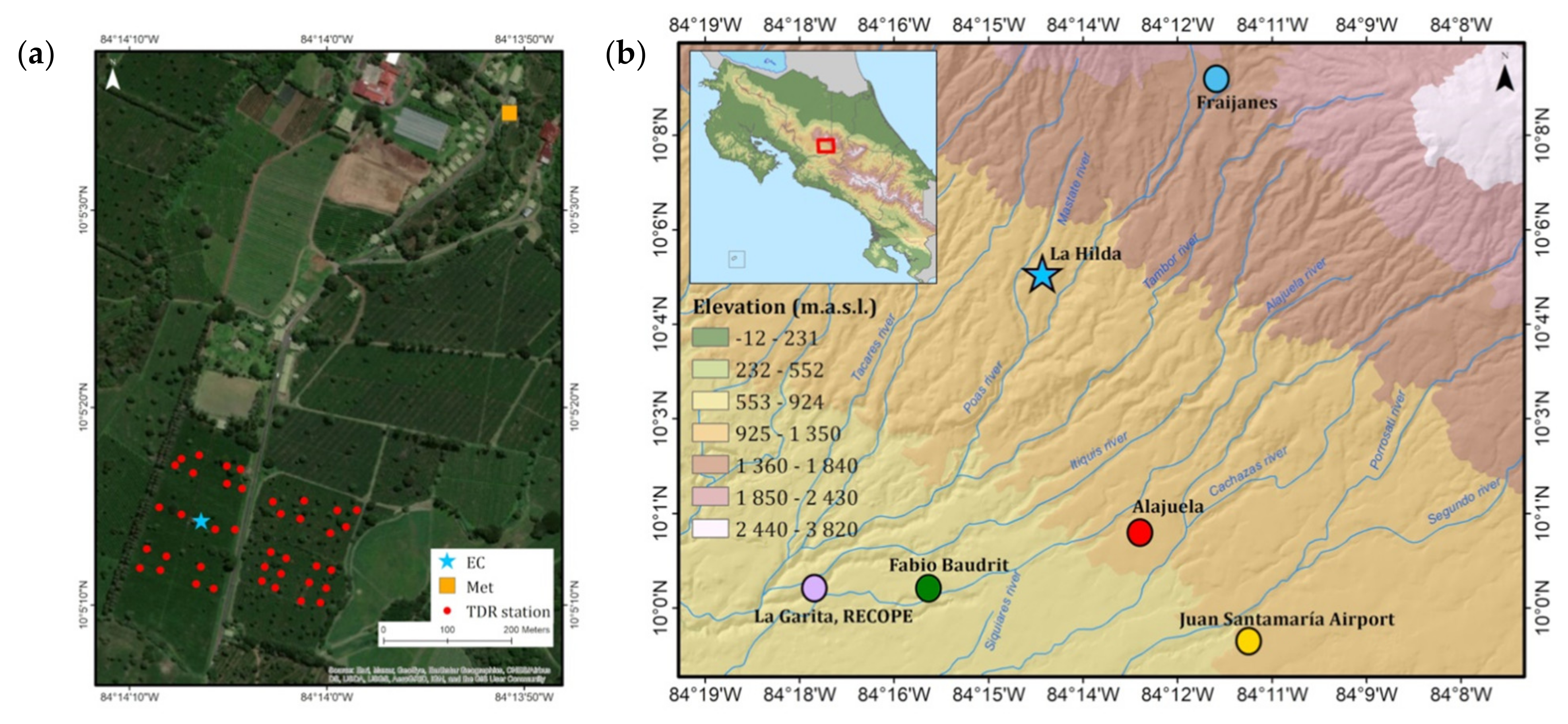
The study site was located at Finca La Hilda in San Pedro de Poás, Aajuela (10.0893″ N 84.235088″ W), Costa Rica (
Figure 2), between 1000 to 1500 m.a.s.l., with optimal conditions for coffee cultivation. The soils are mostly Andosols. The farm has coffee fields with different varieties (Catuaí, Caturra, Costa Rica-95, Obatá).
3.2. Long Term Climatology
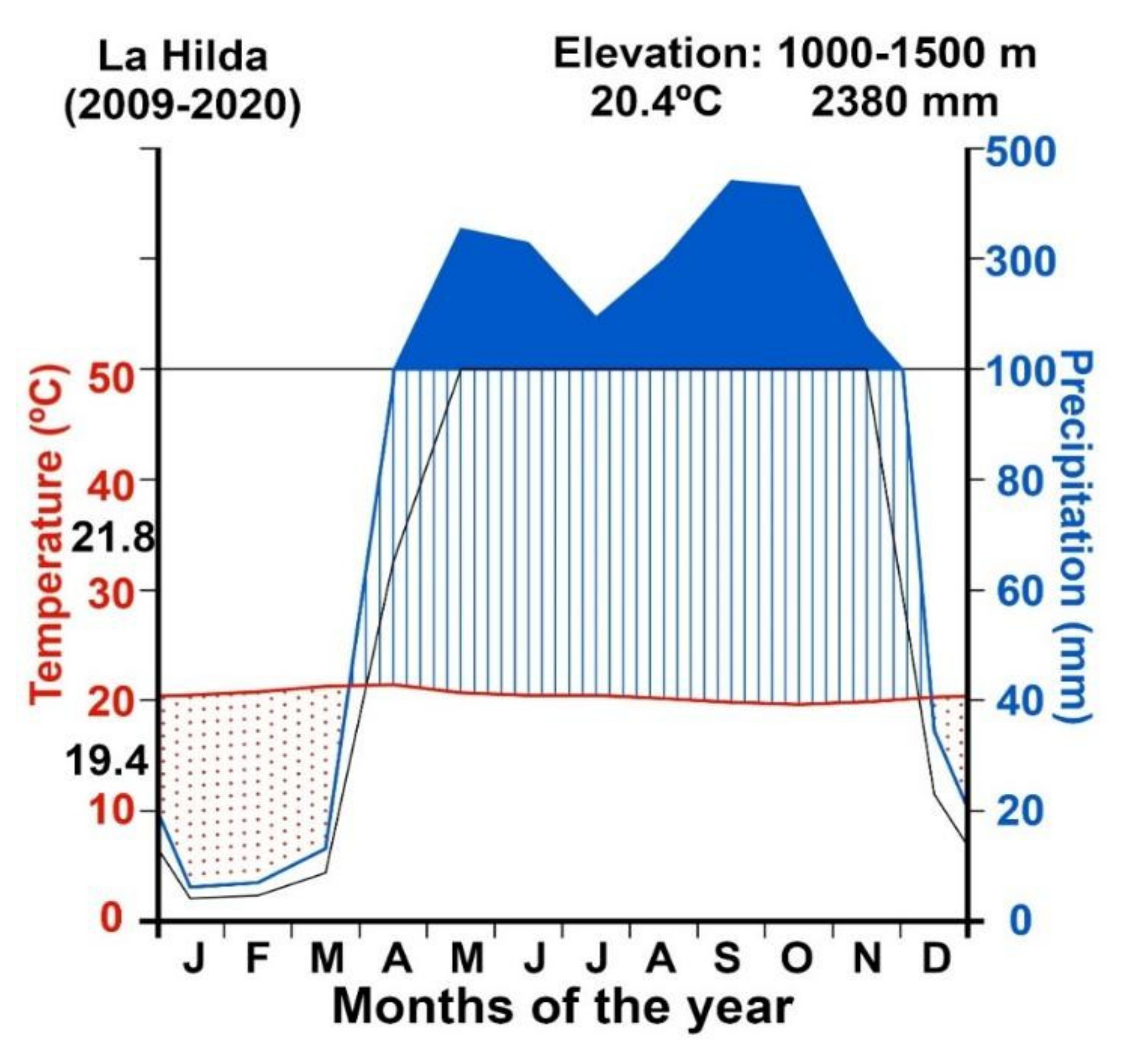
The site is located to the west of the Central Valley of Costa Rica, under strong influence from the ITCZ, with a pronounced dry season (December to April) and a bimodal rainy season (May to November), interrupted in July by the MSD (
Figure 3). The average total annual rainfall is 2380 mm. The driest month is January (<15 mm) and the rainiest is October (>500 mm). The average temperature is 20.4 °C with little annual variation. Rainfall and temperature patterns are the result of the interaction of low-level winds with local topography and the influence of the main synoptic scale systems that contribute to regional rainfall [
32].
The experimental site is influenced by an elevation gradient on the slopes of the Poás volcano. Records of average environmental conditions in the farm are provided by a Davis vantage automatic weather station. The analysis of the long-term climate was based on reference sites at the Fabio Baudrit experiment station (EFB, 10°01′ N, 84°16′ W, 840 m.a.s.l) and Fraijanes experiment station (10°05′ N, 84°16′ W, 1500 m.a.s.l); both sites belong to the University of Costa Rica and are equipped with Campbell automatic meteorological equipment from the National Meteorological Institute (
Figure 2b). Additional stations across the Alajuela province were also considered for the analysis (all sites shown in
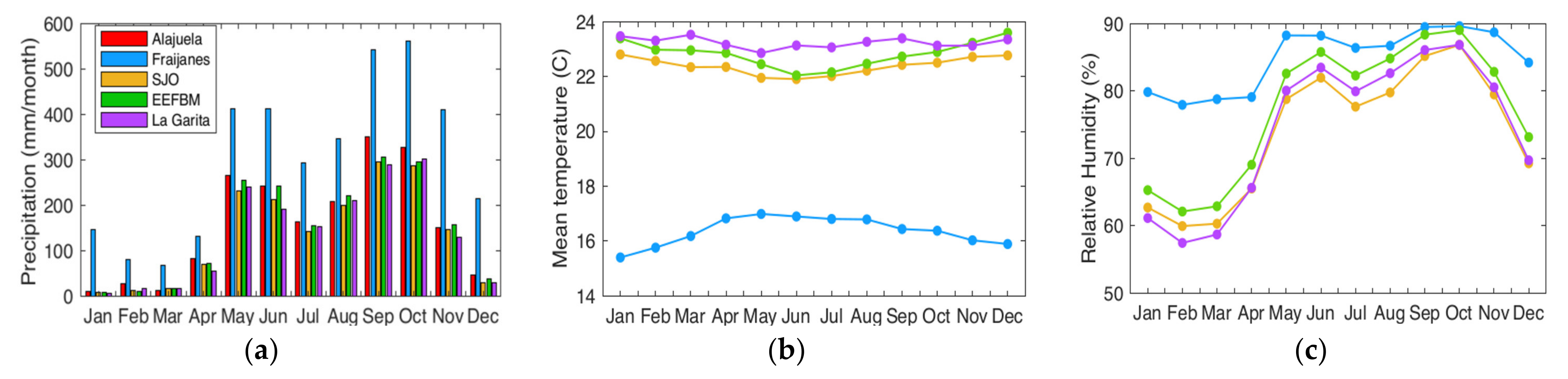
Figure 2b). The 1999–2019 hourly data were used to generate averages for the annual cycle of monthly accumulated rainfall (
Figure 4a), mean temperature (
Figure 4b), and relative humidity (
Figure 4c) for 5, 4, and 4 stations, respectively. Rainfall had a marked bimodal distribution (
Figure 4a) characteristic of the influence of the so-called midsummer drought (MSD) [
33]. The mean temperature was 21–24 °C, with higher temperatures in the drier months for the lower elevation site. This pattern was reversed at the higher elevation site at Fraijanes (
Figure 4b), where temperature increased after the onset of the rainy season and declined as the dry season was established at the end of the year. The relative humidity at Fraijanes was higher by approximately 20%, which implies that a larger amount of moisture was available in the system.
3.3. Coffee Field Structure
Sun-grown coffee fields in the experimental area were restructured to increase light interception, thus improving radiation use efficiency, and to allocate more biomass, thus providing higher and more stable yields. This was achieved by substantially increasing inter-row spacing compared with more traditional spacings, whereas the distance between plants was significantly reduced (
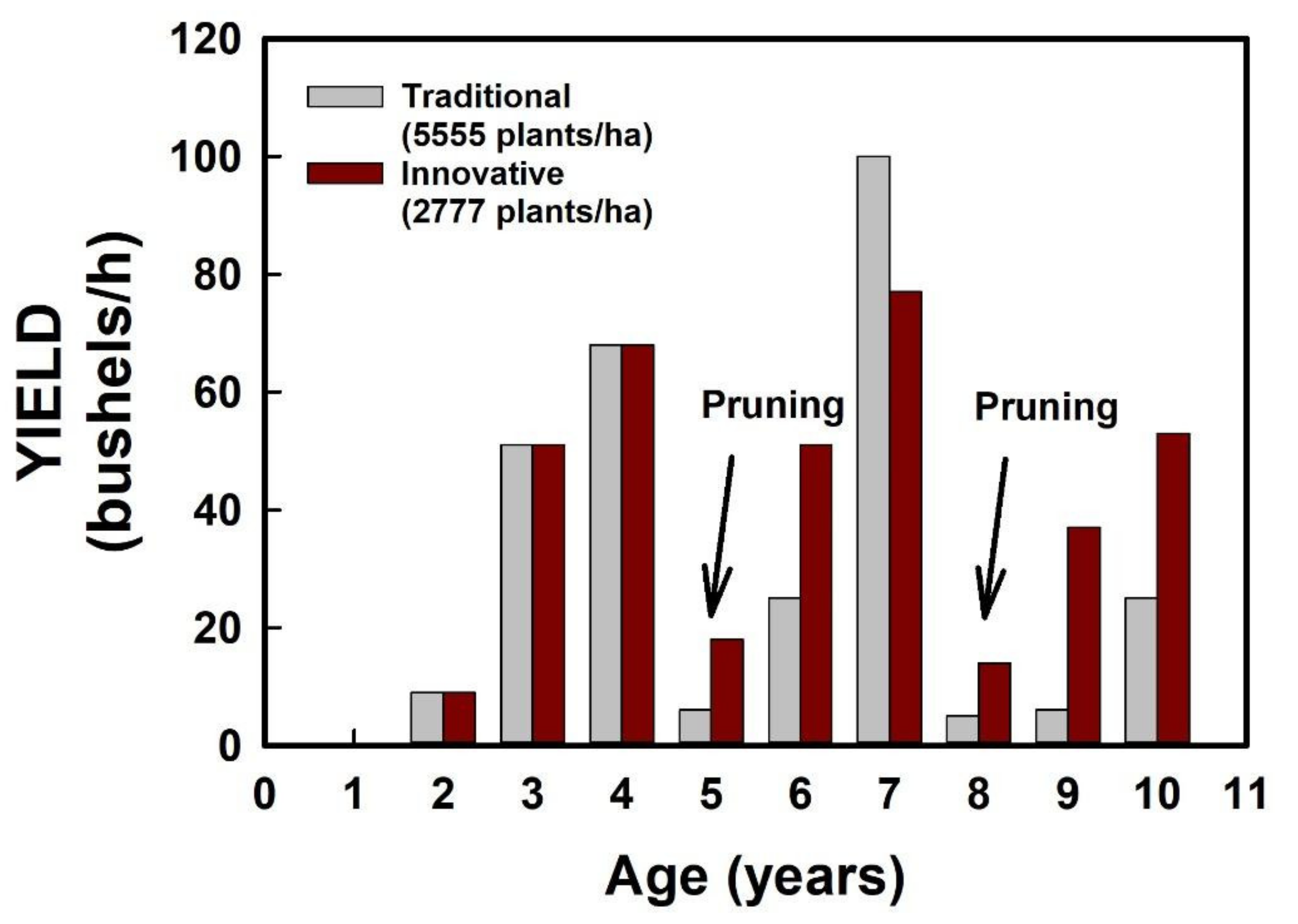
Figure 5). This resulted in substantial differences in plant density, with 5555 vs. 2777 plants/ha in the traditional vs. innovative agroecosystems, respectively. Pruning strategies were also modified by introducing training techniques and frequent pruning of plagiotropic branch tips (i.e., “hedging”) to promote lateral branch growth and a long-lasting tigmo-morphogenetic response that could easily reconstitute the hedgerow leafy surfaces after every pruning pass.
The result was a re-distribution of the LAI and the development of dense, homogeneous leafy walls that extended virtually all the way down to the soil surface and displayed high and uniform photosynthetic rates and high yields, comparable to those obtained with conventional densities (
Figure 5). Moreover, our long-term results showed that mature, fully productive coffee fields (hedged fields) recover faster, especially after the third pruning, and their yields remain more stable than in fields subjected to conventional, more severe pruning of the whole shoot (
Figure 6).
3.4. Surface Fluxes
As previously mentioned, the analysis of mass and energy balance requires higher frequency information (ca. 10 Hz or higher) and a larger number of variables. The experimental setup considered in this experiment to monitor the lower atmosphere was integrated by an automated weather station and an eddy covariance (EC) system, amplified with measurements at the leaf level (leaf temperature, D, GS, and GBL), as explained below. A closed path Li-7200 EC system (LI-COR, NE) was deployed in the sun-grown coffee field. EC was used to compute the mass and energy budgets to estimate evapotranspiration (latent heat, E), net CO2 fluxes (FC), gross primary productivity (GPP), ecosystem respiration (RECO), and, most importantly, the carbon balance of entire fields during different climatic conditions.
3.4.1. Annual Cycle of Surface Fluxes
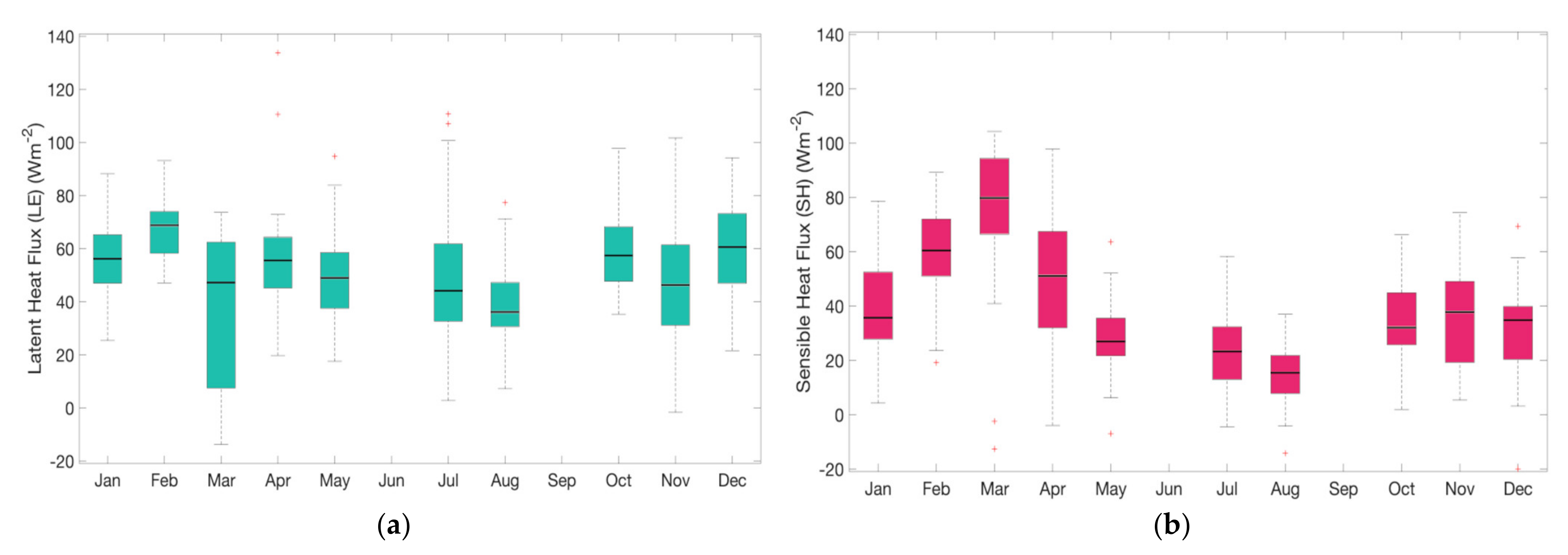
Here we present the preliminary results of the EC-derived measurements for 2020, for latent and sensible heat flux, and for evapotranspiration (
Figure 7). Due to power issues in the field, the flux system was not continuously working during April and September, so the data for those months were excluded, given that missing data exceeded 15%, which was the missing data threshold used. Quality assurance and quality control of data were carried out according to Foken et al. [
34].
During 2020, latent heat flux (LE) presented a monthly average of nearly 40–70 Wm
−2 with significant daily variability (
Figure 7a). The largest daily variability was observed for March, the month in which the transition to the rainy season starts (
Figure 7c). Surface–atmosphere coupling was expected to show major activity during this transition period. As the rainy season ends, soil moisture availability declines and one of the responses tends to be a decline in the latent heat flux, given that the area has previously experienced high evapotranspiration rates (
Figure 7d).
The sensible heat flux (H) annual cycle (
Figure 7b) showed variations of 20–80 Wm
−2 and peaks towards the end of the dry season. In this period of warmer conditions, vertical temperature gradients are maximized by the contrast between the hot surrounding air and the cooling effect that coffee transpiration has on the surfaces underneath (the soil and lower leaves). As the rainy season was established in April (
Figure 7c), H started to decrease and a reduction in the temperature gradient was to be expected. The secondary peak of H was approximately 50% of the value for the first peak and this suggests that surface–atmosphere coupling differed during the two peaks of the rainy season. Furthermore, the surface plays a major role in the establishment of the first half of the rainy season, in contrast with the larger (synoptic)-scale force that dominates the second, stronger half of the rainy season.
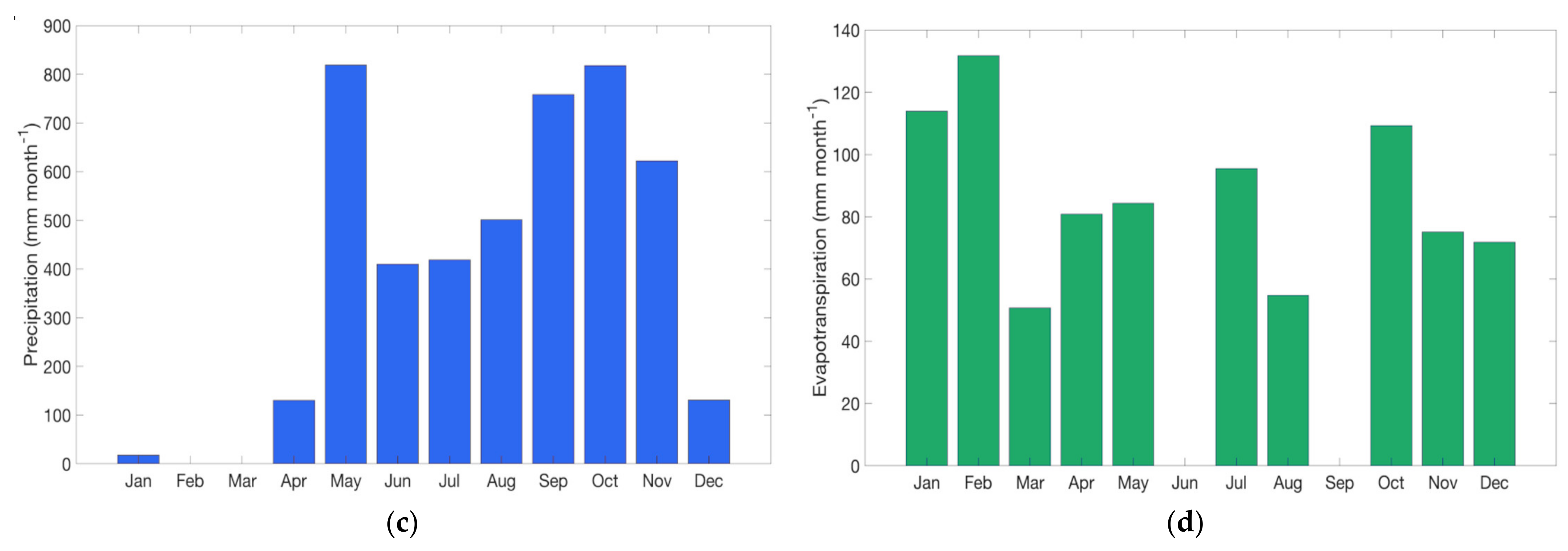
Water flux is dominated by precipitation which shows an annual cycle featured by the climatological dry and wet seasons characteristic of the region while E shows the response to variations in precipitation (
Figure 7d). Monthly cumulative E indicate that it accounted for approximately 25% of the total rainfall (April and September were not considered in the estimation). E was largest during the dry season, showing a large net rate of surface water loss, in agreement with the annual cycle of the surface fluxes (LE and H). After the onset of the rainy season, surface water availability was enough to maintain high soil water content despite the high E rates observed. Hence, the severity of crop water stress was at a maximum during the dry season and prior to the establishment of the rainy season.
3.4.2. Annual Cycle of CO2 Fluxes and Productivity
If surface fluxes provide a measure of the connectivity between the surface and the atmosphere, CO
2 flux and GPP are essential to explain the dynamics of C capture, long-term ecosystem carbon balance, and yield production. On average, the sun-grown coffee field behaved consistently as a CO
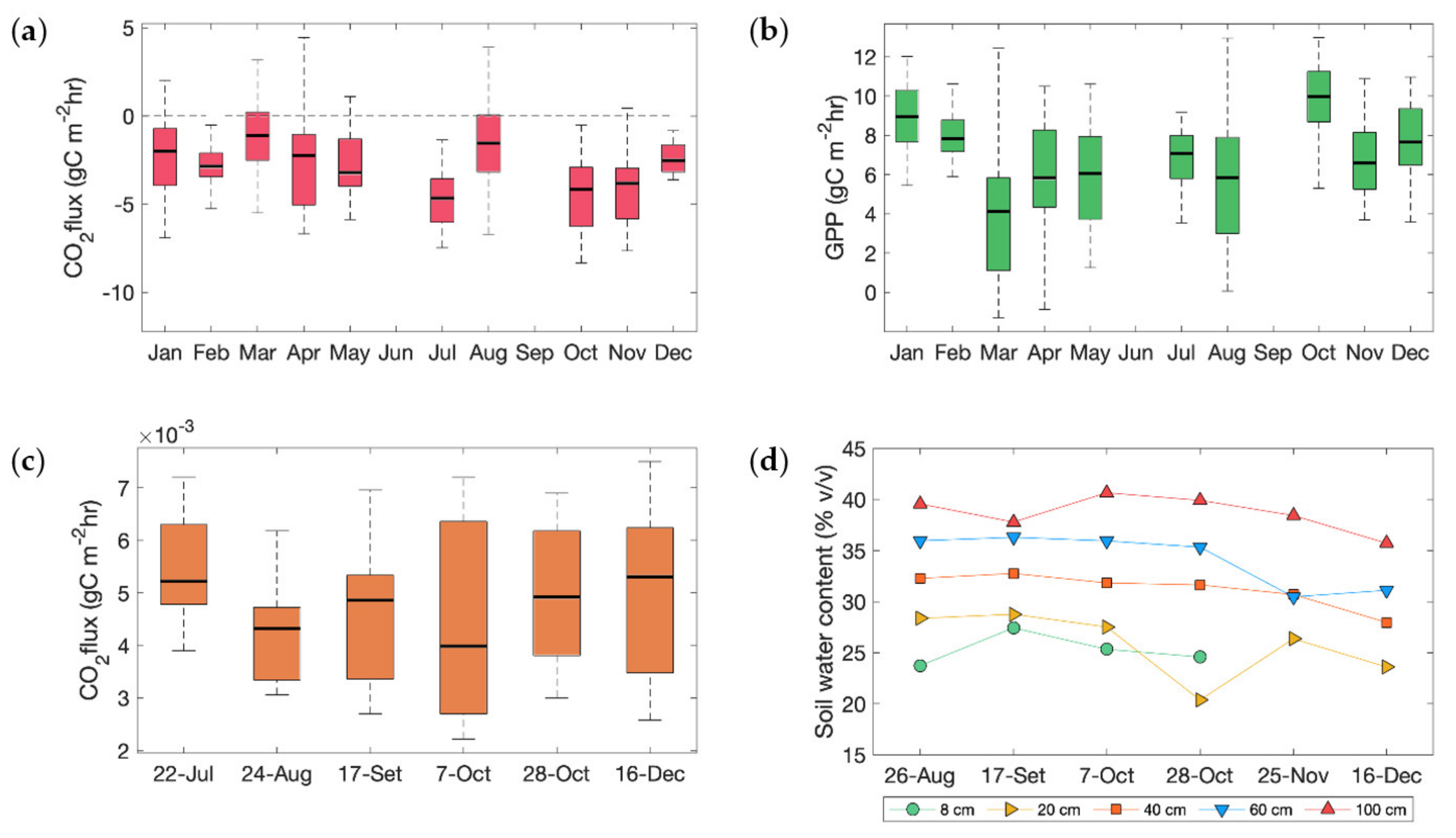
2 sink during the year (
Figure 8a). CO
2 fixation was enhanced during the second half of the rainy season, consistent with the high rainfall and high soil water content. However, such behavior was not observed during the first half of the rainy season. The annual cycle of GPP (
Figure 8b) shows that productivity also peaked during the second half of the rainy season, coinciding with the most active phase of growth and storage of the developing fruit crop, which is known to cause increases in the photosynthetic rates of leaves in response to increased sink demand for assimilated carbon [
35]. This implies that CO
2 capture dynamics could be driven by coupling of the surface and the atmosphere at higher levels.
3.5. Soil CO2 Fluxes and Water Content
Soil CO
2 efflux accounts for the 50–70% of ecosystem respiration (RECO) [
36] and is highly variable at the temporal and spatial scales due to the complexity of the different components that contribute to this flux (i.e., ground surface organic litter, live roots, and soil organic matter pools) [
37]. In our experimental setup, the strategic deployment and placement of stations for soil measurements (soil water content, CO
2) allowed the separation of soil fluxes from whole coffee field fluxes. Soil CO
2 flux measurements provide information about the factors driving soil respiration (Rs) and aid the evaluation of the component processes in models, particularly belowground processes that are not entirely understood. Furthermore, data from chamber measurements are also useful to fill gaps in EC datasets, particularly when a (large) representative portion of the ecosystem is included in the measurements [
36]. The Rs rate (the sum of autotrophic respiration by roots and heterotrophic respiration by microorganisms) is also affected by the other elements that influence microbial activity, such as temperature, soil moisture [
20,
38,
39], organic matter, and the supply of photoassimilates from the shoot (determined by plant type, phenology, and assimilation rate). Thus, soil CO
2 measurements are highly responsive to linkages and feedback between above- and belowground components in the SPAC [
40].
For soil flux measurements, CO
2 fluxes are measured using a closed chamber system with a Li-Cor 8100 (Li-Cor Inc., Lincoln, NE, USA). A total of 20 PVC anchors were installed in the soil at depth of 12 cm, following standard recommendations for an expected error acceptance of 10% [
41]. Fluxes were measured monthly from July 2020. CO
2 measurements were carried out with a chamber deployment of 90 s and three repetitions per anchor, and fluxes were calculated with the standard flux equation using SoilFluxPro™. For each sampling point, volumetric water content was determined at five different depths (8, 20, 40, 60, 80, and 100 cm) with a HandiTrase TDR Soilmoisture Meter, model 6085K3 (Soilmoisture Equipment Corp., Santa Barbara, CA, USA).
Soil at La Hilda was expected to present a good water-holding capacity due to its volcanic origin; accordingly, soil moisture did not change dramatically throughout the measurement period (
Figure 8b), which allowed for microbial activity and other processes responsible for soil respiration to continue relatively unchanged during the rainy season. The preliminary results showed a maximum in July (
Figure 8a) and little variation within the remaining months of the rainy season. As expected, our CO
2 results showed high spatial variability (coefficient of variation (CV) of 20.95–39.87%), which is usually explained by spatial heterogeneity in soil water, temperature, and organic carbon content [
20]. The spatial variability for the dataset tended to increase towards the end of the year, which was perhaps linked to variations in soil moisture (
Figure 8b) that might have created responses in the CO
2 at the microscale level. Several recommendations have been made regarding the number of samples needed to achieve a required uncertainty [
42]; however, the number of chambers needed to reduce the uncertainty can be impractical and unrealistic for many ecosystems and field situations [
36], so new approaches were explored to reduce the variability in CO
2 flux for the site in the near future.
3.6. Sap Flow Measurements and Latent Heat Flux Partitioning
Whole plant transpiration can be estimated through sap flow sensors, which allow measurements of water use and actual transpiration in real time [
43,
44]. Because canopy transpiration (canopy latent heat flux) is estimated as E
C = G
CD, normalization of E
C by the leaf area distal to the location of the sap flow sensors yields the actual canopy conductance and transpiration rate. Simultaneous deployment of sensors to measure sap flow in thinner shoots of coffee and in thicker woody trunks of shade trees facilitates the simultaneous measurement of E
C components in sun-grown and shaded coffee fields. Sap flow measurements were carried out in six plants using the Dynamax Sap Flow sensors SGB 25 and SGB 35 (Dynamax Inc., Houston, TX, USA). In this method, a resistance is used to heat the stem and the heat fluxes that leave the system are calculated from measured temperature gradients using the energy balance method [
45].
Leaf gas exchange can be reliably measured in the field using steady-state automated porometers, which allow rapid and precise measurements of stomatal conductace. In this study, we used an AP4 Porometer (Delta-T Devices Ltd., Cambridge, UK). For the measurement at leaf level, a part of a leaf was enclosed at the base of a cup containing a humidity sensor and dry air was flushed through the cup until a preselected lower level of relative humidity was achieved, as described in detail by Montheit et al. [
46]. Usually, the plant is sectioned into shade and sun leaves and the newest fully expanded leaf is selected for the measurement.
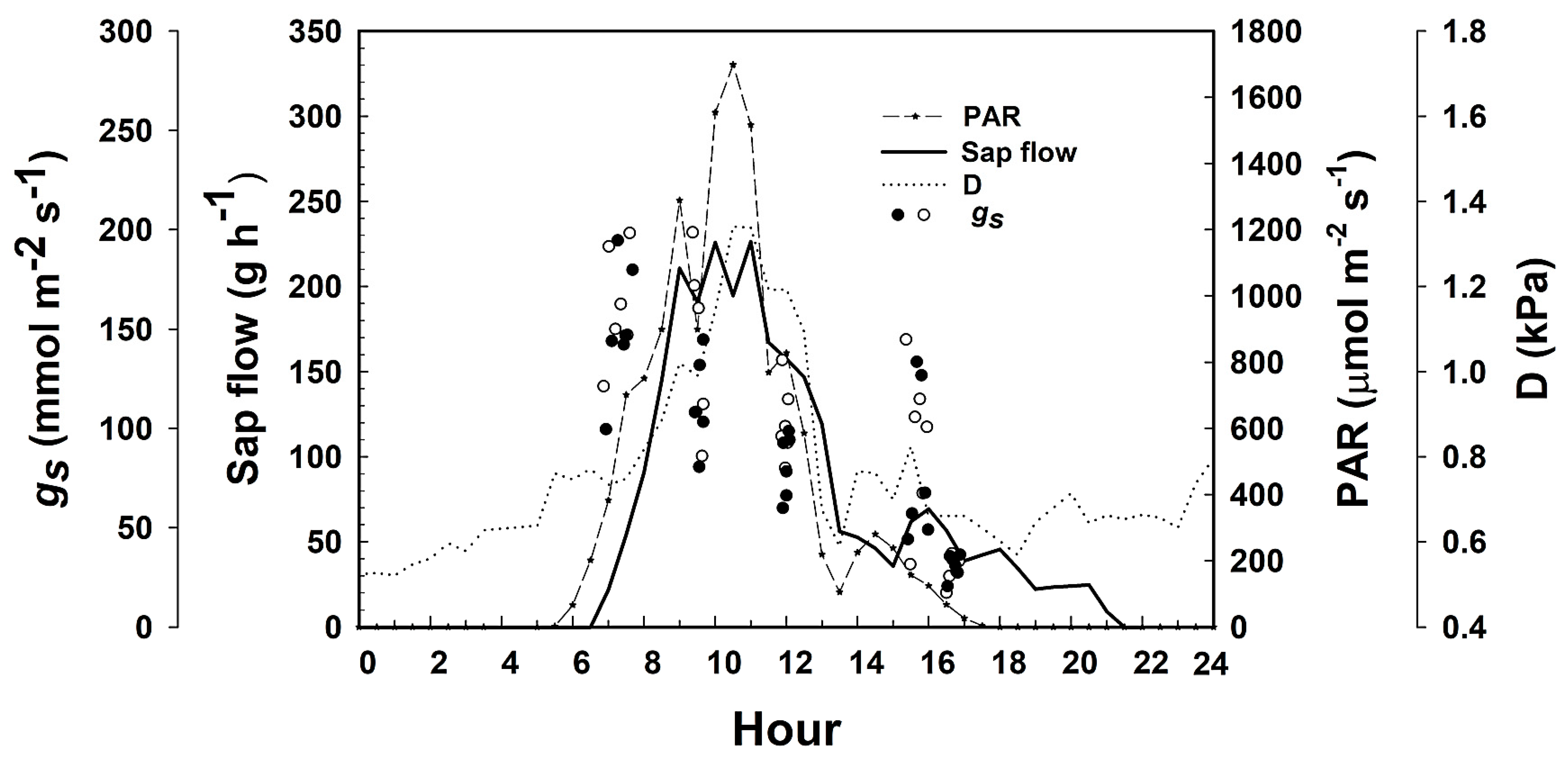
A day during the peak of the rainy season (
Figure 9) is presented here as an example of the typical daily course for the simultaneous measurements of sap flow and stomatal conductance in relation to environmental variables in the field in order to demonstrate the close dependance of transpiration on photosynthetic active radiation (PAR) and vapor pressure deficit (D). There was a flattening of stomatal activity around mid-day when PAR was close to 1500 umol m
−2 s
−1 and atmospheric conditions were drier. High D is likely to trigger direct stomatal responses to air humidity in coffee, resulting in stronger stomatal control of transpiration [
47]). The close correspondence of sap flow with PAR levels also indicates the reduced storage capacity of coffee plants and strong coupling between daily PAR availability and water use. It is important to point out that PAR and D have opposite effects on stomatal conductance throughout the day (see [
47]). However, the low flows observed between 1700 and 2200 h suggest that some stem recharge may have been taking place, or that nocturnal transpiration at high D during the early night hours was driving transpiration at this time. The transpiration rate can be obtained by normalizing the sap flow rate by the leaf area, while the canopy boundary layer conductance can be calculated using Ohm’s analogy along the plant–air interphase, as stomatal and boundary layer conductances are connected in series during the transpirational process. The data obtained were used to feed the canopy decoupling model described above in order to produce information about the relative importance of stomatal and environmental factors in the control of canopy fluxes.
3.7. Water Use Efficiency in the Agroecosystem
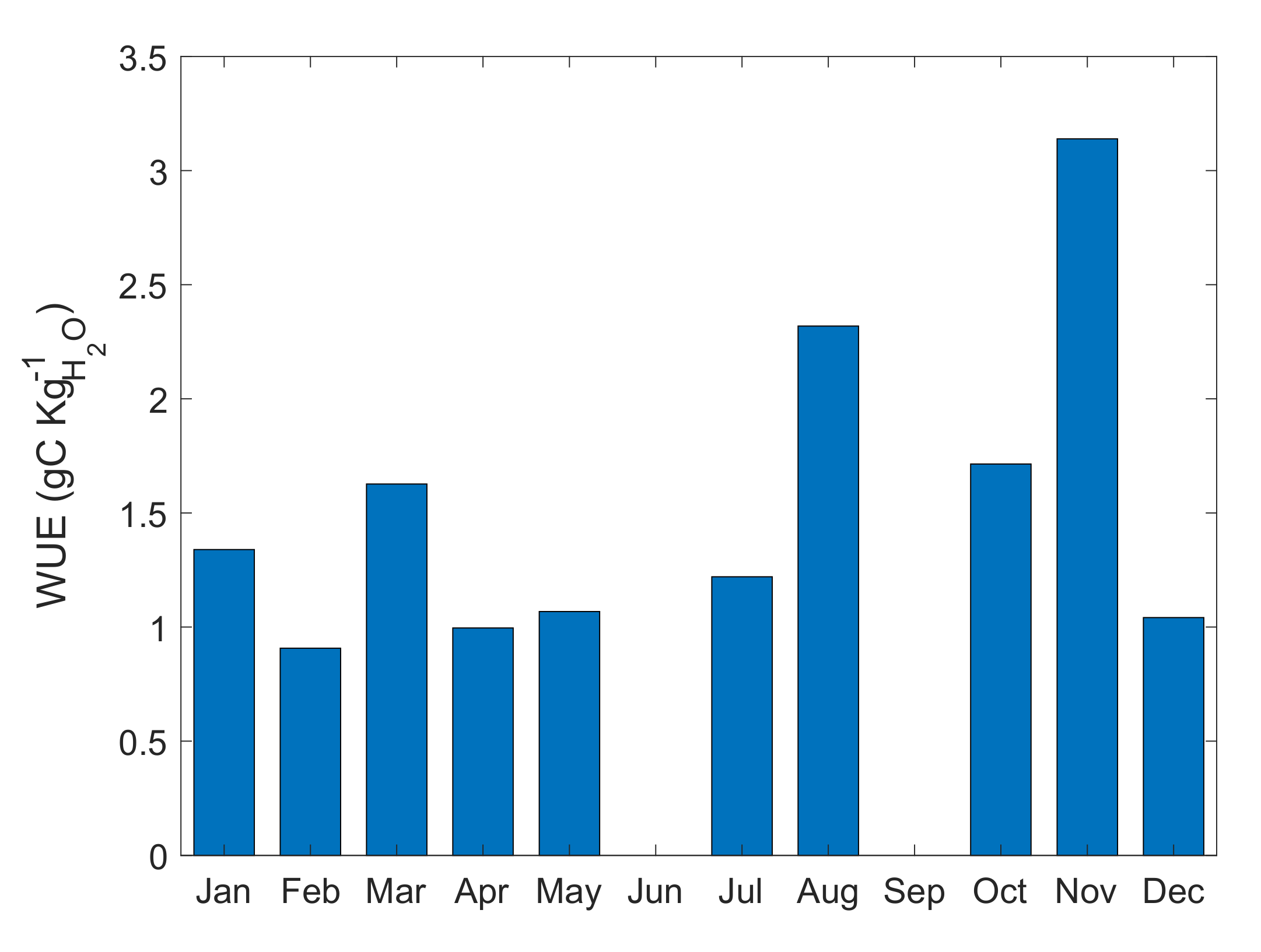
As one of the main concerns of this study was the impact of climate variability and change on coffee production, a link between the productivity in terms of water availability and use was established, given that the rainfall amount and seasonality are known to be affected by climate variability and change. WUE is related to GPP as it is modulated by water, C, and energy feedback between the canopy and the atmosphere. In this case, we used a traditional approach based on the EC-derived GPP and evapotranspiration, with the WUE defined as the ratio between the GPP and E (
Figure 10). As we were interested in the annual cycle of WUE, half hourly EC output was used to estimate daily GPP and E to generate monthly averages of WUE.
WUE varied between 1 and 3 gC Kg
−1 H2O, with largest WUE values for the months featuring larger numbers of heavy rainfall days. It increased prior to the onset of the first leg of the rainy season and during the second leg of the rainy season. The seasonal behaviour observed for WUE suggests that it is subject to larger intraseasonal variability as a result of the influence of heavy rainfall events for which the surface fluxes present the largest variations, which is consistent with other studies in coffee under full sun exposure [
48]. As can be noticed from
Figure 10, WUE does not exhibit the largest values as expected during the drier periods. In this case further aspects should be considered as the monitoring continues including the soil moisture storage, the effect of dew as a secondary water input in the system at the beginning of the dry season as well as the impact of irrigation. Such aspects are relevant to be considered in further research utilizing a longer period for the observational EC data, as future climate scenarios project a likely increase in the number of extreme precipitation events, so a direct impact on the WUE seasonal distribution may be expected as a result of warming scenarios.
4. Shading the Coffee Agroecosystem
It is expected that agroforestry systems will enable higher efficiency and the ability to complement the use of the resources available for the coffee plant [
41]. Thus, use of shade in coffee systems has been promoted as a strategy for mitigating and adapting to future climates, not only due to such systems’ abilities to regulate mean and maximum air temperature [
49] but also because shaded systems are expected to store more carbon than their sun-grown counterparts, thereby contributing to the reduction of greenhouse gases.
To assess the effects of shade trees on WUE and other fluxes in the investigated field site, an EC tower will be installed in a shaded coffee field. We hope to assess not only the existing shade of
Erythrina sp. (very common in coffee fields due to their N
2 fixation capacity) but also the effect of the arborization of a sparsely shaded coffee field through the employment of diverse native shade trees, planted to imitate the local forest structure and with consideration of natural succession and differences in growth form and longevity. Early successional species with rapid growth and high biomass production and turnover will be planted early (
Cecropia sp.,
Carica papaya. Musa sp.,
Trema sp., and
Acnistus sp., among others), in combination with slow-growing trees that will progressively reach maturity at different heights (
Inga sp.,
Gliricidia sp.,
Miconia sp.,
Croton sp., etc). Early agroecosystem arborization can also be assisted by planting “instant trees” (large cuttings; see [
50]). Tall, long-lived trees, like
Erythrina sp.,
Diphysa sp., and
Juglans sp., among others, will later constitute the emergent canopy layer. Shaded agroecosystems may include mixtures of significantly different species, producing a vertically stratified canopy (
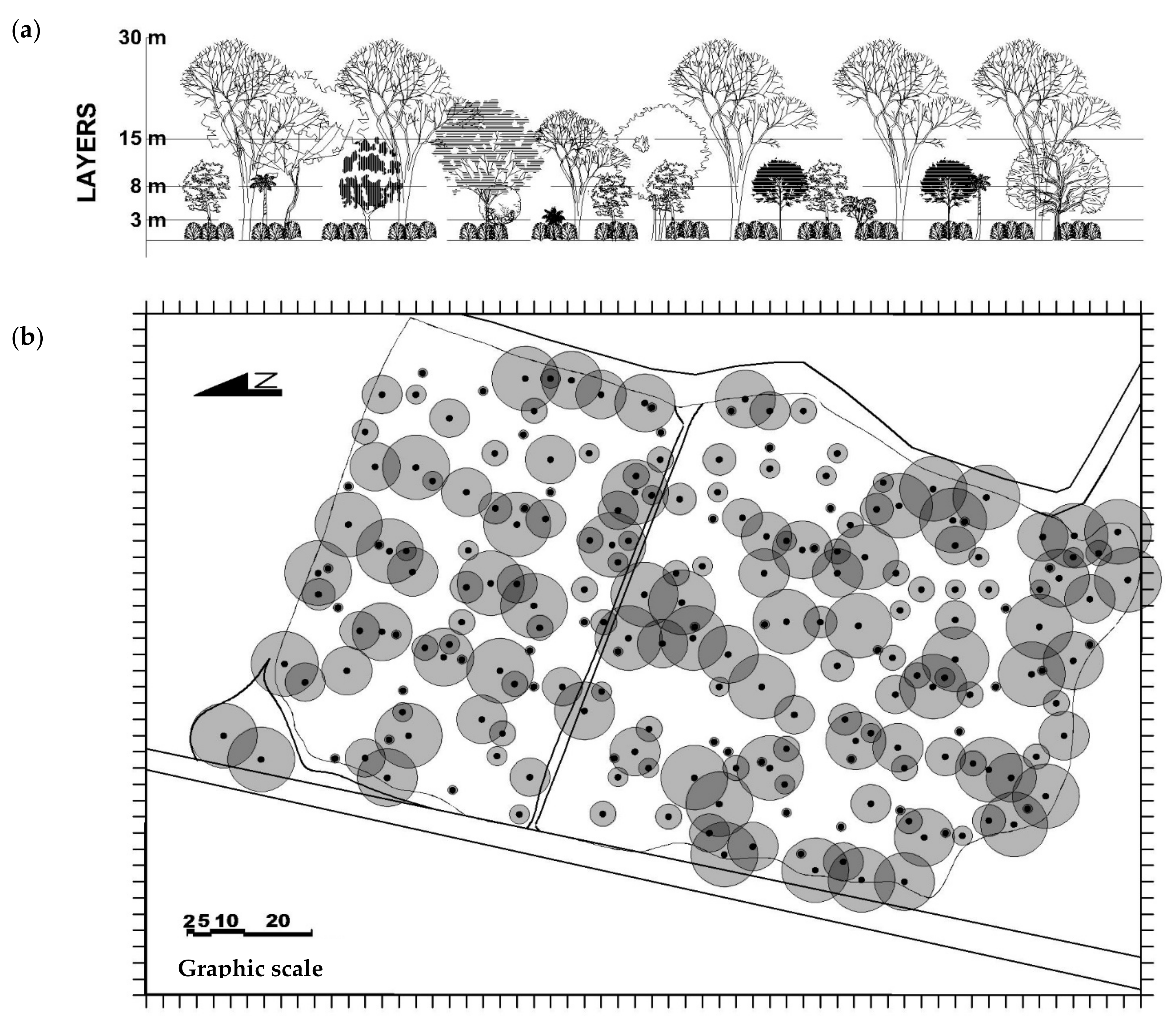
Figure 11).
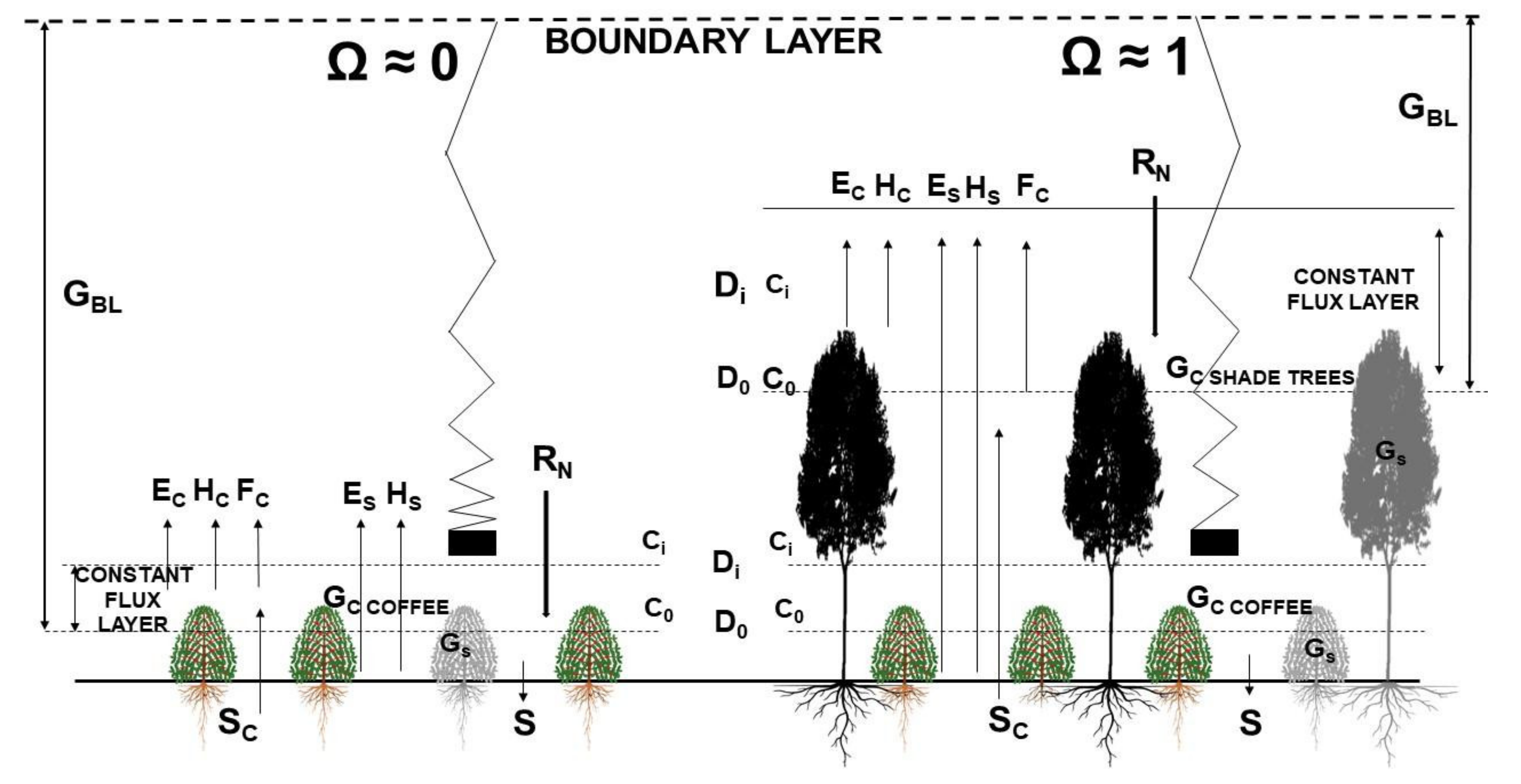
It is expected that shade trees will have a direct impact on WUE as tall shade trees employed as permanent, long-term components of agroforestry systems are aerodynamically rough, generate more turbulence than the short coffee monocrops (2.5–3 m), and have larger roughness lengths and low dragging coefficients [
25]. The wind speed experienced by shade trees is also higher because their tops are located higher in the wind speed profile, which results in a high
GBL and high coupling (low Ω) with the above-canopy air. Emergent shade trees within a coffee agroecosystem should be well-coupled to the atmosphere above and should easily exchange mass and energy. We argue that tall, overstory shade trees will be tightly coupled to the atmosphere, exert strong stomatal control on E and A, and use CO
2 efficiently, especially when compared with the low
GBL and high Ω of intensive sun-grown monocrops
(Figure 1).
The progressive development of the arborization of the shaded field, and the rapidly increasing shade provided by the fast-growing trees, is expected to progressively decouple the coffee plants from the bulk atmosphere and produce highly resilient, stable agroecosystems, supported, from a yield and climatic perspective, by quantitate carbon balance assessments.
5. Concluding Remarks
The preliminary analysis showed the strong influence of the local climate conditions on the partitioning of energy among the dominant fluxes in productive coffee fields. The data indicated that the sun-grown coffee described in this paper consistently functioned as a C sink, depending on the stage of canopy development, the phenology of the crop—especially the reproductive phase—and the prevailing climatic conditions. Canopy development and structure determine the LAI, light interception, and crop productivity, and they greatly affect the partitioning of the radiative flux between evapotranspiration, heat, and soil heat fluxes. Due to its comparatively small magnitude, FC is often excluded from energy balance analyses, but our results are crucial to the management and the design of coffee agroecosystems and the quest to achieve resilience, sustainability, and high-quality yields. It is reasonable to expect high dynamism in the capacity of the coffee crop to fix carbon, a probable result of the high phenotypic plasticity of this species, which allows its cropping under contrasting sun and shade conditions. For example, during more benign years with reduced water and high temperature stress (i.e., La Niña), we can expect coffee fields to behave as C sinks, while in drier, more stressful years (i.e., El Niño), coffee fields can be rather expected to behave as C sources and experience an overall negative C balance for the year. We can also expect crop canopy coupling with the atmosphere to vary seasonally and phenologically, with more decoupled fields during less stressful periods.
The experimental design allows the collection of fundamental data that can be leveraged for different applications within agrometeorology and agronomy. The future directions for this experiment comprise the comparison with the shade coffee field described in
Section 4 and measurement during a period long enough to account for both El Niño–Southern Oscillation (ENSO) phases in order to directly measure the impact of this variability mode for the SPAC coupling and its relationship with WUE and yield... Future directions also include contributions to the modeling community as we are seeking a suitable environment to improve and advance the development of a better boundary layer and hydrological parameterizations based on the observations.
The preliminary results indicate that the evaluation of crops in terms of C fluxes should be considered instead of the use of in situ measurements and that the role of the crop as a C sink should be incorporated to guarantee consumers a more environmentally responsible product. Given the scale of the crops in the Costa Rica and the orography, the performance of climate models is often under-discussion. Surface fluxes remain among the most challenging variables to model because of surface complexity. An initial modeling application on the site was developed by Pateromichelaki [
42], who used the surface observations to evaluate the capacities of the soil–vegetation–energy transfer (SVEN) model. The progress facilitates the integral comprehension of the SPAC and motivates the further improvement of surface parameterizations that could be beneficial for the development of regional climate models and also the validation of satellite-derived products.
The preliminary results of the first observed full annual cycle showed the marked seasonality of the surface fluxes and the strong response of the WUE to the establishment of the rainy seasons, as well as the intraseasonal variations in terms of the rainfall intensity of the events. Such results are promising for the exploration of further aspects of the SPAC feedback and of the interaction with rain-producing systems as potential elements to modulate the WUE and likely LUE in coffee plantations. It has been shown how relevant the deployment of this type of experimental setup is for leveraging knowledge on the SPAC and, moreover, for linking the crop with larger scale meteorological conditions. We expect to continue our experimental analysis in order to better inform models and contribute to the development of more robust frameworks to interpret the potential impacts of warming on coffee production.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}











