
Bioclimatic Suitability of Actual and Potential Cultivation Areas for Jacaranda mimosifolia in Chinese Cities


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Sources
2.2. Variable Selection
2.3. Bioclimatic Indices
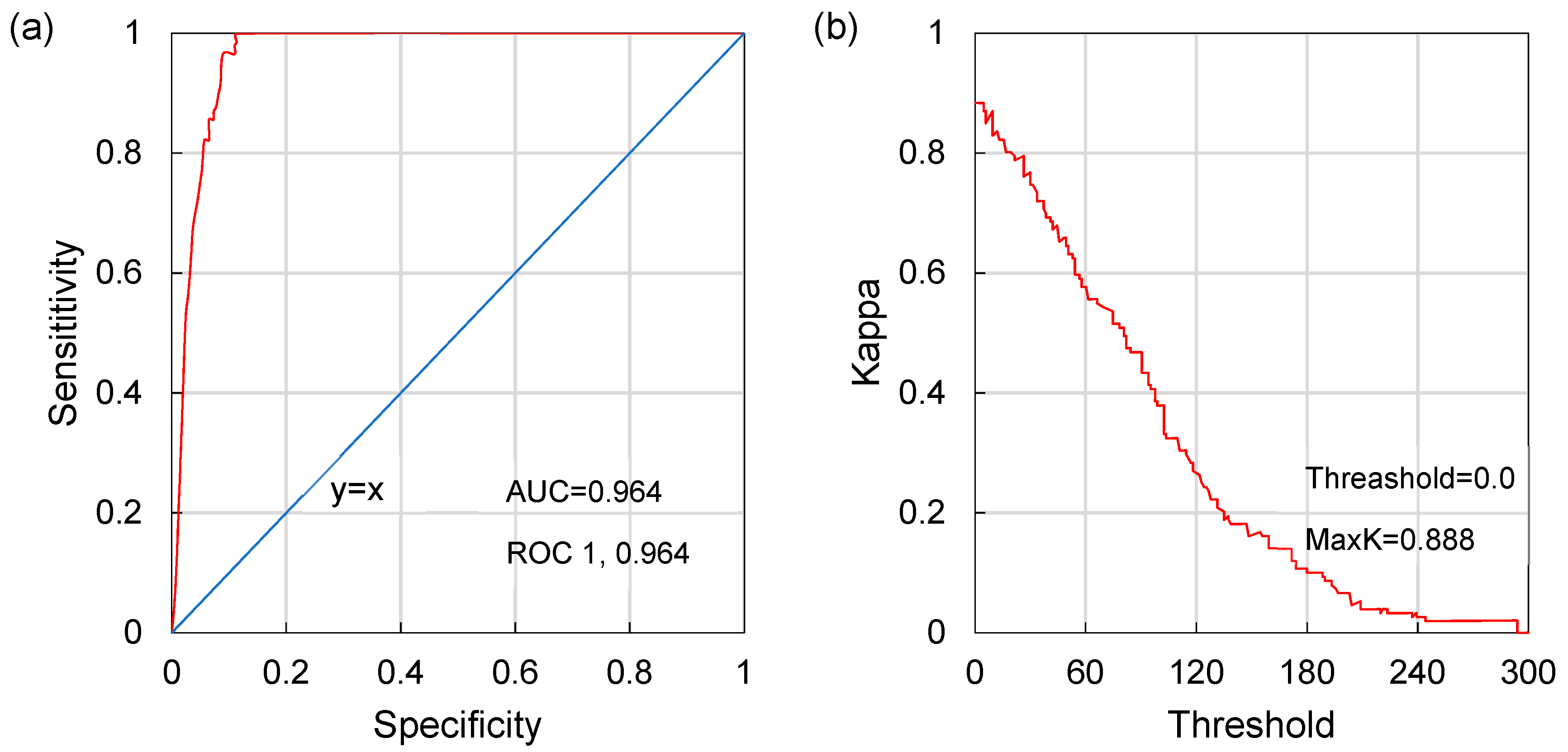
2.4. Modeling Procedures and Validation
3. Results
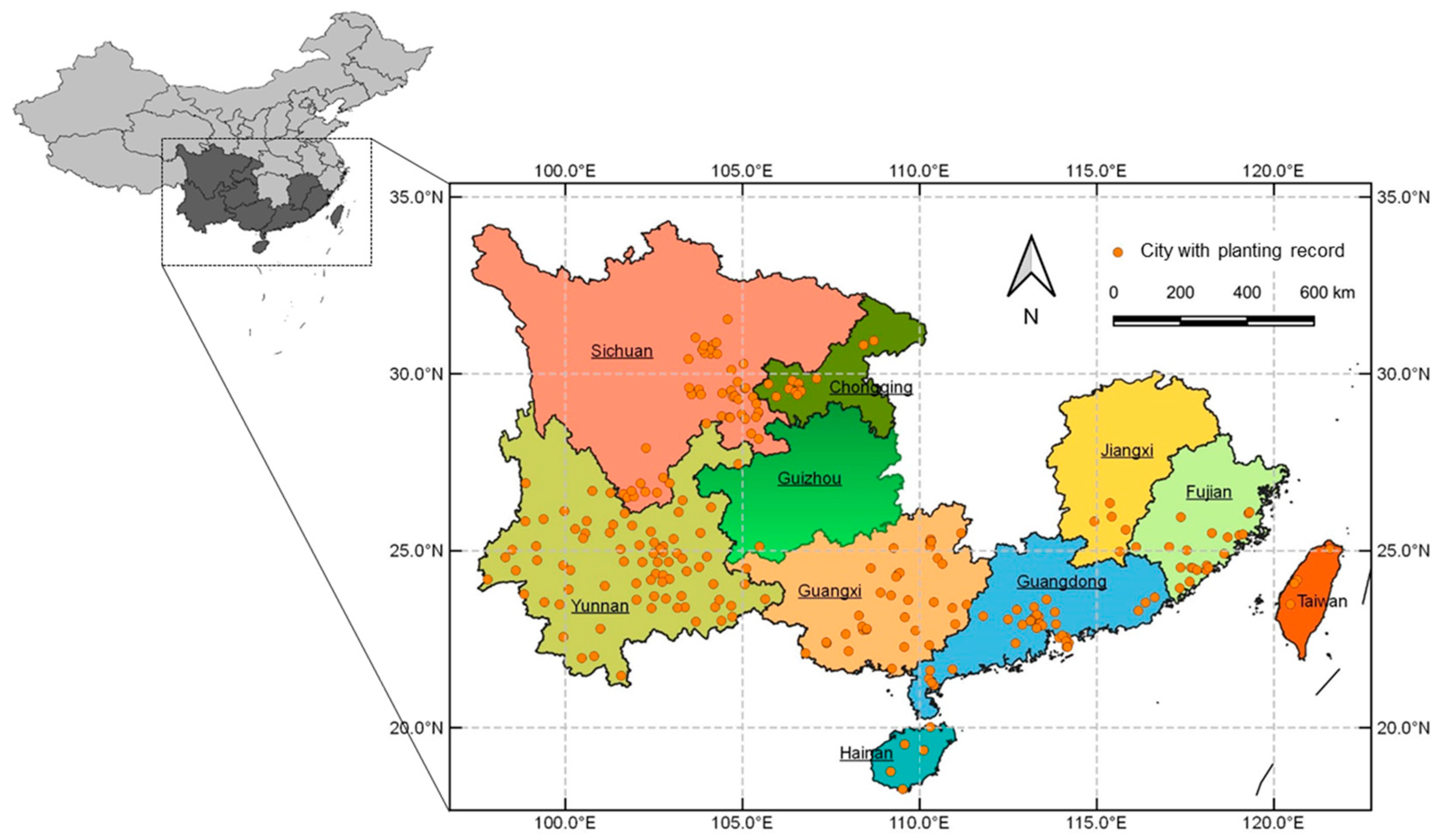
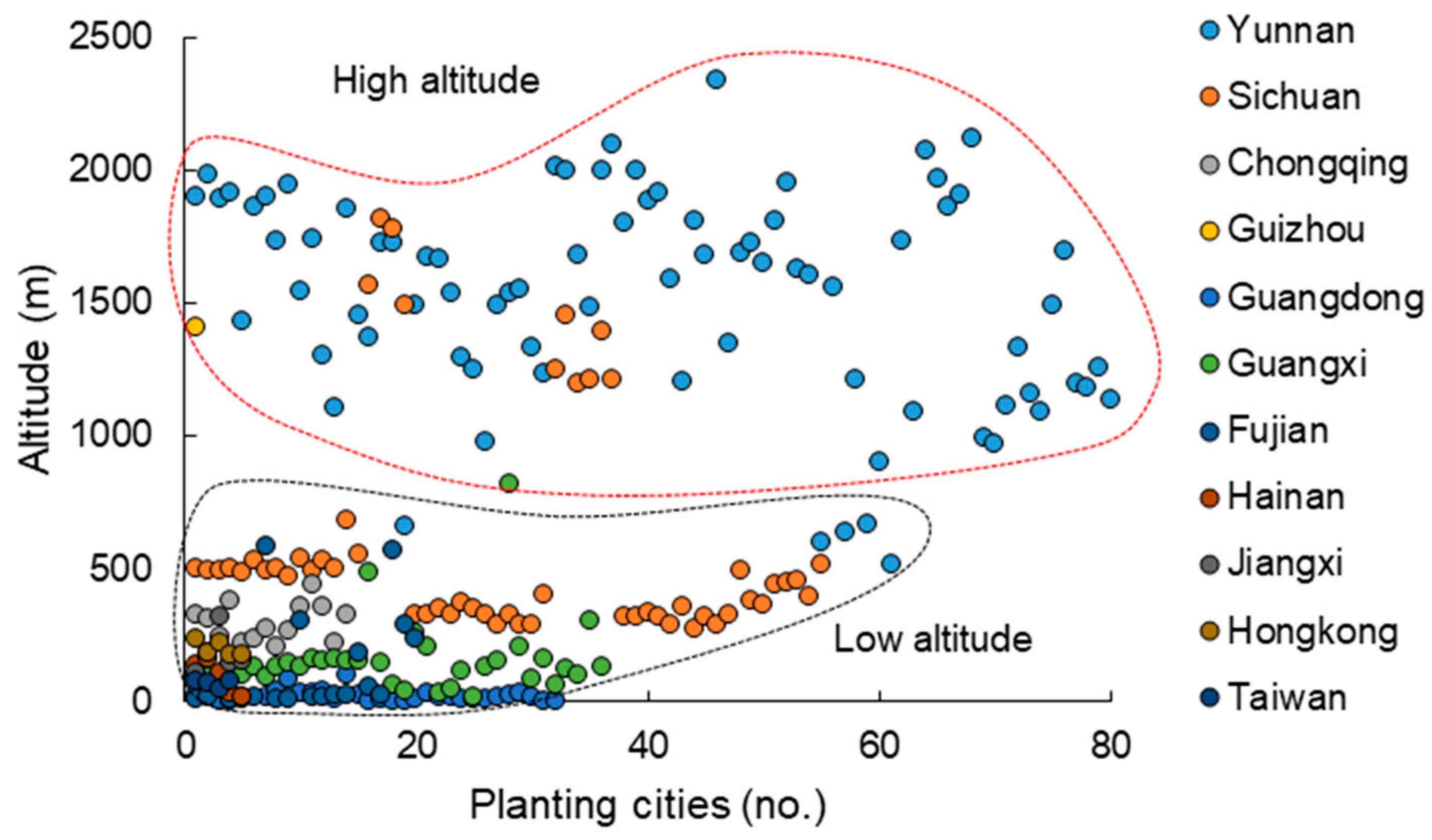
3.1. Current Cultivated Areas
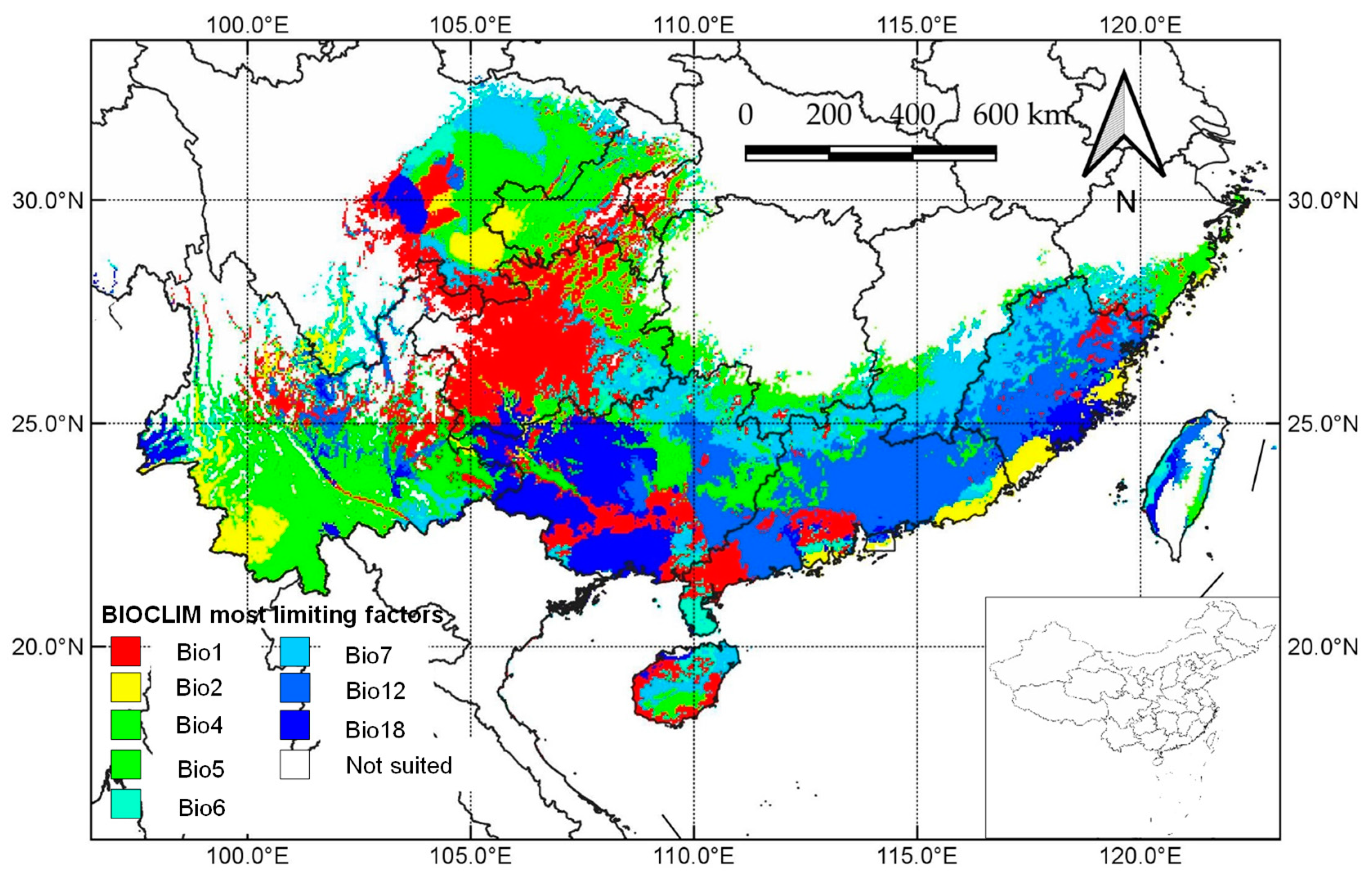
3.2. Climatic Factors Controlling Cultivated Areas
3.3. Main Climatic Parameters and Suitable Ranges
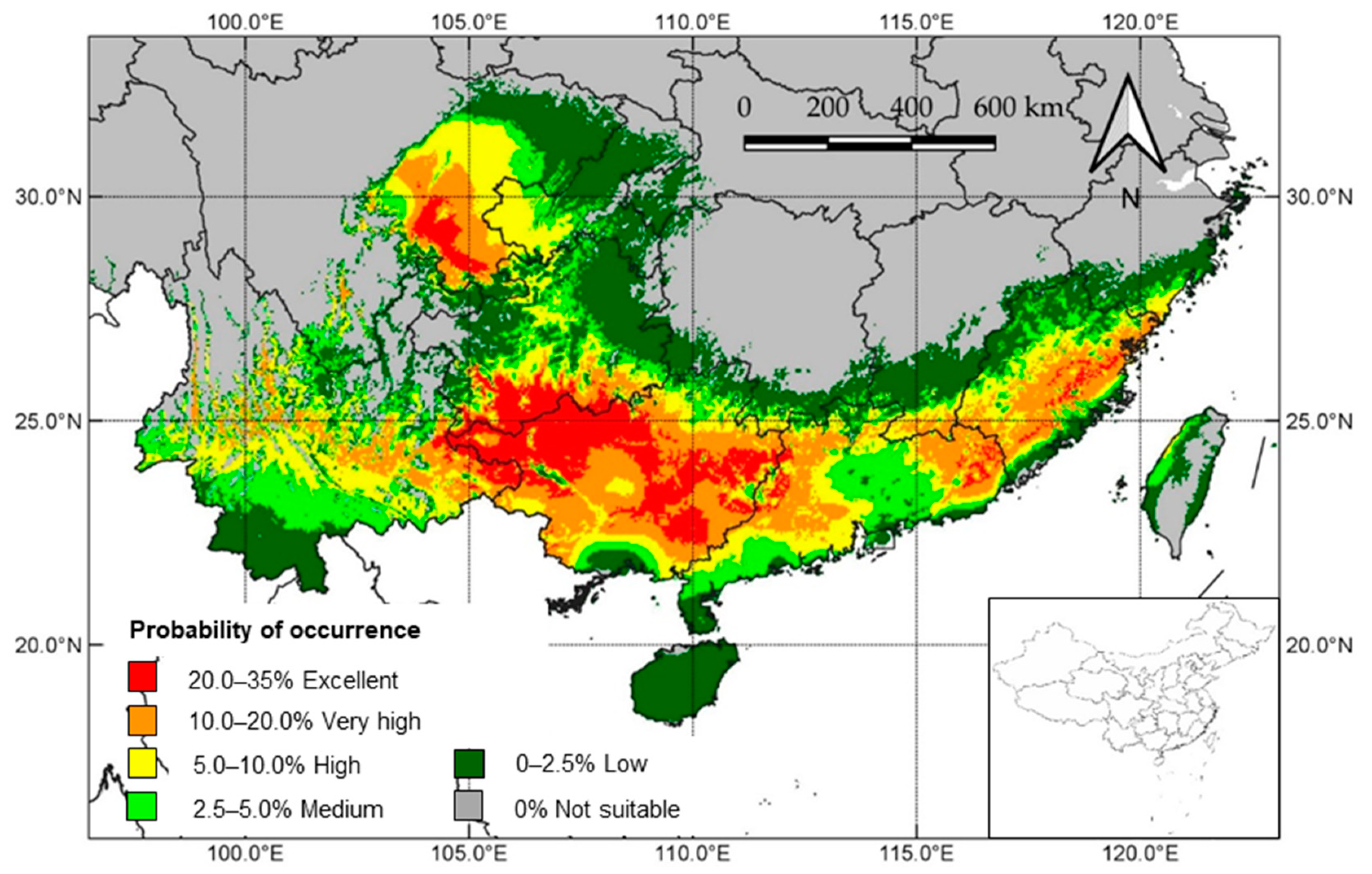
3.4. Modeling Suitable Cultivation Areas
4. Discussion
4.1. Geographical Distribution Pattern of J. mimosifolia in China
4.2. Key Climatic Parameters Limiting the Northward Spread of J. mimosifolia
4.3. Suitable Cultivation Areas of J. mimosifolia in China
4.4. Ecological Invasion Alert and Science-Based Plant Introduction
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Çoban, S.; Yener, Ş.D.; Bayraktar, S. Woody plant composition and diversity of urban green spaces in Istanbul, Turkey. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2021, 155, 83–91. [Google Scholar] [CrossRef]
- Jim, C.Y.; Liu, H.T. Species diversity of three major urban forest types in Guangzhou City, China. For. Ecol. Manag. 2001, 146, 99–114. [Google Scholar] [CrossRef]
- Loram, A.; Thompson, K.; Warren, P.H.; Gaston, K.J. Urban domestic gardens (XII): The richness and composition of the flora in five UK cities. J. Veg. Sci. 2008, 19, 321–330. [Google Scholar] [CrossRef]
- Sjöman, H.; Östberg, J.; Bühler, O. Diversity and distribution of the urban tree population in ten major Nordic cities. Urban For. Urban Green. 2012, 11, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Zisenis, M. Alien plant species: A real fear for urban ecosystems in Europe? Urban Ecosyst. 2015, 18, 355–370. [Google Scholar] [CrossRef]
- Donaldson, J.E.; Hui, C.; Richardson, D.M.; Robertson, M.P.; Webber, B.L.; Wilson, J.R.U. Invasion trajectory of alien trees: The role of introduction pathway and planting history. Glob. Chang. Biol. 2014, 20, 1527–1537. [Google Scholar] [CrossRef] [PubMed]
- Maurel, N.; Hanspach, J.; Kühn, I.; Pyšek, P.; van Kleunen, M. Introduction bias affects relationships between the characteristics of ornamental alien plants and their naturalization success. Glob. Ecol. Biogeogr. 2016, 25, 1500–1509. [Google Scholar] [CrossRef] [Green Version]
- Moro, M.F.; Westerkamp, C. The alien street trees of Fortaleza (NE Brazil): Qualitative observations and the inventory of two districts. Ciência Florest. 2011, 21, 789–798. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.P. Tree diversity in urban parks of Dublin, Ireland. Fresenius Environ. Bull. 2018, 27, 8695–8708. [Google Scholar]
- Kowarik, I. Urban ornamentals escaped from cultivation. In Crop Ferality and Volunteerism; CRC Press: Boca Raton, FL, USA, 2005; pp. 97–121. [Google Scholar] [CrossRef]
- Richardson, D.M.; Rejmánek, M. Trees and shrubs as invasive alien species—A global review. Divers. Distrib. 2011, 17, 788–809. [Google Scholar] [CrossRef]
- Čeplová, N.; Lososová, Z.; Kalusová, V. Urban ornamental trees: A source of current invaders; a case study from a European City. Urban Ecosyst. 2017, 20, 1135–1140. [Google Scholar] [CrossRef]
- Zhu, H.F.; Zhang, C.Q.; Gong, X. A summary on plant introduction and acclimatization research. Guihaia 2003, 23, 52–60. [Google Scholar] [CrossRef]
- Lenzner, B.; Magallón, S.; Dawson, W.; Kreft, H.; König, C.; Pergl, J.; Pyšek, P.; Weigelt, P.; van Kleunen, M.; Winter, M.; et al. Role of diversification rates and evolutionary history as a driver of plant naturalization success. New Phytol. 2021, 229, 2998–3008. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.N.; Mbari, N.J.; Wang, S.W.; Liu, B.; Mwangi, B.N.; Rasoarahona, J.R.E.; Xin, H.P.; Zhou, Y.D.; Wang, Q.F. Modeling impacts of climate change on the potential distribution of six endemic baobab species in Madagascar. Plant Divers. 2021, 43, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Van de Ven, C.M.; Weiss, S.B.; Ernst, W.G. Plant species distributions under present conditions and forecasted for warmer climates in an arid mountain range. Earth Interact. 2007, 11, 1–33. [Google Scholar] [CrossRef]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Using MaxEnt modeling to predict the potential distribution of the endemic plant Rosa arabica Crép. in Egypt. Ecol. Inform. 2019, 50, 68–75. [Google Scholar] [CrossRef]
- Deb, C.R.; Jamir, N.; Kikon, Z.P. Distribution prediction model of a rare orchid species (Vanda bicolor Griff.) using small sample size. Am. J. Plant Sci. 2017, 8, 1388–1398. [Google Scholar] [CrossRef]
- Araújo, M.B.; Luoto, M. The importance of biotic interactions for modelling species distributions under climate change. Glob. Ecol. Biogeogr. 2007, 16, 743–753. [Google Scholar] [CrossRef]
- Zhang, D.S.; Liu, M. Introduction dynamics of landscape trees in Shanghai during the recent 55 years based on climatic envelope model. Chin. Landsc. Archit. 2018, 34, 118–123. [Google Scholar] [CrossRef]
- Wei, B.; Wang, R.L.; Hou, K.; Wang, X.Y.; Wu, W. Predicting the current and future cultivation regions of Carthamus tinctorius L. using MaxEnt model under climate change in China. Glob. Ecol. Conserv. 2018, 16, e00477. [Google Scholar] [CrossRef]
- Kabaš, E.; Batanjski, V.; Glasnović, P.; Vicić, D.; Tanasković, A.; Kuzmanovic, N.; Lakusic, D.; Šinžar-Sekulić, J. Towards detecting bioclimatic niche—Species distribution modelling in four maple species (Acer spp.). Acta Bot. Croat. 2014, 73, 401–417. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.; Lechowicz, M.J. Climatic limits for the present distribution of beech (Fagus L.) species in the world. J. Biogeogr. 2006, 33, 1804–1819. [Google Scholar] [CrossRef]
- Ni, J. An introduction to bioclimatic factors in global change research. Quat. Sci. 2017, 37, 431–441. [Google Scholar] [CrossRef]
- Kadmon, R.; Farber, O.; Danin, A. A systematic analysis of factors affecting the performance of climatic envelope models. Ecol. Appl. 2003, 13, 853–867. [Google Scholar] [CrossRef]
- Villordon, A. GIS on the cheap: DIVA-GIS and other free data visualization tools for research. HortScience 2006, 41, 497–520. [Google Scholar] [CrossRef] [Green Version]
- Yi, Y.J.; Cheng, X.; Yang, Z.F.; Zhang, S.H. Maxent modeling for predicting the potential distribution of endangered medicinal plant (H. riparia Lour) in Yunnan, China. Ecol. Eng. 2016, 92, 260–269. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Silva, L.D.; Costa, H.; de Azevedo, E.B.; Medeiros, V.; Alves, M.; Elias, R.B.; Silva, L. Modelling native and invasive woody species: A comparison of ENFA and MaxEnt applied to the Azorean forest. In Proceedings of the International Conference on Dynamics, Games and Science, Porto, Portugal, 17–21 February 2014; pp. 415–444. [Google Scholar]
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A.; NCEAS Predicting Species Distributions Working Group. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Segurado, P.; Araújo, M.B. An evaluation of methods for modelling species distributions. J. Biogeogr. 2004, 31, 1555–1568. [Google Scholar] [CrossRef]
- Ahmed, N.; Atzberger, C.; Zewdie, W. Species Distribution Modelling performance and its implication for Sentinel-2-based prediction of invasive Prosopis juliflora in lower Awash River basin, Ethiopia. Ecol. Process. 2021, 10, 18. [Google Scholar] [CrossRef]
- Booth, T.H.; Nix, H.A.; Busby, J.R.; Hutchinson, M.F. Bioclim: The first species distribution modelling package, its early applications and relevance to most current MaxEnt studies. Divers. Distrib. 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Beaumont, L.J.; Hughes, L.; Poulsen, M. Predicting species distributions: Use of climatic parameters in BIOCLIM and its impact on predictions of species’ current and future distributions. Ecol. Model. 2005, 186, 251–270. [Google Scholar] [CrossRef]
- Booth, T.H. Why understanding the pioneering and continuing contributions of BIOCLIM to species distribution modelling is important. Austral Ecol. 2018, 43, 852–860. [Google Scholar] [CrossRef] [Green Version]
- Felizardo, M.P.; Merlo, G.R.; Maia, G.D. Modeling drying kinetics of Jacaranda mimosifolia seeds with variable effective diffusivity via diffusion model. Biosys. Eng. 2021, 205, 234–245. [Google Scholar] [CrossRef]
- Nick, P. Jacaranda mimosifolia (Jacaranda). Available online: https://www.cabi.org/isc/datasheet/29212#tosummaryOfInvasiveness (accessed on 8 May 2021).
- Jia, P. Landscape tree Species: Jacaranda mimosifolia. China Flower Hortic. 2008, 4, 24–25. [Google Scholar]
- Liu, X.F.; Li, X.M.; Zhang, P.J.; Zhang, G.W. The preliminary Introduction of Jacaranda mimosifolia provenances to Zhanjiang. Eucalypt Sci. Technol. 2016, 33. [Google Scholar] [CrossRef]
- Oliveira, J.R.d.; Costa, C.A.S.d.; Bezerra, A.M.E.; Abud, H.F.; de Lucena, E.M.P. Characterization of seeds, seedlings and initial growth of Jacaranda mimosifolia D. Don.(Bignoniaceae). Rev. Árvore 2018, 42, e420403. [Google Scholar] [CrossRef]
- Alves, G.; Peruchi, A.; Agostini, K. Pollination in urban area: The Jacaranda mimosifolia D. Don (Bignoniaceae) case study. Bioikos 2010, 24, 31–41. [Google Scholar]
- Miyajima, I.; Mata, D.; Kobayashi, N.; Facciuto, G.; Soto, S.; Hagiwara, J.C.; Serpa, J.C.; Escandon, A. Practical Method of Propagating Jacaranda mimosifolia by Cuttings. J. Jpn. Soc. Hortic. Sci. 2004, 73, 137–139. [Google Scholar] [CrossRef]
- Zhou, J.; Zeng, X.Y.; He, W.; Li, J.Y.; Zhang, W.; Li, X.Q. Mechanism of chilling injury in Jacaranda acutifolia Humb.et Bonpl. under chilling stress. Southwest China J. Agric. Sci. 2016, 29, 74–80. [Google Scholar] [CrossRef]
- Naz, R.; Roberts, T.H.; Bano, A.; Nosheen, A.; Yasmin, H.; Hassan, M.N.; Keyani, R.; Ullah, S.; Khan, W.; Anwar, Z. GC-MS analysis, antimicrobial, antioxidant, antilipoxygenase and cytotoxic activities of Jacaranda mimosifolia methanol leaf extracts and fractions. PLoS ONE 2020, 15, e0236319. [Google Scholar] [CrossRef]
- Sidjui, L.S.; Zeuko’o, E.M.; Toghueo, R.M.K.; Noté, O.P.; Mahiou-Leddet, V.; Herbette, G.; Fekam, F.B.; Ollivier, E.; Folefoc, G.N. Secondary metabolites from Jacaranda mimosifolia and Kigelia africana (Bignoniaceae) and their anticandidal activity. Rec. Nat. Prod. 2014, 8, 307. [Google Scholar]
- Aguirre-Becerra, H.; Pineda-Nieto, S.A.; García-Trejo, J.F.; Guevara-González, R.G.; Feregrino-Pérez, A.A.; Álvarez-Mayorga, B.L.; Pastrana, D.M.R. Jacaranda flower (Jacaranda mimosifolia) as an alternative for antioxidant and antimicrobial use. Heliyon 2020, 6, e05802. [Google Scholar] [CrossRef] [PubMed]
- Brignole, D.; Drava, G.; Minganti, V.; Giordani, P.; Samson, R.; Vieira, J.; Pinho, P.; Branquinho, C. Chemical and magnetic analyses on tree bark as an effective tool for biomonitoring: A case study in Lisbon (Portugal). Chemosphere 2018, 195, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Olowoyo, J.O.; Van Heerden, E.; Fischer, J. Investigating Jacaranda mimosifolia tree as biomonitor of atmospheric trace metals. Environ. Monit. Assess. 2010, 164, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J.Climatol. A J. R. Meteorol. Soc. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Yue, Y.J.; Zhang, P.Y.; Shang, Y.R. The potential global distribution and dynamics of wheat under multiple climate change scenarios. Sci. Total Environ. 2019, 688, 1308–1318. [Google Scholar] [CrossRef] [PubMed]
- Sillero, N. What does ecological modelling model? A proposed classification of ecological niche models based on their underlying methods. Ecol. Model. 2011, 222, 1343–1346. [Google Scholar] [CrossRef]
- Cao Pinna, L.; Axmanová, I.; Chytrý, M.; Malavasi, M.; Acosta, A.T.R.; Giulio, S.; Attorre, F.; Bergmeier, E.; Biurrun, I.; Campos, J.A.; et al. The biogeography of alien plant invasions in the Mediterranean Basin. J. Veg. Sci. 2021, 32, e12980. [Google Scholar] [CrossRef]
- Janžekovič, F.; Novak, T. PCA—A Powerful Method for Analyze Ecological Niches. In Principal Component Analysis—Multidisciplinary Applications; Sanguansat, P., Ed.; InTech: Rijeka, Croatia, 2012; pp. 127–142. [Google Scholar] [CrossRef] [Green Version]
- Federici, F.; Pignatti, S. The warmth index of Kira for the interpretation of vegetation belts in Italy and SW. Australia two regions with Mediterranean type bioclimates. Vegetatio 1991, 93, 91–99. [Google Scholar] [CrossRef]
- Nagai, S.; Saitoh, T.M.; Miura, T. Peak autumn leaf colouring along latitudinal and elevational gradients in Japan evaluated with online phenological data. Int. J. Biometeorol. 2020, 64, 1743–1754. [Google Scholar] [CrossRef]
- Yim, Y.J.; Kira, T. Distribution of forest vegetation and climate in the Korean peninsula.: I. Distribution of some indices of thermal climate. Jpn. J. Ecol. 1975, 25, 77–88. [Google Scholar] [CrossRef]
- Krestov, P.V.; Nakamura, Y. Climatic controls of forest vegetation distribution in Northeast Asia. Ber. Reinhold-Tüxen-Ges. 2007, 19, 131–145. [Google Scholar]
- Chiu, C.A.; Chiou, C.R.; Lin, J.R.; Lin, P.H.; Lin, C.T. Coldness index does not indicate the upper limit of evergreen broad-leaved forest on a subtropical island. J. For. Res. 2014, 19, 115–124. [Google Scholar] [CrossRef]
- Szelepcsényi, Z.; Breuer, H.; Kis, A.; Pongrácz, R.; Sümegi, P. Assessment of projected climate change in the Carpathian Region using the Holdridge life zone system. Theor. Appl. Climatol. 2018, 131, 593–610. [Google Scholar] [CrossRef]
- Li, G.Q.; Wen, Z.M.; Guo, K.; Du, S. Simulating the effect of climate change on vegetation zone distribution on the Loess Plateau, Northwest China. Forests 2015, 6, 2092–2108. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.S.; Quan, C. Late Cenozoic climates of low-latitude East Asia: A paleobotanical example from the Baise Basin of Guangxi, southern China. Palaeoworld 2017, 26, 572–580. [Google Scholar] [CrossRef]
- Quan, C.; Han, S.; Utescher, T.; Zhang, C.; Liu, Y.S. Validation of temperature–precipitation based aridity index: Paleoclimatic implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 386, 86–95. [Google Scholar] [CrossRef]
- Si, S.B.; Cao, F.X.; Peng, J.Q.; Xu, R.X. Determination of northern distribution boundary and relationship with climate parameters for Rhodomyrtus tomentosa in China. J. Cent. South Univ. For. Technol. 2012, 32, 162–165. [Google Scholar] [CrossRef]
- Ni, J. Development of Kira’ s indices and its application to vegetation-climate interaction study of China. J. Appl. Ecol. 1997, 8, 161–170. [Google Scholar]
- Hong, B.G.; Li, S.Z. The preliminary study of the correlations between the distribution of main everygreen broad-leaf tree species in Jiangsu and climates. Acta Ecol. Sin. 1981, 1, 105–111. [Google Scholar]
- Stewart, S.B.; Elith, J.; Fedrigo, M.; Kasel, S.; Roxburgh, S.H.; Bennett, L.T.; Chick, M.; Fairman, T.; Leonard, S.; Kohout, M.; et al. Climate extreme variables generated using monthly time-series data improve predicted distributions of plant species. Ecography 2021, 44, 626–639. [Google Scholar] [CrossRef]
- Li, Z.R.; Zhu, Z.F.; Wu, Y. Scale dependency of pseudo-absences selection and uncertainty in climate scenarios matter when assessing potential distribution of a rare poppy plant Meconopsis punicea Maxim. under a warming climate. Glob. Ecol. Conserv. 2020, 24, e01353. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, C.X.; Sun, C.Z.; Zhang, D.F.; Guo, J.; Wu, M.L.; ZHang, Q. Predicting the global areas for potential distribution of Gastrodia elata based on ecological niche models. Chin. J. Plant Ecol. 2017, 41, 770–778. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, X.; Yang, Q.; Li, D.; Long, W.; Luo, W. Factors affecting the distribution pattern of wild plants with extremely small populations in Hainan Island, China. PLoS ONE 2014, 9, e97751. [Google Scholar] [CrossRef] [Green Version]
- Kosanic, A.; Anderson, K.; Harrison, S.; Turkington, T.; Bennie, J. Changes in the geographical distribution of plant species and climatic variables on the West Cornwall peninsula (South West UK). PLoS ONE 2018, 13, e0191021. [Google Scholar] [CrossRef] [Green Version]
- Huang, E.H.; Chen, Y.X.; Fang, M.; Zheng, Y.; Yu, S.X. Environmental drivers of plant distributions at global and regional scales. Glob. Ecol. Biogeogr. 2021, 30, 697–709. [Google Scholar] [CrossRef]
- Nievola, C.C.; Carvalho, C.P.; Carvalho, V.; Rodrigues, E. Rapid responses of plants to temperature changes. Temperature (Austin) 2017, 4, 371–405. [Google Scholar] [CrossRef]
- He, F.; He, B. Cultivated distribution and site classification for camellia oleifera. Sci. Silvae Sin. 2002, 38, 64–72. [Google Scholar] [CrossRef]
- Takahashi, Y.; Li, X.H.; Qiu, L.J.; Tsukamoto, C.; Wang, K.J. Identification of saponin composition and their geographical distribution in Chinese cultivated soybean (Glycine max). Euphytica 2017, 213, 175. [Google Scholar] [CrossRef]
- Popoola, J.; Obembe, O. Local knowledge, use pattern and geographical distribution of Moringa oleifera Lam. (Moringaceae) in Nigeria. J. Ethnopharmacol. 2013, 150, 682–691. [Google Scholar] [CrossRef]
- Wilson, B.; Dolotbakov, A.; Burgess, B.J.; Clubbe, C.; Lazkov, G.; Shalpykov, K.; Ganybaeva, M.; Sultangaziev, O.; Brockington, S.F. Central Asian wild tulip conservation requires a regional approach, especially in the face of climate change. Biodivers. Conserv. 2021, 30, 1705–1730. [Google Scholar] [CrossRef]
- Edgin, B. Status and Distribution of Illinois Populations of Stenanthium gramineum (Ker-Gawl.) Morong, Grass-Leaved Lily (Liliaceae): An Endangered Plant in Illinois. Castanea 2004, 69, 216–225. [Google Scholar] [CrossRef]
- Dai, S.P.; Li, H.L.; Liu, H.Q.; Liu, E.P. The spatio-temporal change characteristics of agriculture climate resources in southern china under the background of global warming. Chin. J. Agric. Resour. Reg. Plan. 2014, 35, 52–60. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Mao, W.S.; Pang, B. Characteristics of climate change in Chengdu Plain. J. Chengdu Univ. Inf. Technol. 2020, 35, 179–187. [Google Scholar] [CrossRef]
- Pu, J.C.; Huang, Z.Y.; Gao, M. Relationship between climatic characteristics and planting suitability of main cash crops in Yunnan. J. Meteorol. Res. Appl. 2021, 42, 53–57. [Google Scholar] [CrossRef]
- Ighbareyeh, J.M.H.; Carmona, E.C.; Ortiz, A.C.; Suliemieh, A.A.-R.A.; Ighbareyeh, M.M.H.; Daraweesh, A.A.-Q.M. Analysis of physical factors of climate and bioclimate and their effects on almonds production to increase the economy in Hebron area of Palestine. Arab. J. Geosci. 2018, 11, 683. [Google Scholar] [CrossRef]
- Prentice, I.C.; Cramer, W.; Harrison, S.P.; Leemans, R.; Monserud, R.A.; Solomon, A.M. Special paper: A global biome model based on plant physiology and dominance, soil properties and climate. J. Biogeogr. 1992, 19, 117–134. [Google Scholar] [CrossRef]
- Woodward, F.I.; Williams, B.G. Climate and plant distribution at global and local scales. Vegetatio 1987, 69, 189–197. [Google Scholar] [CrossRef]
- Zhang, C.J.; Liao, Y.M.; Song, Y.L. The progress of dry–wet climate divisional research in China. Earth Sci. 2020, 9, 8–15. [Google Scholar] [CrossRef]
- Zhang, X.W.; Wang, J.R.; Wang, M.H.; Yang, Y.; Zhao, C.M. Dominant climatic factors influencing the geographical distribution pattern of Picea in China. Sci. Silvae Sin. 2020, 56, 1–11. [Google Scholar] [CrossRef]
- Syfert, M.M.; Smith, M.J.; Coomes, D.A. The effects of sampling bias and model complexity on the predictive performance of MaxEnt species distribution models. PLoS ONE 2013, 8, e55158. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Zhu, H.; Yi, X.G.; Li, Y.F.; Duan, Y.F.; Wang, X.R.; Zhang, L.B. Limiting climatic factors in shaping the distribution pattern and niche differentiation of Prunus dielsiana in subtropical China. J. For. Res. 2020, 1–11. [Google Scholar] [CrossRef]
- Brenzel, K.N. Sunset Western Garden Book; Sunset Publishing Group: Southern Progress’s Menlo Park, CA, USA, 2007. [Google Scholar]
- Liu, X.F.; Li, X.M.; Zhang, P.J.; Xu, Y.X.; Fang, L.; Zhang, G.W. Research progress of Jacaranda mimosifolia and the development prospects. Eucalypt Sci. Technol. 2015, 32, 51–55. [Google Scholar] [CrossRef]
- Miyajima, I.; Takemura, C.; Kobayashi, N.; Soto, M.S.; Facciuto, G. Flower bud initiation and Development of Jacaranda mimosifolia (Bignoniaceae) in Japan. In Proceedings of the VII International Symposium on New Floricultural Crops 1000, Buenos Aires, Argentina, 22–25 November 2011; pp. 71–76. [Google Scholar]
- Jenerette, G.D.; Clarke, L.W.; Avolio, M.L.; Pataki, D.E.; Gillespie, T.W.; Pincetl, S.; Nowak, D.J.; Hutyra, L.R.; McHale, M.; McFadden, J.P.; et al. Climate tolerances and trait choices shape continental patterns of urban tree biodiversity. Glob. Ecol. Biogeogr. 2016, 25, 1367–1376. [Google Scholar] [CrossRef]
- Widrlechner, M.P.; Daly, C.; Keller, M.; Kaplan, K. Horticultural applications of a newly revised USDA plant Hardiness Zone Map. Am. Soc. Hortic. Sci. 2012, 22, 6–19. [Google Scholar] [CrossRef] [Green Version]
- Widrlechner, M.P. Interactive Gardening and Plant Hardiness Zone Map for China. Available online: https://www.plantmaps.com/interactive-china-plant-hardiness-zone-map-celsius.php (accessed on 6 May 2021).
- Richardson, D.M.; Van Wilgen, B.W. Invasive alien plants in South Africa: How well do we understand the ecological impacts? S. Afr. J. Sci. 2004, 100, 45–52. [Google Scholar]
- Zimmermann, H.G.; Moran, V.C.; Hoffmann, J. Biological control in the management of invasive alien plants in South Africa, and the role of the Working for Water Programme. S. Afr. J. Sci. 2004, 100, 34–40. [Google Scholar]
- Witt, A.; Luke, Q. Guide to the Naturalized and Invasive Plants of Eastern Africa; CABI: Nairobi, Kenya, 2017. [Google Scholar]
- Velasco-Jiménez, M.J.; Alcázar, P.; Cariñanos, P.; Galán, C. Allergenicity of the urban green areas in the city of Córdoba (Spain). Urban For. Urban Green. 2020, 49, 126600. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioclimatic Variable | Abbreviation | Unit | Data Source |
|---|---|---|---|
| Annual mean temperature | Bio1 | °C | CMDSC |
| Mean diurnal range | Bio2 | °C | WorldClim |
| Temperature seasonality a | Bio4 | - | WorldClim |
| Max. temperature of warmest month | Bio5 | °C | CMDSC |
| Min. temperature of coldest month | Bio6 | °C | CMDSC |
| Annual temperature range | Bio7 | °C | WorldClim |
| Annual precipitation | Bio12 | mm | CMDSC |
| Precipitation of warmest quarter | Bio18 | mm | WorldClim |
| Bioclimatic Variable | PC 1 | PC 2 | PC 3 | PC 4 | PC 5 | PC 6 | PC 7 |
|---|---|---|---|---|---|---|---|
| Bio1 Annual mean temperature | 0.291 | 0.112 | −0.236 | 0.122 | 0.163 | −0.036 | 0.020 |
| Bio2 Mean diurnal range a | −0.198 | 0.241 | −0.067 | −0.248 | 0.577 | 0.062 | −0.074 |
| Bio3 Isothermality b | −0.145 | 0.375 | −0.114 | −0.215 | 0.283 | 0.089 | −0.050 |
| Bio4 Temperature seasonality c | 0.051 | −0.445 | 0.148 | 0.126 | 0.063 | −0.092 | 0.039 |
| Bio5 Max. temperature of warmest month | 0.254 | −0.214 | −0.149 | 0.178 | 0.346 | −0.124 | −0.050 |
| Bio6 Min. temperature of coldest month | 0.288 | 0.129 | −0.218 | 0.153 | −0.171 | −0.019 | 0.041 |
| Bio7 Annual temperature range d | −0.098 | −0.373 | 0.118 | −0.009 | 0.557 | −0.102 | −0.101 |
| Bio8 Mean temperature of wettest quarter | 0.266 | −0.079 | −0.114 | 0.326 | 0.148 | 0.599 | 0.062 |
| Bio9 Mean temperature of driest quarter | 0.246 | 0.237 | −0.253 | −0.042 | 0.044 | −0.295 | 0.064 |
| Bio10 Mean temperature of warmest quarter | 0.288 | −0.169 | −0.131 | 0.192 | 0.145 | −0.110 | 0.033 |
| Bio11 Mean temperature of coldest quarter | 0.217 | 0.297 | −0.269 | 0.033 | 0.081 | −0.020 | 0.005 |
| Bio12 Annual precipitation | 0.283 | 0.106 | 0.287 | −0.078 | 0.018 | −0.089 | −0.254 |
| Bio13 Precipitation of wettest month | 0.195 | 0.180 | 0.428 | 0.102 | 0.086 | −0.309 | 0.216 |
| Bio14 Precipitation of driest month | 0.255 | −0.105 | 0.097 | −0.412 | 0.016 | 0.361 | 0.257 |
| Bio15 Precipitation seasonality e | −0.231 | 0.185 | 0.153 | 0.329 | 0.145 | 0.058 | 0.761 |
| Bio16 Precipitation of wettest quarter | 0.198 | 0.211 | 0.414 | 0.090 | 0.116 | −0.144 | −0.053 |
| Bio17 Precipitation of driest quarter | 0.264 | −0.095 | 0.101 | −0.404 | 0.025 | 0.257 | 0.202 |
| Bio18 Precipitation of warmest quarter | 0.146 | 0.238 | 0.418 | 0.181 | 0.006 | 0.331 | −0.328 |
| Bio19 Precipitation of coldest quarter | 0.261 | −0.103 | 0.031 | −0.404 | −0.044 | −0.247 | 0.245 |
| Eigenvalue | 8.951 | 4.472 | 2.400 | 1.727 | 0.808 | 0.242 | 0.178 |
| Variance % | 47.108 | 23.539 | 12.630 | 9.091 | 4.250 | 1.274 | 0.934 |
| Cumulative % | 47.108 | 70.647 | 83.277 | 92.368 | 96.618 | 97.892 | 98.826 |
| Bioclimatic Variable | Mean | Standard Deviation | Max | Min | Variation Coefficient % | Optimum Range |
|---|---|---|---|---|---|---|
| Bio1 Annual mean temperature | 19.37 | 2.53 | 25.76 | 12.38 | 13.05 | 16.39–22.34 |
| Bio3 Isothermality | 36.15 | 9.07 | 55.05 | 23.35 | 25.10 | 25.47–46.83 |
| Bio4 Temperature seasonality | 572.52 | 121.79 | 803.27 | 279.18 | 21.27 | 429.17–715.87 |
| Bio6 Min. temperature of coldest month | 6.35 | 3.40 | 17.40 | −0.60 | 53.54 | - |
| Bio7 Annual temperature range | 24.11 | 2.77 | 30.50 | 14.50 | 11.49 | 20.85–27.38 |
| Bio10 Mean temperature of warmest quarter | 25.76 | 2.80 | 29.07 | 18.83 | 10.86 | 22.46–29.05 |
| Bio11 Mean temperature of coldest quarter | 11.83 | 3.12 | 21.88 | 3.83 | 26.34 | 8.16–15.50 |
| Bio12 Annual precipitation | 1295.02 | 344.04 | 2742.00 | 677.00 | 26.57 | 890.08–1699.96 |
| WI Kira’s warmth index | 120.76 | 28.05 | 193.19 | 54.71 | 23.23 | 87.74–153.78 |
| CI Kira’s coldness index | 0.73 | 1.73 | 14.60 | 0.00 | 236.05 | - |
| ABT Holdridge’s annual biotemperature | 20.00 | 2.41 | 26.10 | 13.34 | 12.05 | 17.17–22.84 |
| AI Aridity index | 24.59 | 5.82 | 50.07 | 12.44 | 23.68 | 17.74–31.45 |
| BK Biological aridity | 10.60 | 2.24 | 20.51 | 5.14 | 21.15 | 7.96–13.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, C.; Zhang, G.; Jim, C.; Liu, X.; Zhang, P.; Qiu, J.; Liu, D. Bioclimatic Suitability of Actual and Potential Cultivation Areas for Jacaranda mimosifolia in Chinese Cities. Forests 2021, 12, 951. https://doi.org/10.3390/f12070951
Xie C, Zhang G, Jim C, Liu X, Zhang P, Qiu J, Liu D. Bioclimatic Suitability of Actual and Potential Cultivation Areas for Jacaranda mimosifolia in Chinese Cities. Forests. 2021; 12(7):951. https://doi.org/10.3390/f12070951
Chicago/Turabian StyleXie, Chunping, Guowu Zhang, Chiyung Jim, Xuefeng Liu, Peijian Zhang, Jianhuang Qiu, and Dawei Liu. 2021. "Bioclimatic Suitability of Actual and Potential Cultivation Areas for Jacaranda mimosifolia in Chinese Cities" Forests 12, no. 7: 951. https://doi.org/10.3390/f12070951
APA StyleXie, C., Zhang, G., Jim, C., Liu, X., Zhang, P., Qiu, J., & Liu, D. (2021). Bioclimatic Suitability of Actual and Potential Cultivation Areas for Jacaranda mimosifolia in Chinese Cities. Forests, 12(7), 951. https://doi.org/10.3390/f12070951