
Comparing Rooting Ability and Physiological Changes of Two Eucommia ulmoides Improved Varieties

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Materials
2.2. Experimental Design
2.3. Growth Index Statistics
2.4. Determination of Physiological Index
2.5. Data Analysis
3. Results
3.1. The Rooting of Softwood Cuttings of E. ulmoides
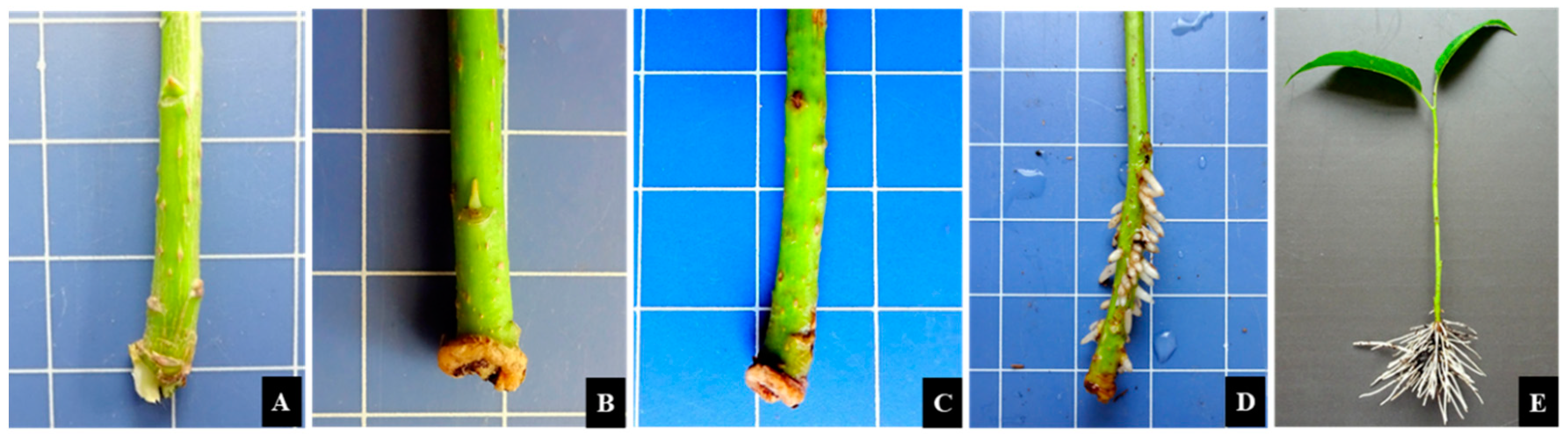
3.1.1. Morphological changes
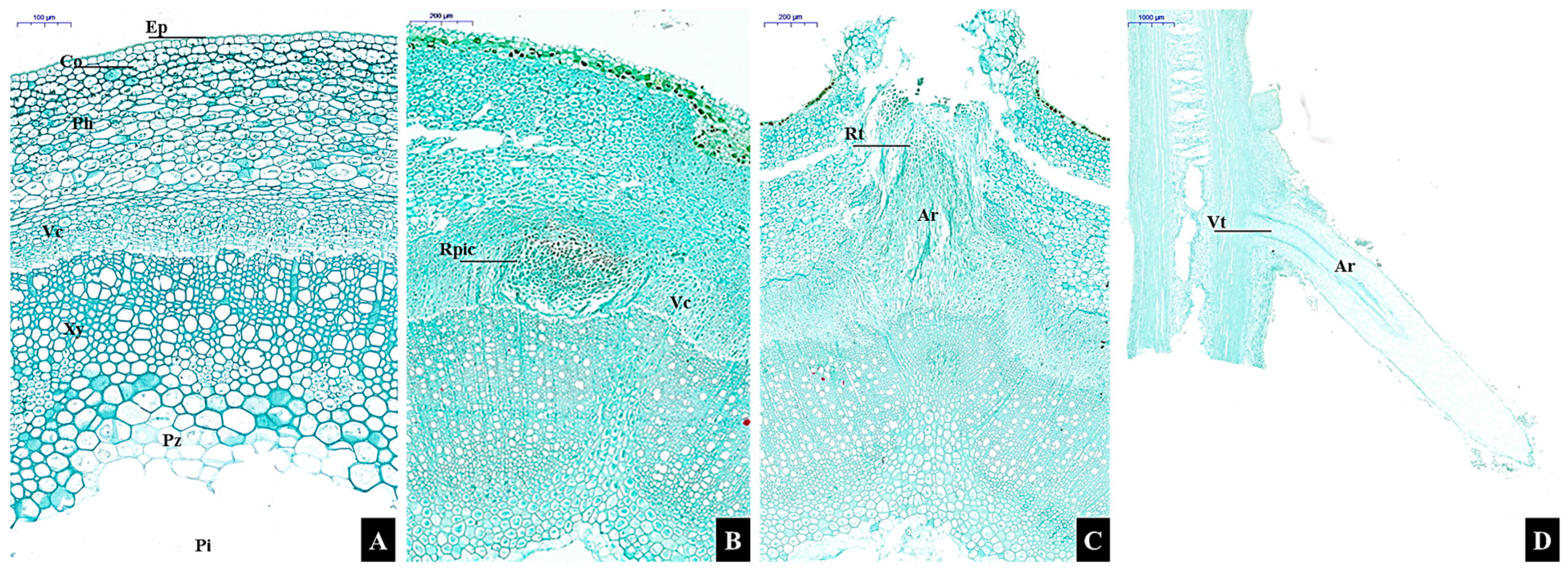
3.1.2. Microscopic Morphological Changes
3.1.3. Anatomical structure changes
3.2. Morphological Comparison of E. ulmoides Cuttings
3.3. The Content and Ratio of Endogenous Hormones
3.3.1. Changes in Endogenous Hormone Content
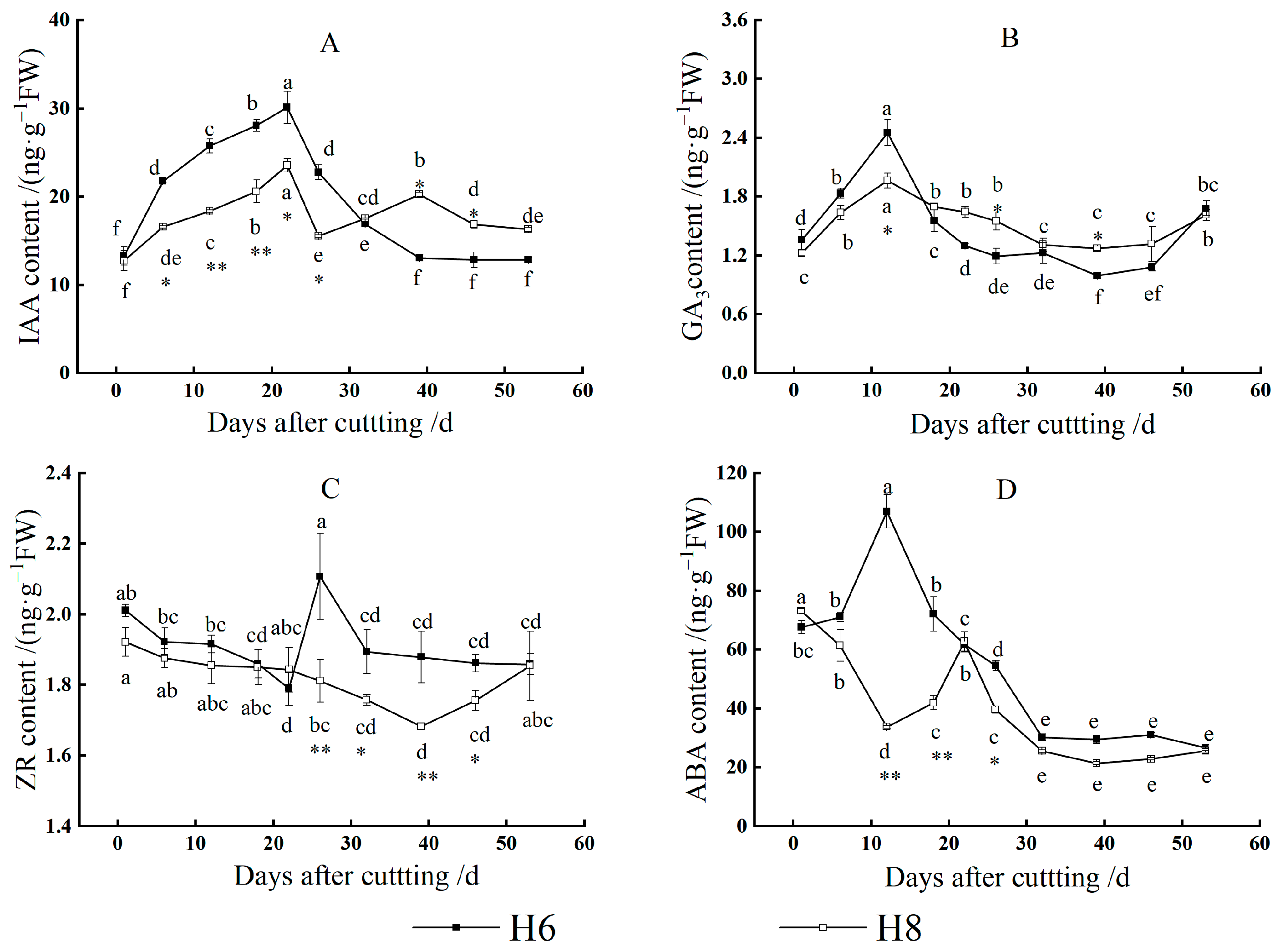
Changes in IAA Content
Changes in GA3 Content
Changes in ZR Content
Changes in ABA Content
3.3.2. Changes of Endogenous Hormones Ratio
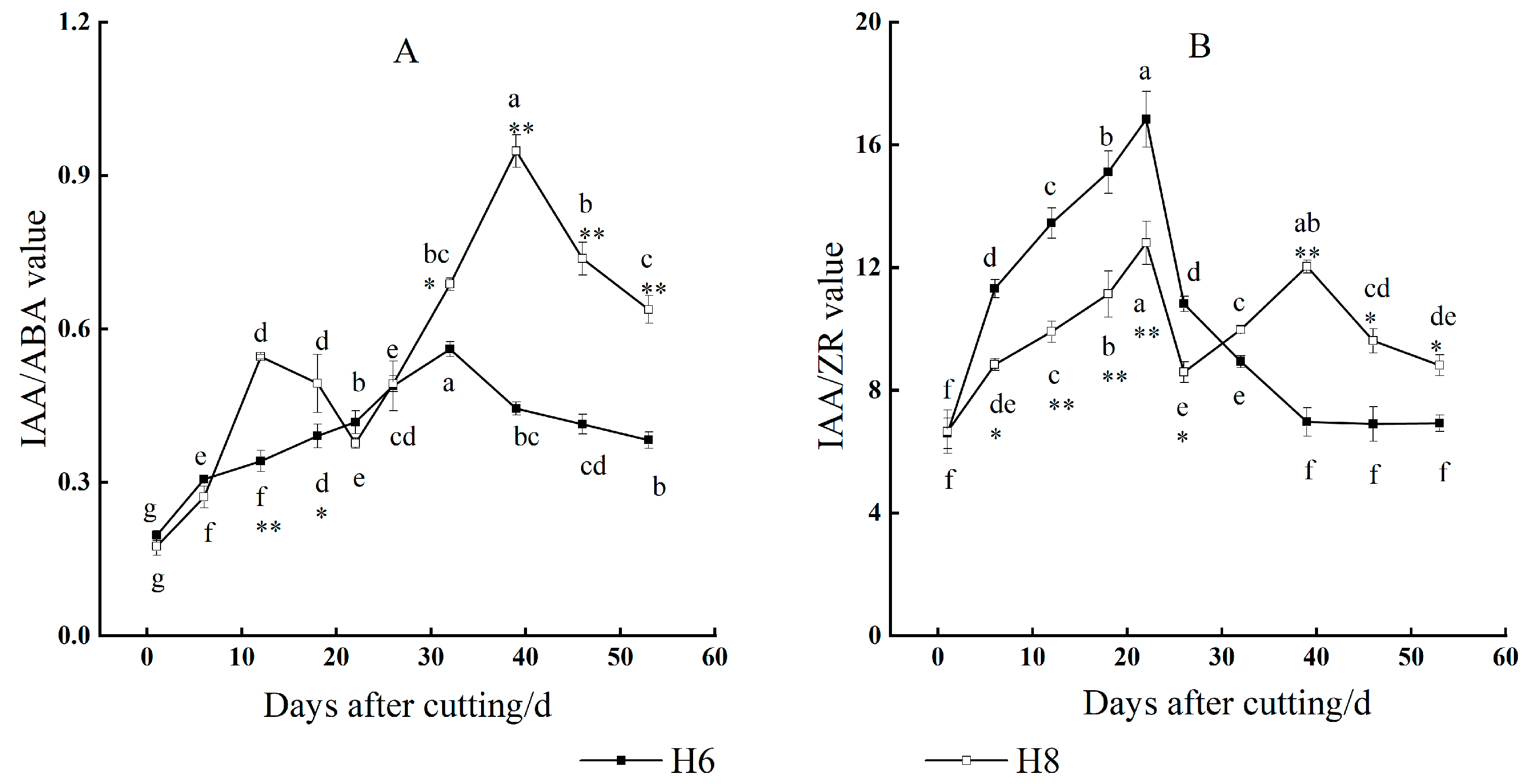
Changes in IAA/ABA
Changes in IAA/ZR Value
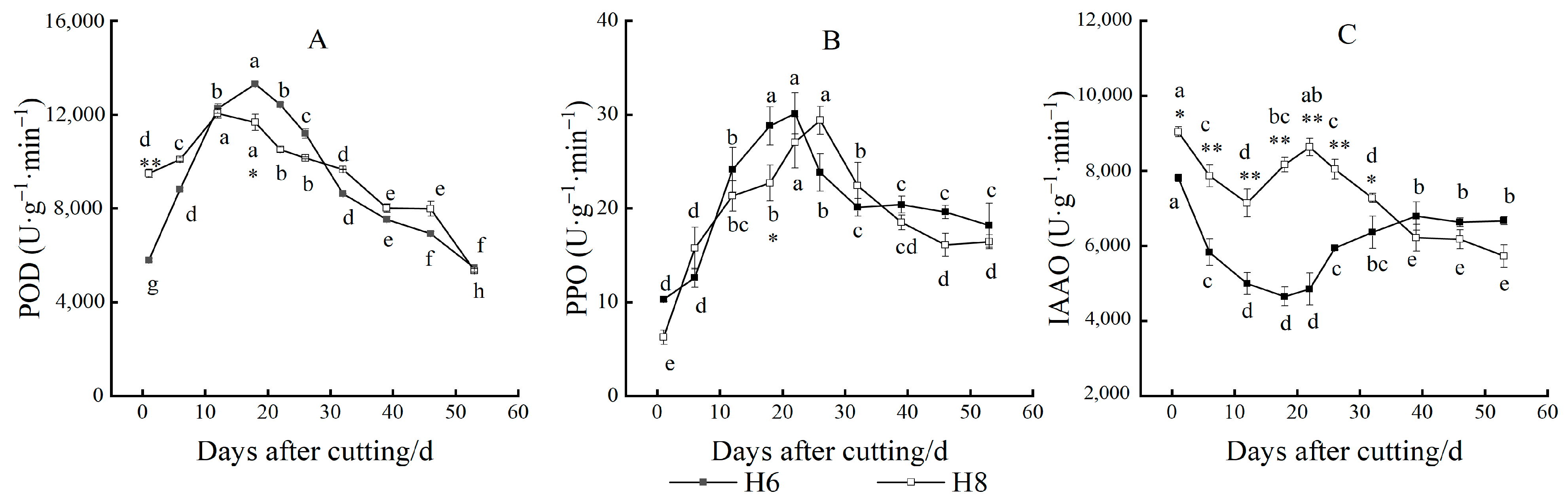
3.4. Changes in Oxidase Activity
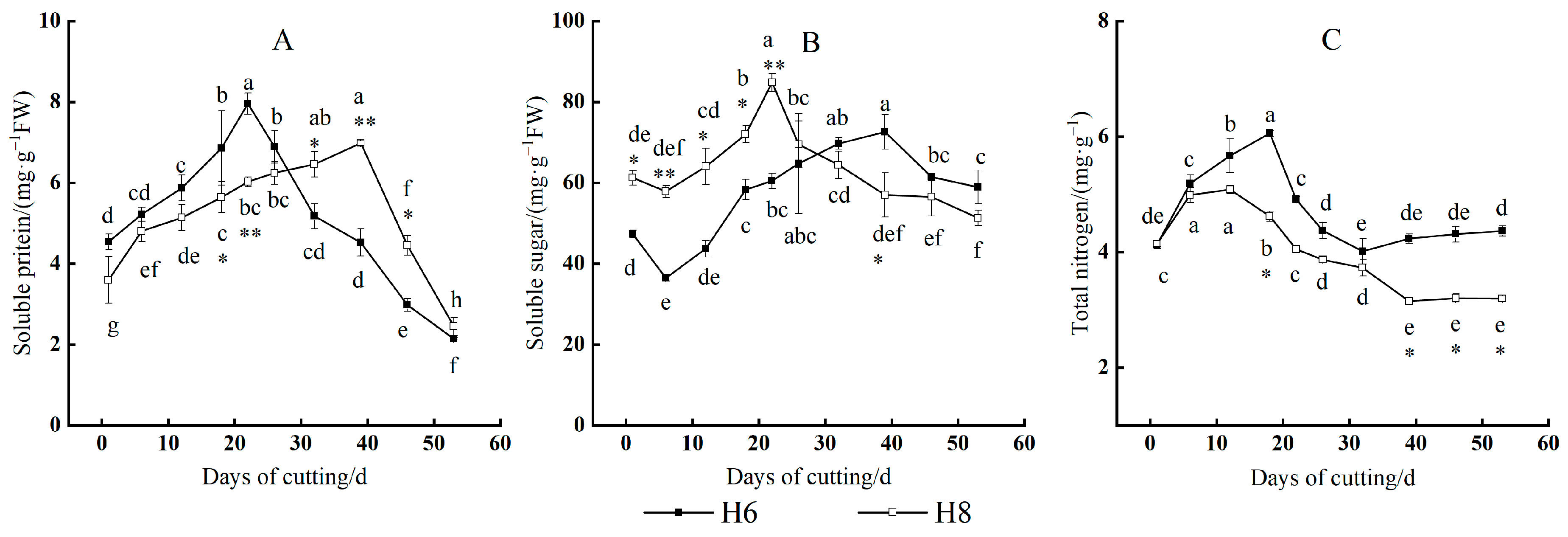
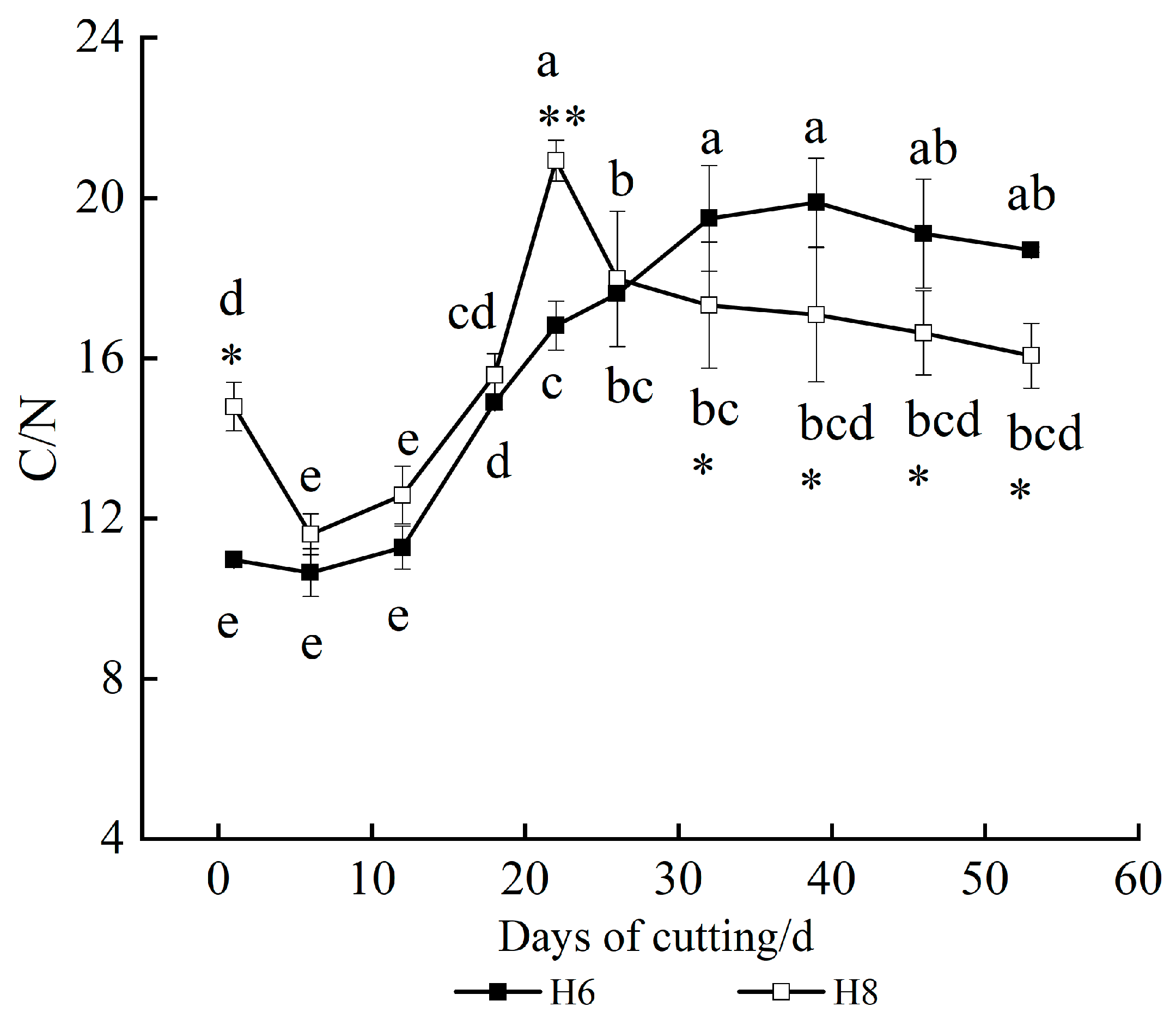
3.5. Changes in Nutrient Content and C/N Value
3.5.1. Changes in Nutrient Content
3.5.2. Change in C/N Value
4. Discussion
4.1. The Endogenous Hormones of Cutting Rooting
4.2. The Oxidase Activity and Rooting of E. ulmoides Cuttings
4.3. The Nutrients and Rooting of E. ulmoides Cuttings
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| IAA | Indole–3–acetic acid |
| ABA | Abscisic acid |
| GA3 | Gibberellic acid |
| ZR | Zeatin riboside |
| POD | Peroxidase |
| PPO | Polyphenol oxidase |
| IAAO | Indoleacetic acid oxidase |
| C/N | Carbon–nitrogen ratio |
| AR | Adventitious root |
References
- Wang, C.Y.; Tang, L.; He, J.W.; Li, J.; Wang, Y.Z. Ethnobotany, phytochemistry and pharmacological properties of Eucommia ulmoides: A review. Am. J. Chin. Med. 2019, 47, 259–300. [Google Scholar] [CrossRef]
- Kwan, C.Y.; Zhang, W.B.; Deyama, T.; Nishibe, S. In vitro vascular effects of Eucommia ulmoides Oliv.—A plausible scientific basis for its use in the elimination of blood stagnation in traditional Chinese medicine (Kanpo). Int. Symp. Eucommia Ulmoides 2007, 1, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Nakazawa, Y.; Toda, Y. Eucommia ulmoides Oliv. (Eucommiaceae): In Vitro Culture and the Production of Iridoids, Lignans, and Other Secondary Metabolites. Biotechnol. Agric. For. 1995, 1, 215–231. [Google Scholar]
- Hou, J.T.; Shen, C.C.; Zhang, Y.F.; Lin, N. Review on rooting mechanism of plant cuttings propagation. J. Anhui Agric. Sci. 2019, 47, 1–3. [Google Scholar]
- Wang, Z.M.; Cao, J.; Zhang, Y.P.; Bai, J.; Liu, G.B.; Liao, T.; Zheng, S.Q. Study on cutting propagation of four superior Sabina chinensis varieties and one clone. J. Cent. South Univ. For. Technol. 2019, 39, 62–69. [Google Scholar]
- Wang, Q.; Zhang, J.; Zhong, C.L.; Zhang, Y.; Wei, Y.C.; Meng, J.X. Variation of endogenesis hormone and nutritive matter concentration in Chukrasia tabularis cuttings during rooting. J. Cent. South Univ. For. Technol. 2020, 40, 111–119. [Google Scholar]
- Ouyang, F.Q.; Fu, G.Z.; Wang, J.H.; Ma, J.H.; An, S.P.; Wang, M.Q.; Li, Y. Qualitative analysis of endogenesis hormone and polyphenol during rooting of cuttings in Norway Spruce (Picea abies). Sci. Silvae Sin. 2015, 51, 155–162. [Google Scholar]
- Shang, C.Q.; Yang, H.; Ma, S.; Shen, Q.; Liu, L.; Hou, C.; Cao, X.; Cheng, J.L. Physiological and transcriptomic changes during the early phases of adventitious root formation in mulberry stem hardwood cuttings. Int. J. Mol. Sci. 2019, 20, 3707. [Google Scholar] [CrossRef] [Green Version]
- Li, S.W. Molecular bases for the regulation of adventitious root generation in plants. Front. Plant Sci. 2021, 12, 614072. [Google Scholar] [CrossRef]
- Zhou, Y.C.; Zhong, Q.P.; Li, Q.P.; Liu, W.F.; Zou, Y.L.; Tian, F.; Ge, X.N.; Cao, L.Q. Study on cutting propagation and rooting mechanism of semi–lignified spring shoots of Vernicia montana. J. Cent. South Univ. For. Technol. 2020, 40, 25–36. [Google Scholar]
- Zheng, Q.Q.; Lan, S.R.; Liu, X.D.; Feng, Y.C.; Peng, D.H. Effects of growth regulators on rooting and correlative enzyme activities of Michelia figo ’Xiangfei’ cuttings. J. Cent. South Univ. For. Technol. 2020, 40, 67–76. [Google Scholar]
- Guo, S.J.; Ling, H.Q.; Li, F.L. Physiological and biochemical basis of rooting of Pinus bun–geana cuttings. J. Beijing For. Univ. 2004, 2, 43–47. [Google Scholar]
- Li, H.S.; Sun, Q.; Zhao, S.J. Principle and Technology of Plant Physiological and Biochemical Experiment; Higher Education Press: Beijing, China, 2000; Volume 164–168, pp. 186–197. [Google Scholar]
- Truskina, J.; Han, J.; Chrysanthou, E.; Galvan–Ampudia, C.S.; Laine, S.; Brunoud, G.; Mace, J.; Bellows, S.; Legrand, J.; Bagman, A.M.; et al. A network of transcriptional repressors modulates auxin responses. Nature 2021, 589, 116–119. [Google Scholar] [CrossRef]
- Herud–sikimic, O.; Stiel, A.C.; Kolb, M.; Shanmugaratnam, S.; Berendzen, K.W.; Feldhaus, C.; Hocker, B.; Jurgens, G. A biosensor for the direct visualization of auxin. Nature 2021, 592, 768–772. [Google Scholar] [CrossRef] [PubMed]
- Marhava, P.; Bassukas, A.; Zourelidou, M.; Kolb, M.; Moret, B.; Fastner, A.; Schulze, W.X.; Cattaneo, P.; Hammes, U.Z.; Schwechheimer, C.; et al. A molecular rheostat adjusts auxin flux to promote root protophloem differentiation. Nature 2018, 558, 297–300. [Google Scholar] [CrossRef]
- Shi, F.H.; Zhao, R.; Luo, S.; Zheng, C.; Shen, Y.B. Study on plant hormone changes in soft cuttings of Tilia miqueliana during rooting. J. Cent. South Univ. For. Technol. 2019, 39, 21–26. [Google Scholar]
- Naoki, N.; Masatoshi, O.; Akiyoshi, K. Chemical screening for promotion of adventitious root formation in Eucalyptus globulus. BMC Proc. 2011, 5, 139. [Google Scholar]
- Guan, L.; Li, Y.; Huang, K.; Cheng, Z.M. Auxin regulation and MdPIN expression during adventitious root initiation in apple cuttings. Hortic. Res. 2020, 7, 143. [Google Scholar] [CrossRef]
- Chi, Y.; Hou, Y.J.; Yu, Y.J.; Yu, D.Y.; Feng, B.M.; Yao, Z.A.; Jiang, G.B. Dynamic changes of endogenous hormones contents in mutant of Populus tomentosaand and its relative species in the cutting and rooting procedure. J. Anhui Agric. Sci. 2007, 35, 8438–8439, 8442. [Google Scholar]
- Hanno, W.; Gerd, J. Survival of the flexible: Hormonal growth control and adaptation in plant development. Nat. Rev. Genet. 2009, 10, 306–317. [Google Scholar] [CrossRef]
- Han, J.H.; Shao, W.H.; Liu, J.F.; Diao, S.F. Content changes of endogenous hormone and polyphenols during hardwood cuttage progress in Sapindus mukorossi. Nonwood For. Res. 2019, 37, 37–43. [Google Scholar]
- Zhao, Y.L.; Chen, X.; Li, C.J. Dynamic of physiology and biochemistry during wild Rhododendron scabrifolium cutting propagation. Sci. Silvae Sin. 2013, 49, 45–51. [Google Scholar]
- Song, Y.; Chen, P.; Xuan, A.; Bu, C.; Liu, P.; Ingvarsson, P.K.; El–Kassaby, Y.A.; Zhang, D. Integration of genome wide association studies and co–expression networks reveal roles of PtoWRKY 42–PtoUGT76C1–1 in trans–zeatin metabolism and cytokinin sensitivity in poplar. New. Phytol. 2021, 231, 1462–1477. [Google Scholar] [CrossRef] [PubMed]
- Kuroha, T.; Kato, H.; Asami, T.; Yoshida, S.; Kamada, H.; Satoh, S. A trans–zeatin riboside in root xylem sap negatively regulates adventitious root formation on cucumber hypocotyls. J. Exp. Bot. 2002, 53, 2193–2200. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.W.; Guo, S.J.; Wang, G.Y. Effects of in vitro subculture on the physiological characteristics of adventitious root formation in microshoots of Castanea mollissima cv. ‘yanshanhong’. J. For. Res. 2010, 21, 155–160. [Google Scholar] [CrossRef]
- Lomin, S.N.; Krivosheev, D.M.; Steklov, M.Y.; Osolodkin, D.I. Romanov GA. Receptor properties and features of cytokinin signaling. Acta Nat. 2012, 4, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.M.; Zhu, L.Y.; Chen, Y.; Yao, X.W.; Bai, J.W.; Chen, X.; Zhang, Q.Q.; Cao, F.L. Study on rooting process and rooting mechanism of hardwood cuttings of Ginkgo biloba ’Golden–leaf’. J. Cent. South Univ. For. Technol. 2020, 40, 28–37. [Google Scholar]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Bian, B.; Gong, T.; Liao, W. Comparative proteomic analysis of key proteins during abscisic acid–hydrogen peroxide–induced adventitious rooting in cucumber (Cucumis sativus L.) under drought stress. J. Plant Physiol. 2018, 229, 185–194. [Google Scholar] [CrossRef]
- Wu, W.H.; Cao, F.; Liu, Z.Z.; Peng, F.R.; Liang, Y.W.; Tang, P.P. Effects of NAA treatment on the endogenous hormone changes in cuttings of Carya illinoinensis during rooting. J. Nanjing For. Nat. Sci. Ed. 2016, 40, 191–196. [Google Scholar]
- Nag, S.; Saha, K.; Choudhuri, M.A. Role of auxin and polyamines in adventitious root formation in relation to changes in compounds involved in rooting. J. Plant Growth Regul. 2001, 20, 182–194. [Google Scholar] [CrossRef]
- Muhammad, Z.M.; Saleem, U.D.; Muhammad, I.; Zhang, Z.J. Transcriptomic and Anatomic Profiling Reveal Etiolation Promotes Adventitious Rooting by Exogenous Application of 1–Naphthalene Acetic Acid in Robinia pseudoacacia L. Forests 2021, 12, 789. [Google Scholar] [CrossRef]
- Skoog, F.; Miller, C.O. Chemical regulation of growth and organ formation in plant tissues cultured in vitro. Symp. Soc. Exp. Biol. 1957, 11, 118–130. [Google Scholar] [PubMed]
- Li, K.; Liang, Y.; Xing, L.; Mao, J.; Liu, Z.; Dong, F.; Meng, Y.; Han, M.; Zhao, C.; Bao, L.; et al. Transcriptome analysis reveals multiple hormones, wounding and sugar signaling pathways mediate adventitious root formation in apple rootstock. Int. J. Mol. Sci. 2018, 19, 2201. [Google Scholar] [CrossRef] [Green Version]
- Mao, J.; Zhang, D.; Meng, Y.; Li, K.; Wang, H.; Han, M. Inhibition of adventitious root development in apple rootstocks by cytokinin is based on its suppression of adventitious root primordia formation. Physiol. Plant. 2019, 166, 663–676. [Google Scholar] [CrossRef]
- Shang, W.Q.; Wang, Z.; He, S.L.; He, D.; Dong, N.L.; Guo, Y. Changes of endogenous IAA and related enzyme activities during rooting of Paeonia suffruticosa in vitro. J. Northwest A F Univ. Nat. Sci. Ed. 2021, 49, 129–136. [Google Scholar]
- Zhang, W.X.; Fan, J.J.; Tan, Q.Q.; Zhao, M.M.; Zhou, T.; Cao, F.L. The effects of exogenous hormones on rooting process and the activities of key enzymes of Malus hupehensis stem cuttings. PLoS ONE 2017, 12, e0172320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, L.N. Comparison of Rooting Ability of Different Genotypes of Sorbus and Determination of Physiology and Biochemistry; Liaoning Normal University: Dalian, China, 2017. [Google Scholar]
- Gyana, R.R. Effect of Auxins on Adventitious Root Development from Single Node Cuttings of Camellia sinensis (L.) Kuntze and Associated Biochemical Changes. Plant Growth Regul. 2006, 48, 111–117. [Google Scholar]
- Bassuk, N.L.; Hunter, L.D.; Howard, B.H. The apparent involvement of polyphenol oxidase and phloridzin in the production of apple rooting cofactors. J. Hortic. Sci. 2015, 56, 313–322. [Google Scholar] [CrossRef]
- Yan, S.P.; Wu, X.D.; Wang, Q.Y.; Song, X.S.; Yang, R.H. Changes in relevant oxidase activities during the rooting of cuttings of Populus tremula × P. tremuloides and cutting propagation technique. J. Northeast For. Univ. 2011, 39, 5–7. [Google Scholar]
- Kose, C.; Erdal, S.; Kaya, O.; Atici, O. Comparative evaluation of oxidative enzyme activities during adventitious rooting in the cuttings of grapevine rootstocks. J. Sci. Food Agric. 2011, 91, 738–741. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Liu, X.; Yuan, Y. Water cultured propagation of Polygonum multiflorum and dynamic changes of physiological and biochemical characteristics during adventitious roots formation. China J. Chin. Mater. Med. 2011, 36, 375–378. [Google Scholar]
- Song, J.Y.; Liu, Y.J.; Song, G.; Wang, G.F.; Wang, H.S. Changes of peroxidase and IAA oxidase activities and phenol content in the cutting root course of several familiar tree species. Jiangsu Agric. Sci. 2007, 6, 115–118. [Google Scholar]
- Wang, X.J.; He, W.; Zhang, Q.J.; Wang, N.; Zhu, Y.J.; Zhu, Y.L. Change of nutrient content and oxidase activities during rooting of Catalpa bungei cl. ‘Yu–1’. Sci. Silvae Sin. 2009, 45, 156–161. [Google Scholar]
- Shao, F.; Wang, S.; Huang, W.; Liu, Z. Effects of IBA on the rooting of branch cuttings of Chinese jujube (Zizyphus jujuba Mill.) and changes to nutrients and endogenous hormones. J. For. Res. 2018, 29, 1557–1567. [Google Scholar] [CrossRef]
- Druege, U.; Hilo, A.; Perez–Perez, J.M.; Klopotek, Y.; Acosta, M.; Shahinnia, F.; Zerche, S.; Franken, P.; Hajirezaei, M.R. Molecular and physiological control of adventitious rooting in cuttings: Phytohormone action meets resource allocation. Ann. Bot. 2019, 123, 929–949. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.M.; Fang, W.M.; Chen, F.D.; Chen, S.M. Cuttings rooting process and changes in carbohydrate, nitrogen and endogenous hormone levels during the rooting of two chrysanthemum cultivars. J. Nanjing Agric. Univ. 2010, 33, 19–24. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material | Rooting Rate (%) | Average Length of Root (cm) | Length of Longest Root (cm) | No. of Root per Shoot | * Effect of Rooting |
|---|---|---|---|---|---|
| H8 | 22.53 ± 1.97 a | 9.55 ± 4.54 a | 13.75 ± 6.55 a | 15.25 ± 3.79 a | 145.64 |
| H6 | 85.30 ± 2.52 b | 8.41 ± 2.57 a | 13.30 ± 2.36 a | 25.60 ± 4.01 b | 215.3 |
| CK | 21.3 ± 1.53 a | 4.33 ± 1.17 c | 8.3 ± 1.76 b | 5.35 ± 1.68 c | 23.17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, G.; Qing, J.; Du, H.; Du, Q.; Meng, Y.; He, F.; Liu, P.; Du, L.; Wang, L. Comparing Rooting Ability and Physiological Changes of Two Eucommia ulmoides Improved Varieties. Forests 2021, 12, 1267. https://doi.org/10.3390/f12091267
Lv G, Qing J, Du H, Du Q, Meng Y, He F, Liu P, Du L, Wang L. Comparing Rooting Ability and Physiological Changes of Two Eucommia ulmoides Improved Varieties. Forests. 2021; 12(9):1267. https://doi.org/10.3390/f12091267
Chicago/Turabian StyleLv, Gengxin, Jun Qing, Hongyan Du, Qingxin Du, Yide Meng, Feng He, Panfeng Liu, Lanying Du, and Lu Wang. 2021. "Comparing Rooting Ability and Physiological Changes of Two Eucommia ulmoides Improved Varieties" Forests 12, no. 9: 1267. https://doi.org/10.3390/f12091267
APA StyleLv, G., Qing, J., Du, H., Du, Q., Meng, Y., He, F., Liu, P., Du, L., & Wang, L. (2021). Comparing Rooting Ability and Physiological Changes of Two Eucommia ulmoides Improved Varieties. Forests, 12(9), 1267. https://doi.org/10.3390/f12091267