
Effects of CaO on the Clonal Growth and Root Adaptability of Cypress in Acidic Soils

Abstract
:1. Introduction
2. Materials and methods
2.1. Study Site and Selection of Materials
2.2. Experimental Design
2.3. Measurement Indices
2.4. Data Analysis
3. Results
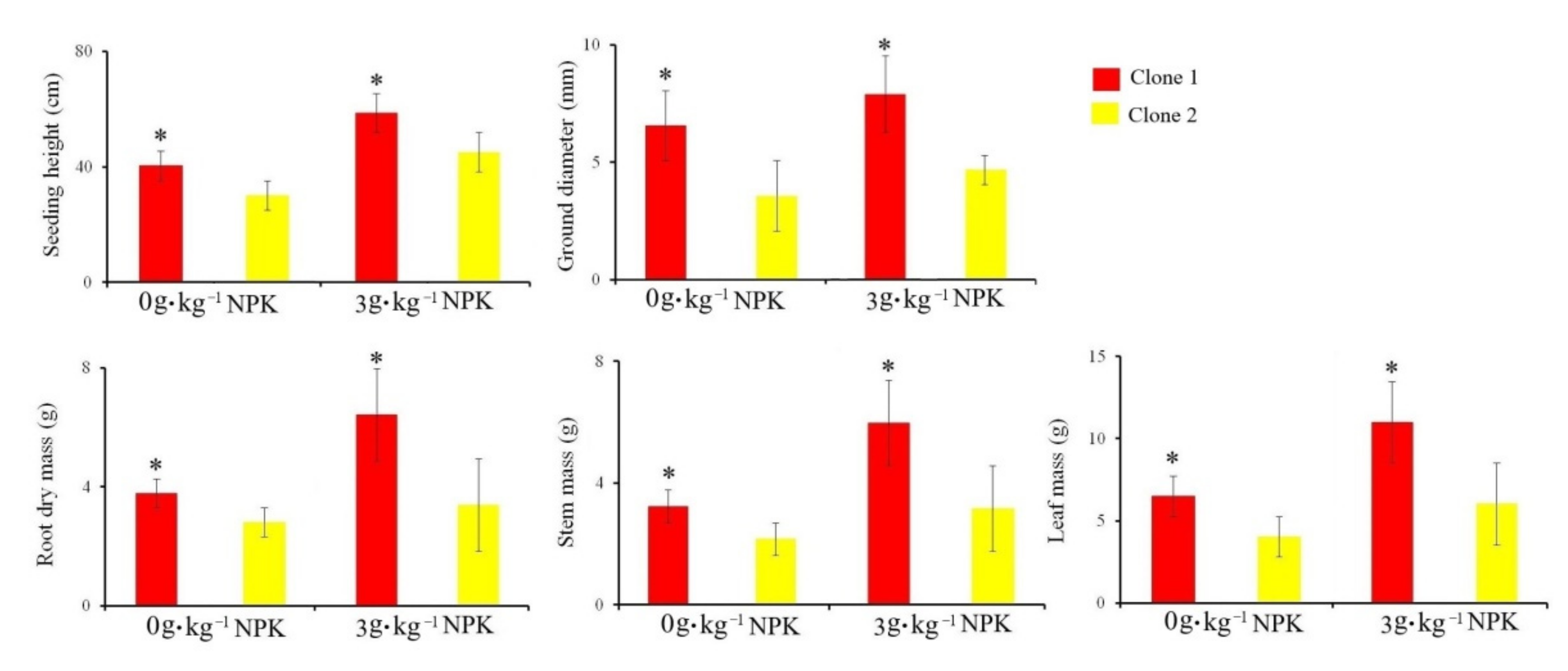
3.1. Effects of Soil Fertility and Calcium Fertilizers on Seedling Height, Ground Diameter, and Dry Biomass
3.2. Effects of Soil Conditions and Calcium Fertilizers on Root Growth and Development
3.3. N, P, and Ca Accumulation Efficiencies
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Azevedo, L.B.; Zelm, R.V.; Hendriks, A.J.; Bobbink, R.; Mark, A.J.H. Global assessment of the effects of terrestrial acidification on plant species richness. Environ. Pollut. 2013, 174, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.X.; Pan, G.X.; Li, L.Q.; Hu, Z.L.; Wang, X.Z. Leaf N/P ratio and nutrient reuse between dominant species and stands: Predicting phosphorus deficiencies in karst ecosystems, southwestern China. Environ. Earth Sci. 2011, 64, 299–309. [Google Scholar] [CrossRef]
- Jiang, Z.C.; Lian, Y.Q.; Qin, X.Q. Rocky desertification in Southwest China: Impacts, causes, and restoration. Earth Sci. Rev. 2014, 132, 1–12. [Google Scholar] [CrossRef]
- Kudla, J.; Batistič, O.; Hashimoto, K. Calcium signals: The lead currency of plant information processing. Plant Cell 2010, 22, 541–563. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.H.U.; Tigabu, M.; Chen, H.; Farooq, T.H.; Ma, X.Q.; Wu, P.F. Calcium-mediated adaptive responses to low phosphorus stress in Chinese fir. Trees Struct. Funct. 2020, 34, 825–834. [Google Scholar] [CrossRef]
- Hepler, P.K. Calcium: A central regulator of plant growth and development. Plant Cell 2005, 17, 2142–2155. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, M.K.; Manzoor, M. Effect of soil-applied calcium carbide and plant derivatives on nitrification inhibition and plant growth promotion. Int. J. Environ. Sci. Technol. 2013, 10, 961–972. [Google Scholar] [CrossRef] [Green Version]
- Jörg, P.; Karl, E.R.; Ulrich, S.; Hans, P. Changes of soil chemistry, stand nutrition, and stand growth at two Scots pine (Pinus sylvestris L.) sites in Central Europe during 40 years after fertilization, liming, and lupine introduction. Eur. J. For. Res. 2008, 127, 43–61. [Google Scholar]
- Nilsson, S.I.; Andersson, S.; Valeur, I.; Persson, P.; Bergholm, J.; Wirén, A. Influence of dolomite lime on leaching and storage of C, N and S in a Spodosol under Norway spruce (Picea abies (L.) Karst.). For. Ecol. Manag. 2001, 146, 55–73. [Google Scholar] [CrossRef]
- Jammes, F.; Hu, H.C.; Villiers, F.; Bouten, R.; Kwak, J.M. Calcium permeable channels in plant cells. FEBS J. 2011, 278, 4262–4276. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, G.; Yu, K.X.; Li, P.; Xiao, L.; Liu, G. A new method to optimize root order classification based on the diameter interval of fine root. Sci. Rep. 2018, 8, 2960. [Google Scholar] [CrossRef] [Green Version]
- Souri, M.K.; Hatamian, M. Aminochelates in plant nutrition: A review. J. Plant Nutr. 2019, 42, 67–78. [Google Scholar] [CrossRef]
- Littke, K.M.; Zabowski, D. Calcium uptake, partitioning, and sinuous growth in Douglas-fir seedlings. For. Sci. 2007, 53, 692–700. [Google Scholar]
- Lundström, U.S.; Bain, D.C.; Taylor, A.F.S.; Van Hees, P.A.W.; Geibe, C.E.; Holmström, S.J.M.; Melkerud, P.A.; Finlay, R.; Jones, D.L.; Nyberg, L.; et al. Effects of Acidification and its Mitigation with Lime and Wood Ash on Forest Soil Processes in Southern Sweden. Water Air Soil Pollut. Focus 2003, 3, 167–188. [Google Scholar] [CrossRef]
- Hodge, A. The plastic plant: Root responses to heterogeneous supplies of nutrients. New Phytol. 2004, 162, 9–24. [Google Scholar] [CrossRef]
- Mommer, L.; van Ruijven, J.; Jansen, C.; van de Steeg, H.M.; de Kroon, H. Interactive effects of nutrient heterogeneity and competition: Implications for root foraging theory? Funct. Ecol. 2012, 26, 66–73. [Google Scholar] [CrossRef]
- Eric, D.R.; Philip, N.B. Regulation of plant root system architecture:implications for crop advancement. Curr. Opin. Biotechnol. 2015, 32, 93–98. [Google Scholar]
- Meinen, C.; Hertel, D.; Leuschner, C. Biomass and morphology of fine roots in temperate broad-leaved forests differing in tree species diversity: Is there evidence of below-ground overyielding? Oecologia 2009, 161, 99–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.P.; Zhang, M.L.; Smet, I.D.; Ding, Z.J. Designer crops: Optimalroot system architecture for nutrient acquisition. Trends Biotechnol. 2014, 32, 597–598. [Google Scholar] [CrossRef]
- Zhang, Z.; Jin, G.Q.; Zhou, Z.C. Seedling growth, root development and nutrient use efficiency of Cypress clones in response to calcium fertilizer. Dendrobiology 2020, 84, 39–48. [Google Scholar] [CrossRef]
- Nakahata, R. Pioneer root invasion and fibrous root development into disturbed soil space observed with a flatbed scanner method. Trees Struct. Funct. 2020, 34, 731–743. [Google Scholar] [CrossRef]
- Schlesinger, W.H. Community Structure, Dynamics and Nutrient Cycling in the Okefenokee Cypress Swamp. Ecol. Monograghs 1978, 48, 43–65. [Google Scholar] [CrossRef]
- Moorberg, C.J.; Vepraskas, M.J.; Niewoehner, C.P. Dynamics of P dissolution processes in the matrix and rhizospheres of bald cypress growing in saturated soil. Geoderma 2013, 202, 153–160. [Google Scholar] [CrossRef]
- Geibe, C.E.; Holmström, S.J.M.; van Hees, P.A.W.; Lundström, U.S. Impact of lime and ash applications on soil solution chemistry of an acidified podzolic soil. Water Air Soil Pollut. Focus 2002, 3, 77–96. [Google Scholar] [CrossRef]
- Pagani, A.; Mallarino, A.P. Soil pH and Crop Grain Yield as Affected by the Source and Rate of Lime. Soil Sci. Soc. Am. J. 2012, 76, 1877–1886. [Google Scholar] [CrossRef]
- Gu, J.C.; Xu, Y.; Dong, X.Y.; Wang, H.F.; Wang, Z.Q. Root diameter variations explained by anatomy and phylogeny of 50 tropical and temperate tree species. Tree Physiol. 2014, 34, 415–425. [Google Scholar] [CrossRef] [Green Version]
- Holdaway, R.J.; Richardson, S.J.; Dickie, I.A.; Peltzer, D.A.; Coomes, D.A. Speciesand community-level patterns in fine root traits along a 120,000-year soil chronosequence in temperate rain forest. J. Ecol. 2011, 99, 954–963. [Google Scholar] [CrossRef]
- He, Y.L.; Liu, A.; Tigabu, M.; Wu, P.; Ma, X.; Wang, C.; Oden, P.C. Physiological responses of needles of Pinus massoniana elite families to phosphorus stress in acid soil. J. For. Res. 2013, 24, 325–332. [Google Scholar] [CrossRef]
- Ohat, Y.; Yamamoto, K.; Deguchi, M. Chemical fractionation of calcium in the fresh leaf blade and influences of deficiency or over supply of calcium and age of leaf on the content of each calcium fraction. J. Sci. Soil Manure 1970, 41, 19–26. [Google Scholar]
- Christian, H.; Roland, B.; Axel, G.; Wendelin, W. Changes in soil, seepage water and needle chemistry between 1984 and 2004 after liming an N-saturated Norway spruce stand at the Höglwald, Germany. For. Ecol. Manag. 2006, 233, 11–20. [Google Scholar]
- Taylor, A.F.S.; Finlay, R.D. Effects of liming and ash application on below ground ectomycorrhizal community structure in two Norway spruce forests. Water Air Soil Pollut. Focus 2003, 3, 63–76. [Google Scholar] [CrossRef]
- Valeur, I.; Andersson, S.; Nilsson, I. Calcium content of liming material and its effect on sulphur release in a coniferous forest soil. Biogeochemistry 2000, 50, 1–20. [Google Scholar] [CrossRef]
- Borchert, R. Calcium acetate induces calcium uptake and formation of calcium-oxalate crystals in isolated leaflets of Gleditsia tracanthos L. Planta 1986, 168, 571–578. [Google Scholar] [CrossRef]
- Franceschi, V.R. Calcium oxalate formation is a rapid and reversible process in Lemna minor L. Protoplasma 1989, 148, 130–137. [Google Scholar] [CrossRef]
- Chan, C.W.M.; Wohlbach, D.J.; Rodesch, M.J.; Sussman, M.R. Transcriptional changes in response to growth of Arabidopsis in high external calcium. FEBS. Lett. 2008, 582, 967–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.L.; Helmisaari, H.S.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B.; et al. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef]
- Pourranjbari, S.S.; Souri, M.K. Root growth characteristics of K hatouni melon seedlings as affected by potassium nutrition. Hortorum Cultus 2018, 17, 191–198. [Google Scholar]
- Eissenstat, D.M.; Yanni, R.D. Root Lifespan, Efficiency and Turnover; CRC Press: New York, NY, USA, 2002; pp. 221–238. [Google Scholar]
- Liao, Y.C.; McCormack, M.L.; Fan, H.B.; Wang, H.M.; Wu, J.P.; Tu, J.; Liu, W.F.; Guo, D.L. Relation of fine root distribution to soil C in a Cunninghamia lanceolata plantation in subtropical China. Plant Soil 2014, 381, 225–234. [Google Scholar] [CrossRef]
- Mou, P.; Robert, H.J.; Tan, Z.Q.; Bao, Z.; Chen, H.M. Morphological and physiological plasticity of plant roots when nutrients are both spatially and temporally heterogeneous. Plant Soil 2013, 364, 373–384. [Google Scholar] [CrossRef]
- Eissenstat, D.M.; Wells, C.E.; Yanai, R.D.; Whitbeck, J.L. Building roots in a changing environment: Implications for root longevity. New Phytol. 2000, 147, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Espeleta, J.F.; Donovan, L.A. Fine root demography and morphology in response to soil resources availability among xeric and mesic sandhill tree species. Funct. Ecol. 2002, 16, 113–121. [Google Scholar] [CrossRef]
- Brassard, B.W.; Chen, H.Y.H.; Bergeron, Y.; Paré, D. Differences in fine root productivity between mixed and single-species stands. Funct. Ecol. 2011, 25, 238–246. [Google Scholar] [CrossRef]
- Guo, D.L.; Li, H.; Mitchell, R.J.; Han, W.X.; Hendricks, J.J.; Fahey, T.J.; Hendrick, R.L. Heterogeneity by root branch order: Exploring the discrepancy in root longevity and turnover estimates between minirhizotron and C isotope methods. New Phytol. 2008, 177, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Band, L.R.; Bennett, M.J. Mapping the site of action of the Green Revolution hormone gibberellin. Proc. Natl. Acad. Sci. USA 2013, 110, 4443–4444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barger, S.A.; Walker, J.M.; Vasey, E.H. Mechanisms for the movement of plant nutrients from the soil and fertilizer to the plant root. J. Agric. Food Chem. 1963, 11, 204–207. [Google Scholar]
- Kinzel, H. Calcium in the vacuoles and cell walls of plant tissue. Flora 1989, 182, 99–125. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient Elements | Texture | Soil Type | Total N (g·kg−1) | Total P (g·kg−1) | Hydrolytic N (mg·kg−1) | Available K (mg·kg−1) | Available P (mg·kg−1) | Organic Matter (g·kg−1) | Exchange Ca (mg·kg−1) | Exchange Mg (mg·kg−1) | pH Value |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Average content | light loam | red acid soil | 0.75 ± 0.09 | 0.32 ± 0.05 | 53.5 ± 4.70 | 18.50 ± 1.12 | 0.99 ± 0.14 | 15.8 ± 1.89 | 128 ± 12.5 | 9.24 ± 0.85 | 4.65 ± 0.21 |
| NPK Fertilizer Treatment | Trait | CaO Treatment | F Value | ||||
|---|---|---|---|---|---|---|---|
| 0 g·kg−1 | 3 g·kg−1 | 6 g·kg−1 | CaO | Clones | Clones × CaO | ||
| NPK fertilizer 0 g·kg−1 soil | Seedling height (cm) | 29.67 ± 2.56 | 44.49 ± 3.87 | 33.17 ± 3.28 | 8.26 ** | 5.57 ** | 3.54 * |
| ground diameter (cm) | 4.57 ± 0.35 | 5.74 ± 0.51 | 4.89 ± 0.38 | 15.65 ** | 7.21 ** | 4.07 ** | |
| Root dry matter (g) | 2.84 ± 0.21 | 3.96 ± 0.36 | 3.43 ± 0.34 | 7.58 ** | 5.53 ** | 2.43 * | |
| Stem dry matter (g) | 2.09 ± 0.18 | 3.16 ± 0.34 | 2.56 ± 0.31 | 3.59 * | 5.32 ** | 1.46 | |
| Leaf dry matter (g) | 4.19 ± 0.36 | 6.29 ± 0.61 | 5.33 ± 0.42 | 6.41 ** | 4.34 ** | 2.89 * | |
| NPK fertilizer 3 g·kg−1 soil | Seedling height (cm) | 48.67 ± 5.17 | 49.70 ± 4.88 | 51.36 ± 5.04 | 0.59 | 8.65 ** | 2.45 * |
| Ground diameter (mm) | 6.02 ± 0.58 | 6.38 ± 0.54 | 6.16 ± 0.57 | 1.57 | 12.04 ** | 4.03 ** | |
| Root dry matter (g) | 3.91 ± 0.34 | 5.48 ± 0.52 | 4.56 ± 0.34 | 2.74 * | 6.81 ** | 2.22 * | |
| Stem dry matter (g) | 3.76 ± 0.29 | 4.88 ± 0.39 | 4.38 ± 0.42 | 1.20 | 10.04 ** | 2.51 * | |
| Leaf dry matter (g) | 7.74 ± 0.59 | 8.75 ± 0.81 | 8.59 ± 0.78 | 0.97 | 8.12 ** | 2.81 * | |
| NPK Fertilizer Treatment | Trait | Diameter Class (mm) | CaO Treatment | F Value | ||||
|---|---|---|---|---|---|---|---|---|
| 0 g·kg−1 | 3 g·kg−1 | 6 g·kg−1 | CaO | Clones | Clones × CaO | |||
| NPK fertilizer 0 g·kg−1 soil | Root length (cm) | 0–0.5 (D1) | 1093.24 ± 89.21 | 1451.61 ± 100.36 | 1302.28 ± 98.65 | 6.11 ** | 1.46 | 1.27 |
| 0.5–1.0 (D2) | 458.41 ± 31.41 | 569.99 ± 50.24 | 566.18 ± 60.04 | 4.16 ** | 2.45 * | 1.54 | ||
| 1.0–1.5 (D3) | 92.93 ± 10.02 | 125.71 ± 10.32 | 114.12 ± 8.97 | 5.88 ** | 3.90 ** | 1.87 | ||
| 1.5–2.0 (D4) | 21.70 ± 2.14 | 25.53 ± 2.07 | 23.21 ± 1.97 | 7.07 ** | 3.18* | 1.37 | ||
| >2.0 (D5) | 15.64 ± 1.85 | 18.52 ± 1.58 | 17.15 ± 2.04 | 1.21 | 2.03 | 1.17 | ||
| Root Surface area (cm2) | 0–0.5 (D1) | 117.38 ± 17.21 | 138.49 ± 15.47 | 129.98 ± 16.04 | 5.32 ** | 3.12 * | 1.77 | |
| 0.5–1.0 (D2) | 87.63 ± 8.36 | 128.69 ± 13.04 | 112.37 ± 12.11 | 4.97 ** | 2.43 * | 1.54 | ||
| 1.0–1.5 (D3) | 38.02 ± 3.21 | 45.69 ± 4.23 | 42.97 ± 3.04 | 6.45 ** | 2.76 * | 1.52 | ||
| 1.5–2.0 (D4) | 7.96 ± 0.47 | 13.74 ± 0.98 | 10.91 ± 0.75 | 7.03 ** | 0.98 | 1.04 | ||
| >2.0 (D5) | 15.91 ± 1.02 | 20.46 ± 1.68 | 16.46 ± 1.47 | 1.63 | 0.34 | 2.68 * | ||
| Root volume (cm3) | 0–0.5 (D1) | 0.90 ± 0.08 | 1.25 ± 0.11 | 1.14 ± 0.09 | 4.93 ** | 1.76 | 7.75 ** | |
| 0.5–1.0 (D2) | 1.64 ± 0.12 | 2.22 ± 0.09 | 1.98 ± 0.10 | 5.08 ** | 4.43 ** | 0.88 | ||
| 1.0–1.5 (D3) | 0.98 ± 0.06 | 1.35 ± 0.07 | 1.22 ± 0.06 | 7.12 ** | 6.75 ** | 1.43 | ||
| 1.5–2.0 (D4) | 0.40 ± 0.02 | 0.61 ± 0.03 | 0.54 ± 0.03 | 7.54 ** | 0.81 | 1.60 | ||
| >2.0 (D5) | 1.52 ± 0.11 | 2.31 ± 0.16 | 1.99 ± 0.16 | 2.32 | 0.61 | 2.20 | ||
| NPK fertilizer 3 g·kg−1 soil | Root length (cm) | 0–0.5 (D1) | 1485.46 ± 105.24 | 1749.83 ± 135.36 | 1395.10 ± 101.24 | 3.27 * | 5.67 ** | 1.36 |
| 0.5–1.0 (D2) | 981.03 ± 85.31 | 1147.28 ± 91.04 | 863.67 ± 74.36 | 7.31 ** | 7.22 ** | 5.67 ** | ||
| 1.0–1.5 (D3) | 205.25 ± 18.36 | 232.34 ± 17.65 | 218.88 ± 19.32 | 6.45 ** | 5.16 ** | 5.49 ** | ||
| 1.5–2.0 (D4) | 40.48 ± 3.65 | 45.28 ± 4.02 | 37.95 ± 3.21 | 2.09 | 9.26 ** | 1.24 | ||
| >2.0 (D5) | 22.71 ± 2.11 | 28.28 ± 2.05 | 25.02 ± 2.24 | 1.53 | 4.89 ** | 0.98 | ||
| Root Surface area (cm2) | 0–0.5 (D1) | 136.62 ± 9.57 | 169.39 ± 14.03 | 149.19 ± 12.32 | 5.52 ** | 7.14 ** | 1.65 | |
| 0.5–1.0 (D2) | 197.27 ± 15.36 | 225.99 ± 19.65 | 191.71 ± 17.65 | 9.23 ** | 5.05 ** | 1.88 | ||
| 1.0–1.5 (D3) | 79.78 ± 6.32 | 85.60 ± 7.05 | 81.28 ± 6.95 | 6.77 ** | 6.28 ** | 3.08 * | ||
| 1.5–2.0 (D4) | 20.10 ± 1.83 | 24.09 ± 2.04 | 20.61 ± 1.76 | 1.68 | 8.16 ** | 4.73 ** | ||
| >2.0 (D5) | 23.73 ± 1.51 | 29.72 ± 2.38 | 25.37 ± 2.01 | 0.98 | 4.78 ** | 1.69 | ||
| Root volume (cm3) | 0–0.5 (D1) | 1.18 ± 0.11 | 1.60 ± 0.13 | 1.27 ± 0.10 | 9.32 ** | 8.01 ** | 4.79 ** | |
| 0.5–1.0 (D2) | 3.14 ± 0.39 | 4.22 ± 0.32 | 3.54 ± 0.23 | 5.11 ** | 5.44 ** | 2.04 | ||
| 1.0–1.5 (D3) | 2.17 ± 0.19 | 2.51 ± 0.21 | 2.40 ± 0.21 | 7.02 ** | 8.26 ** | 3.14 * | ||
| 1.5–2.0 (D4) | 0.86 ± 0.07 | 1.03 ± 0.11 | 0.90 ± 0.06 | 1.43 | 10.31 ** | 6.70 ** | ||
| >2.0 (D5) | 2.75 ± 0.21 | 3.46 ± 0.28 | 2.83 ± 0.20 | 1.58 | 12.21 ** | 1.59 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Jin, G.; Chen, T.; Zhou, Z. Effects of CaO on the Clonal Growth and Root Adaptability of Cypress in Acidic Soils. Forests 2021, 12, 922. https://doi.org/10.3390/f12070922
Zhang Z, Jin G, Chen T, Zhou Z. Effects of CaO on the Clonal Growth and Root Adaptability of Cypress in Acidic Soils. Forests. 2021; 12(7):922. https://doi.org/10.3390/f12070922
Chicago/Turabian StyleZhang, Zhen, Guoqing Jin, Tan Chen, and Zhichun Zhou. 2021. "Effects of CaO on the Clonal Growth and Root Adaptability of Cypress in Acidic Soils" Forests 12, no. 7: 922. https://doi.org/10.3390/f12070922
APA StyleZhang, Z., Jin, G., Chen, T., & Zhou, Z. (2021). Effects of CaO on the Clonal Growth and Root Adaptability of Cypress in Acidic Soils. Forests, 12(7), 922. https://doi.org/10.3390/f12070922