
Mangrove Dieback and Leaf Disease in Sonneratia apetala and Sonneratia caseolaris in Vietnam

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Canopy Assessment
2.2. Sample Collection
2.3. Fungal Isolation
2.4. Fungal Identification
- DNA extraction, PCR amplification, and sequencing analysis
- Phylogenetic analyses
2.5. Pathogenicity Tests
2.6. Statistical Analysis
3. Results
3.1. Tree Health Survey
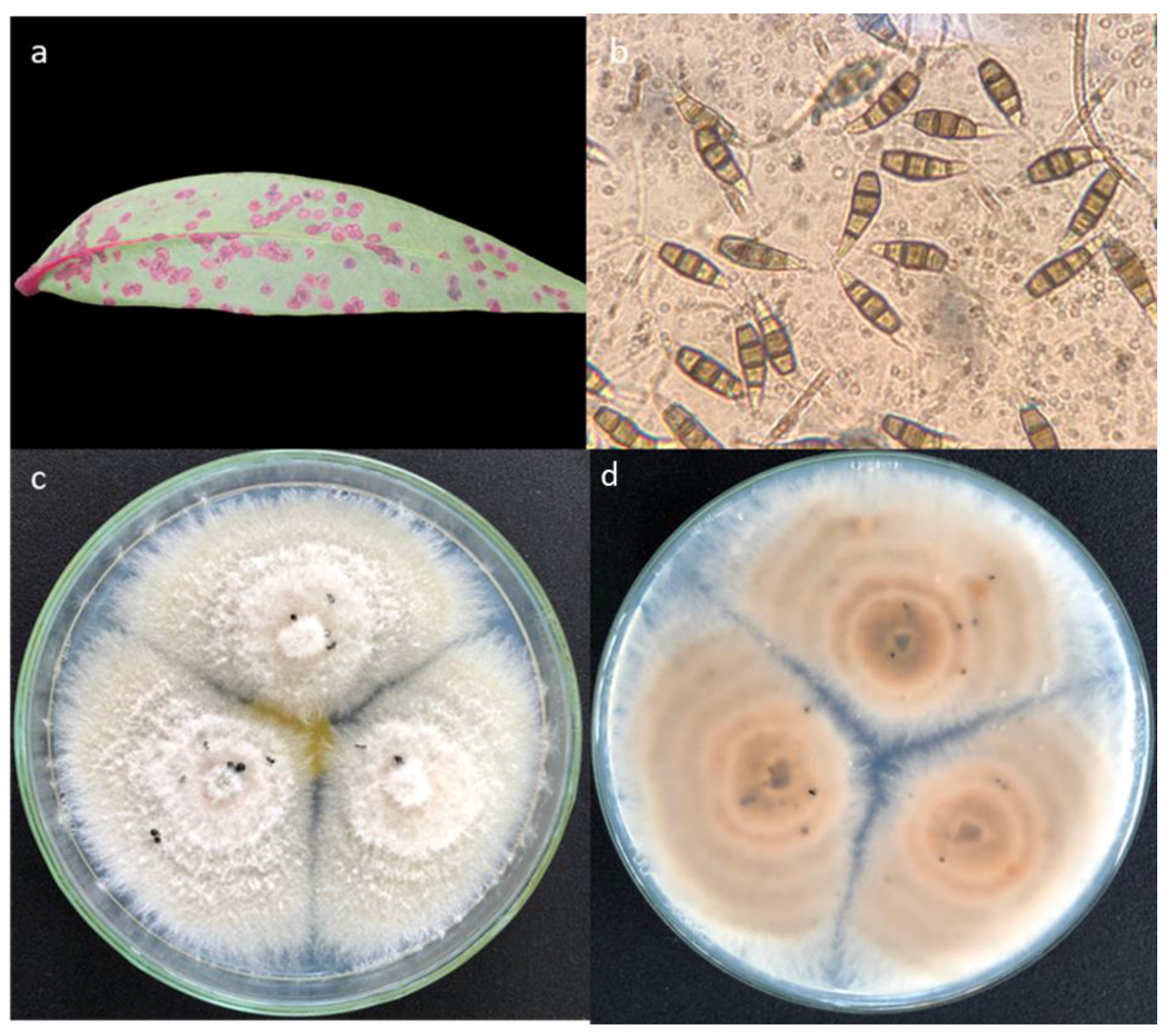
3.2. Symptoms of Disease on Sonneratia Species
3.3. Taxonomy
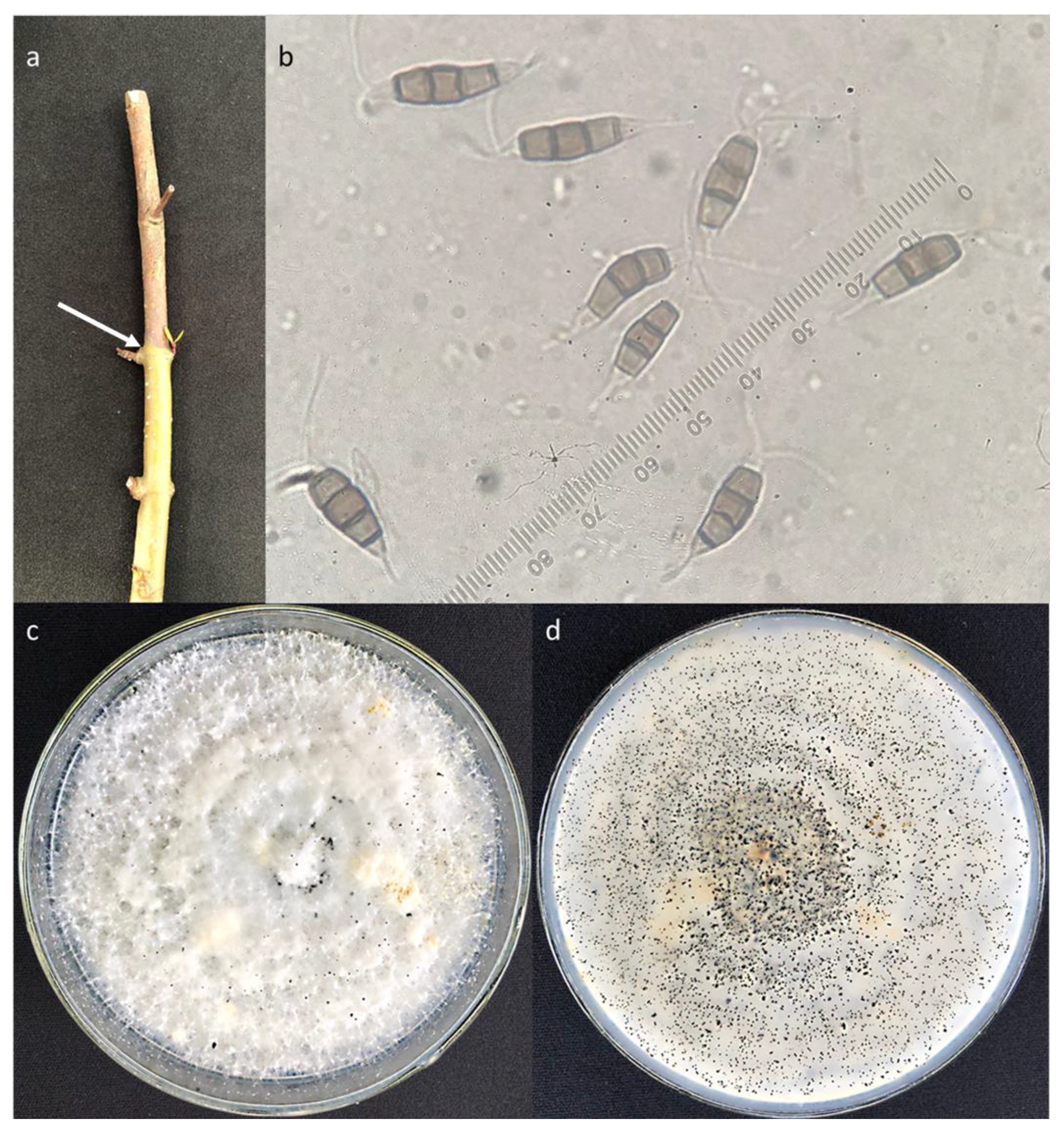
3.4. Morphology of the Main Taxa
- Curvularia aff. tsudae
- Neopestalotiopsis sp.1
- Pestalotiopsis sp.1
- Pestalotiopsis sp.4a
3.5. Pathogenicity Trial
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sherman, R.E.; Fahey, T.J.; Martinez, P. Hurricane Impacts on a Mangrove Forest in the Dominican Republic: Damage Patterns and Early Recovery 1. Biotropica 2001, 33, 393–408. [Google Scholar] [CrossRef]
- Kathiresan, K.; Bingham, B.L. Biology of Mangroves and Mangrove Ecosystems. Adv. Mar. Biol. 2001, 40, 81–251. [Google Scholar]
- Taylor, M.; Ravilious, C.; Green, E.P. Mangroves of East Africa; UNEP–WCMC: Cambridge, UK, 2003. [Google Scholar]
- Martinuzzi, S.; Gould, W.A.; Lugo, A.E.; Medina, E. Conversion and Recovery of Puerto Rican Mangroves: 200 years of change. For. Ecol. Manag. 2009, 257, 75–84. [Google Scholar] [CrossRef]
- Abuodha, P.; Kairo, J. Human-induced Stresses on Mangrove Swamps along the Kenyan Coast. Hydrobiologia 2001, 458, 255–265. [Google Scholar] [CrossRef]
- Ellison, A.M. Mangrove Communities. In Marine Community Ecology; Sinauer Associates, Inc.: Sunderland, MA, USA, 2001; pp. 423–442. [Google Scholar]
- Spalding, M. World Atlas of Mangroves; Routledge: London, UK, 2010. [Google Scholar]
- Smith, T.J., III; Robblee, M.B.; Wanless, H.R.; Doyle, T.W. Mangroves, Hurricanes, and Lightning Strikes: Assessment of Hurricane Andrew Suggests an Interaction across Two Differing Scales of Disturbance. BioScience 1994, 44, 256–262. [Google Scholar] [CrossRef]
- Norm, D.; Jock, M.; John, K.; Grant, S.; Rob, C.; Apanie, W.; Ysabel, C. Assessing the Gulf of Carpentaria Mangrove Dieback 2017–2019; James Cook University: Douglas, Australia, 2020; p. 249. [Google Scholar]
- Van, O.P.; Duke, N. Extreme Weather Likely behind Worst Recorded Mangrove Dieback in Northern Australia. Conversation 2017, 14, 1–6. [Google Scholar]
- Gilbert, G.S. Evolutionary Ecology of Plant Diseases in Natural Ecosystems. Annu. Rev. Phytopathol. 2002, 40, 13–43. [Google Scholar] [CrossRef] [Green Version]
- Hulcr, J.; Dunn, R.R. The Sudden Emergence of Pathogenicity in Insect–fungus Symbioses Threatens Naive Forest Ecosystems. Proc. R. Soc. B Biol. Sci. 2011, 278, 2866–2873. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.; Cannon, P. Polystigma sonneratiae Causing Leaf Spots on The Mangrove Genus Sonneratia. Aust. Syst. Bot. 1992, 5, 415–420. [Google Scholar] [CrossRef]
- Jones, E.G.; Pang, K.-L. Tropical Aquatic Fungi. Biodivers. Conserv. 2012, 21, 2403–2423. [Google Scholar] [CrossRef]
- Jones, E.G.; Pang, K.-L.; Abdel-Wahab, M.A.; Scholz, B.; Hyde, K.D.; Boekhout, T.; Ebel, R.; Rateb, M.E.; Henderson, L.; Sakayaroj, J. An Online Resource for Marine Fungi. Fungal Divers. 2019, 96, 347–433. [Google Scholar] [CrossRef]
- Alias, S.; Jones, E. Colonization of Mangrove Wood by Marine Fungi at Kuala Selangor Mangrove Stand, Malaysia. Fungal Divers. 2000, 5, 9–21. [Google Scholar]
- Kohlmeyer, J.; Kohlmeyer, E. Marine Fungi From Tropical America and Africa. Mycologia 1971, 63, 831–861. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.G.; Abdel-Wahab, M.A. Marine Fungi from the Bahamas Islands. Bot. Mar. 2005, 48, 356–364. [Google Scholar]
- Sarma, V.; Hyde, K.D. A review on Frequently Occurring Fungi in Mangroves. Fungal Divers 2001, 8, 1–34. [Google Scholar]
- Steinke, T. Mangrove Fungi on Dead Proproots of Rhizophora mucronata at Three Localites in South Africa. S. Afr. J. Bot. 2000, 66, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Osorio, J.A.; Wingfield, M.J.; Roux, J. A Review of Factors Associated with Decline and Death of Mangroves, with Particular Reference to Fungal Pathogens. S. Afr. J. Bot. 2016, 103, 295–301. [Google Scholar] [CrossRef]
- Xie, S.; He, H.; Yang, R.; Xu, Z.; He, J.; Lu, H. First Report of Leaf Spot Caused by Corynespora cassiicola on Acanthus ilicifolius in China. Plant Dis. 2021, 105, 509. [Google Scholar] [CrossRef]
- Stevens, F.L. New or Noteworthy Porto Rican Fungi. Bot. Gaz. 1920, 70, 399–402. [Google Scholar] [CrossRef]
- Rahman, M.A. Top Dying of Sundri (Heritiera fomes) and Its Impact on the Regeneration and Management in the Mangrove Forests of Sunderbans in Bangladesh. In Proceedings of the Impact of Diseases and Insect Pests in Tropical Forests, Peechi City, India, 23–26 November 1993; pp. 117–133. [Google Scholar]
- Yan, T.L.; Zhu, X.J.; Yang, Z.D.; Long, Q.H.; Yang, H.; Chang, S.Q. New Records of Celoporthe guangdongensis and Cytospora rhizophorae on Mangrove Apple in China. Biodivers. Data J. 2020, 8, 15. [Google Scholar]
- McMillan, J.R. Studies of a Recently Described Cercospora on Rhizophora mangle. Plant Dis. Report. 1964, 48, 909–991. [Google Scholar]
- Creager, D.B. A New Cercospora on Rhizophora mangle. Mycologia 1962, 54, 536–539. [Google Scholar] [CrossRef]
- Osorio, J.A.; Crous, C.J.; Wingfield, M.J.; De Beer, Z.W.; Roux, J. An assessment of Mangrove Diseases and Pests in South Africa. For. Int. J. For. Res. 2017, 90, 343–358. [Google Scholar] [CrossRef] [Green Version]
- Osorio, J.A.; Crous, C.J.; De Beer, Z.W.; Wingfield, M.J.; Roux, J. Endophytic Botryosphaeriaceae, Including Five New Species, Associated with Mangrove Trees in South Africa. Fungal Biol. 2017, 121, 361–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wier, A.M.; Tattar, T.A.; Klekowski, E.J., Jr. Disease of Red Mangrove (Rhizophora mangley) in Southwest Puerto Rico Caused by Cytospora rhizophorae 1. Biotropica 2000, 32, 299–306. [Google Scholar]
- Kohlmeyer, J. Ecological Notes on Fungi in Mangrove Forests. Trans. Br. Mycol. Soc. 1969, 53, 237–250. [Google Scholar] [CrossRef]
- Teas, H.; McEwan, R. An Epidemic Dieback Gall Disease of Rhizophora mangroves in The Gambia, West Africa. Plant Dis. 1982, 66, 522–523. [Google Scholar] [CrossRef]
- Sakayaroj, J.; Preedanon, S.; Suetrong, S.; Klaysuban, A.; Jones, E.G.; Hattori, T. Molecular Characterization of Basidiomycetes Associated with The Decayed Mangrove Tree Xylocarpus granatum in Thailand. Fungal Divers. 2012, 56, 145–156. [Google Scholar] [CrossRef]
- Pegg, K.; Gillespie, N.; Forsberg, L. Phytophthora sp. Associated with Mangrove Death in Central Coastal Queensland. Australas. Plant Pathol. 1980, 9, 6–7. [Google Scholar] [CrossRef]
- Xu, J.; Kjer, J.; Sendker, J.; Wray, V.; Guan, H.; Edrada, R.; Lin, W.; Wu, J.; Proksch, P. Chromones From the Endophytic Fungus Pestalotiopsis sp. Isolated from the Chinese Mangrove Plant Rhizophora mucronata. J. Nat. Prod. 2009, 72, 662–665. [Google Scholar] [CrossRef]
- Olexa, M.; Freeman, T. Occurrence of Three Unrecorded Diseases on Mangroves in Florida. In Proceedings of the International Symposium on Biology and Management of Mangroves, Honolulu, HI, USA, 8–11 October 1974; pp. 688–692. [Google Scholar]
- Shivas, R.; Young, A.; Crous, P. Pseudocercospora avicenniae RG Shivas, AJ Young & Crous, sp. nov. Persoonia 2009, 23, 193. [Google Scholar]
- Rafael, A.; Calumpong, H.P. Fungal Infections of Mangroves in Natural Forests and Reforestation Sites from Philippines. Aquac. Aquar. Conserv. Legis. 2019, 12, 2062–2074. [Google Scholar]
- Hong, P.N.; San, H.T. Mangroves of Vietnam; IUCN: Bangkok, Thailand, 1993; pp. 150–180.
- Ministry of Agriculture and Rural Development. Announcement of National Forest Status in 2019; MARD: Hanoi, Vietnam, 2020.
- Hai, N.; Dell, B.; Phuong, V.; Harper, R. Towards a More Robust Approach for The Restoration of Mangroves in Vietnam. Ann. For. Sci. 2020, 77, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.H.; Nghia, N.H.; Nguyen, H.T.T.; Le, A.T.; Tran, L.T.N.; Duong, L.V.K.; Bohm, S.; Furniss, M.J. Classification Methods for Mapping Mangrove Extents and Drivers of Change in Thanh Hoa Province, Vietnam During 2005–2018. For. Soc. 2020, 4, 225–242. [Google Scholar] [CrossRef]
- Vietnam Admistration of Forestry. Protective Forest in Coastal Is Unexplained Death. Available online: http://bentre.tintuc.vn/tin-tuc/ben-tre-rung-phong-ho-dac-dung-ven-bien-chet-chua-ro-nguyen-nhan.html (accessed on 1 July 2021).
- Baotainguyenvamoitruong. Proposing Scientific Research to Find the Cause of Death of Mangroves in Ben Tre. Available online: https://baotainguyenmoitruong.vn/moi-truong/ben-tre-de-xuat-nghien-cuu-khoa-hoc-tim-nguyen-nhan-rung-chet-1258495.html (accessed on 1 July 2021).
- Sggp.org.vn. Death of Rhizophora in Mass: Identify reasons (Vietnamese). Available online: http://www.sggp.org.vn/vu-cay-duoc-chet-hang-loat-xac-dinh-loai-sinh-vat-la-gay-hai-451729.html (accessed on 1 July 2021).
- Antv.gov.vn. Death of Mangroves in Mass in Ha Tinh (Vietnamese). Available online: http://www.antv.gov.vn/tin-tuc/xa-hoi/rung-cay-ngap-man-o-ha-tinh-chet-hang-loat-186342.html (accessed on 1 July 2021).
- Nguyen, H.T.T.; Hardy, G.E.; Le, T.V.; Nguyen, H.Q.; Nguyen, H.H.; Nguyen, T.V.; Dell, B. Mangrove Forest Landcover Changes in Coastal Vietnam: A Case Study from 1973 to 2020 in Thanh Hoa and Nghe An Provinces. Forests 2021, 12, 637. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Khan, A.M.; Bhadauria, S. Molecular Characterization of Keratin Degrading Fungi Isolated from Semi-arid Soil by PCR Using ITS4 and ITS5 Primers. J. King Saud Univ.-Sci. 2019, 31, 1418–1423. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: New York, NY, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis Across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Allen, P.; Bennett, K.; Heritage, B. SPSS Statistics Version 22: A Practical Guide; Cengage Learning Australia Pty Limited: Sydney, Australia, 2014; pp. 200–250. [Google Scholar]
- Shamsi, S.; Hosen, S.; Ahmed, A. Fungi Associated with Leaves of Sonneratia apetala Buch. Ham and Sonneratia caseolaris (L.) Engler from Rangabali Coastal Zone of Bangladesh. Dhaka Univ. J. Biol. Sci. 2018, 27, 155–162. [Google Scholar] [CrossRef]
- Abbas, M.F.; Rafiq, M.; Al-Sadi, A.M.; Alfarraj, S.; Alharbi, S.A.; Arif, M.; Ansari, M.J. Molecular Characterization of Leaf Spot caused by Alternaria alternata on Buttonwood (Conocarpus erectus L.) and Determination of Pathogenicity by a Novel Disease Rating Scale. PLoS ONE 2021, 16, e0251471. [Google Scholar] [CrossRef]
- Tattar, T.; Klekowski, E.; Stern, A. Dieback and Mortality in Red Mangrove, Rhizophora mangle L., in Southwest Puerto Rico. Arboric. J. 1994, 18, 419–429. [Google Scholar] [CrossRef]
- Liu, A.; Wu, X.; Xu, T. Research advances in endophytic fungi of mangrove. J. Appl. Ecol. 2007, 18, 912–918. [Google Scholar]
- Suryanarayanan, T.; Kumaresan, V. Endophytic fungi of some halophytes from an estuarine mangrove forest. Mycol. Res. 2000, 104, 1465–1467. [Google Scholar] [CrossRef]
- Chaeprasert, S.; Piapukiew, J.; Whalley, A.J.; Sihanonth, P. Endophytic Fungi from Mangrove Plant Species of Thailand: Their Antimicrobial and Anticancer Potentials. Bot. Mar. 2010, 53, 555–564. [Google Scholar] [CrossRef]
- Xing, X.; Guo, S. Fungal Endophyte Communities in four Rhizophoraceae Mangrove Species on The South Coast of China. Ecol. Res. 2011, 26, 403–409. [Google Scholar] [CrossRef]
- Khan, A.A.H. Endophytic Fungi and Their Impact on Agroecosystems. In Medicinal Plants: Biodiversity, Sustainable Utilization and Conservation; Springer: Singapore, 2020; pp. 443–499. [Google Scholar]
- Nurunnabi, T.R.; Sabrin, F.; Sharif, D.I.; Nahar, L.; Sohrab, M.H.; Sarker, S.D.; Rahman, S.M.; Billah, M.M. Antimicrobial Activity of Endophytic Fungi Isolated From the Mangrove Plant Sonneratia apetala (Buch.-Ham) from the Sundarbans Mangrove Forest. Adv. Tradit. Med. 2020, 40, 1–7. [Google Scholar] [CrossRef]
- Goudarzi, A.; Moslehi, M. Distribution of a Devastating Fungal Pathogen in Mangrove Forests of Southern Iran. Crop Prot. 2020, 128, 104987. [Google Scholar] [CrossRef]
- Debbab, A.; Aly, A.H.; Proksch, P. Mangrove Derived Fungal Endophytes–a Chemical and Biological Perception. Fungal Divers. 2013, 61, 1–27. [Google Scholar] [CrossRef]
- Ebrahim, W.; Aly, A.H.; Mándi, A.; Totzke, F.; Kubbutat, M.H.; Wray, V.; Lin, W.H.; Dai, H.; Proksch, P.; Kurtán, T. Decalactone Derivatives from Corynespora cassiicola, an Endophytic Fungus of the Mangrove Plant Laguncularia racemosa. Eur. J. Org. Chem. 2012, 2012, 3476–3484. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.S.; Tang, W.C.; Xu, S.L.; Sun, S.F.; Huang, B.Y.; Yan, X.; Chen, Q.J.; Lin, Y.C. First Report of An endophyte (Diaporthe phaseolorum var. sojae) from Kandelia candel. J. For. Res. 2008, 19, 277–282. [Google Scholar] [CrossRef]
- Xing, X.; Chen, J.; Xu, M.; Lin, W.; Guo, S. Fungal Endophytes Associated with Sonneratia (Sonneratiaceae) Mangrove Plants on the South Coast of China. For. Pathol. 2011, 41, 334–340. [Google Scholar] [CrossRef]
- Olexa, M.; Freeman, T. A Gall Disease of Red Mangrove Caused by Cylindrocarpon didymum. Plant Dis. Report. 1978, 62, 283–286. [Google Scholar]
- Chi, W.-C.; Chen, W.; He, C.-C.; Guo, S.-Y.; Cha, H.-J.; Tsang, L.M.; Ho, T.W.; Pang, K.-L. A Highly Diverse Fungal Community Associated with Leaves of the Mangrove Plant Acanthus ilicifolius var. xiamenensis Revealed by Isolation and Metabarcoding Analyses. PeerJ 2019, 7, e7293. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.; Lin, Y.; Luo, M.; Lu, Y.; Huang, X.; She, Z. Diaporisoindoles A–C: Three isoprenylisoindole alkaloid derivatives from the mangrove endophytic fungus Diaporthe sp. SYSU-HQ3. Org. Lett. 2017, 19, 5621–5624. [Google Scholar] [CrossRef] [PubMed]
- Norphanphoun, C.; Jayawardena, R.; Chen, Y.; Wen, T.; Meepol, W.; Hyde, K. Morphological and Phylogenetic Characterization of Novel Pestalotioid Species Associated with Mangroves in Thailand. Mycosphere 2019, 10, 531–578. [Google Scholar] [CrossRef]
- Hamzah, T.N.T.; Lee, S.Y.; Hidayat, A.; Terhem, R.; Faridah-Hanum, I.; Mohamed, R. Diversity and Characterization of Endophytic Fungi Isolated from the Tropical Mangrove Species, Rhizophora mucronata, and Identification of Potential Antagonists Against the Soil-Borne Fungus, Fusarium solani. Front. Microbiol. 2018, 9, 1707. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Diao, X.; Wang, T.; Chen, G.; Lin, Q.; Yang, X.; Xu, J. Phylogenetic Diversity and Antioxidant Activities of Culturable Fungal Endophytes Associated with the Mangrove Species Rhizophora stylosa and R. mucronata in the South China Sea. PLoS ONE 2018, 13, e0197359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshmukh, S.K.; Gupta, M.K.; Prakash, V.; Reddy, M.S. Mangrove-Associated Fungi: A Novel Source of Potential Anticancer Compounds. J. Fungi 2018, 4, 101. [Google Scholar] [CrossRef] [Green Version]
- Ha, N.T.; Ngat, H.T.; Ha, N.T.T.; Hoi, P.X. Study on Pathogenicity of Colletotrichum gloeosporioides on Coffee in North of Vietnam. Biology 2011, 33, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Tran, N.B.B. Isolation and Identification of Fungal Diseases in Rice (Oryza sativa) from Mekong Delta. Ph.D. Thesis, International University-HCMC, Ho Chi Minh City, Vietnam, 2019. [Google Scholar]
- Ngoc, N.; Phong Nguyen, N.; An, P.; Woolf, A.; Fullerton, R. Effect of Storage Temperatures on Postharvest Diseases of Dragon Fruits (Hylocereus undatus Haw.) in the Mekong Delta Region, Vietnam. In Proceedings of the III Asia Pacific Symposium on Postharvest Research, Education and Extension: APS2014 1213, Hochiminh City, Vietnam, 8 December 2014; pp. 453–460. [Google Scholar]
- Yen, T.T.N.; Thanh, N.V.; Phong, N.V. Research on the Causative Agent of Rambutan fruit rot (Nephelium lappaceum L.) after Harvest in the Mekong Delta. J. Sci. Technol. 2017, 13, 1868–1873. [Google Scholar]
- Dung, L.; Dienb, N.T.; Daic, P.H.; Tuana, P.N. The First Report of Pestalofiopsis sp. Causing Crown Rot Disease on Strawberries in Dalat. Dalat Univ. J. Sci. 2016, 6, 364–376. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungus | Disease | Host | Location | Reference |
|---|---|---|---|---|
| Acanthus ilicifolius | leaf spot | Corynespora cassiicola | China | [22] |
| Anthostomella rhizomorphae | leaf spot | Rhizophora mangle | Puerto Rico | [23] |
| Botrosphaeria ribis | death of twigs and branches | Heritiera fomes | Bangladesh | [24] |
| Celoporthe guangdongensis | canker | Sonneratia apetala | China | [25] |
| Cercospora | leaf spot | Rhizophora mangle | Florida | [26] |
| Cercospora | leaf disease | Rhizophora mangle | Florida | [27] |
| Colletotrichum sp. | leaf spot | Rhizophora sp. | Australia | [13] |
| Cyphellophora sp. | stem canker, branch dieback | Avicennia marina | South Africa | [28,29] |
| Cytospora rhizophorae | dieback | Rhizophora mangle | Puerto Rico | [30] |
| Cytospora rhizophorae | canker | Sonneratia apetala | China | [25] |
| Cytospora sp. | dieback | Rhizophora mangle | Hawaii | [31] |
| Eutypella sp. | stem/branch canker | Avicennia marina | South Africa | [28,29] |
| Fulvifomes | heart/butt infections | Xylocarpus granatum | Thailand | [32] |
| Fulvifomes sp. | decay | Xylocarpus granatum | Thailand | [33] |
| Lasiodiplodia theobromae | branch dieback, branch/stem canker | Avicennia marina | South Africa | [28,29] |
| Oomycete | mortality | Avicennia marina | Australia | [34] |
| Pestalotiopsis sp. | leaf spot | Rhizophora mucronata | China | [35] |
| Phyllosticta hibiscina | necrosis and death | Avicennia germinans | Florida | [36] |
| Polystigma sonneratiae | leaf spot | Rhizophora sp. | Australia | [13] |
| Pseudocercospora avicenniae | leaf spot | Avicennia marina | Australia | [37] |
| Pseudocercospora mapelanensis | disease on fruit, flowers and leaves | Barringtonia racemosa | South Africa | [28] |
| Pseudocercosporella | leaf spot | Ceriops decandra, Sonneratia caseolaris, Rhizophora stylosa and Osbornia octodonta | Philippines | [38] |
| Location | Sonneratia Species | Year Planted | Preliminary Study | Main Study |
|---|---|---|---|---|
| Sam Son town, Thanh Hoa Province | S. apetala | 2015, 2016 | December 2018 | |
| S. caseolaris | 2015, 2016 | December 2018 | ||
| Hoang Phu District, Thanh Hoa Province | S. caseolaris | 2018 | December 2018 | August 2019 |
| S. apetala | 2018 | December 2018 | August 2019 |
| Rating | Dieback Criteria | Leaf Spot Criteria |
|---|---|---|
| 1 | No dead branches visible | Dense, healthy foliage; no leaf spots |
| 2 | Few dead twigs present; may have an occasional large branch stub on an upper bole | Dense foliage; spots (pink/black) present at low density on leaves |
| 3 | Many dead twigs present; an occasional large dead branch; may have large branch stubs on the upper bole | Foliage density appears subnormal; numerous pink or black spots present |
| Trees with Dieback (Damage Class) | Trees with Pink Spot (Damage Class) | Trees with Black Spot (Damage Class) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | |
| Mixed stands | |||||||||
| Sonneratia apetala | 1354 | 53 | 22 | 1415 | 12 | 2 | 926 | 487 | 16 |
| (94.8) | (3.7) | (1.5) | (99) | (0.8) | (0.1) | (64.8) | (34.1) | (1.1) | |
| Sonneratia caseolaris | 1129 | 107 | 157 | 929 | 438 | 26 | 1377 | 14 | 2 |
| (81) | (7.7) | (11.3) | (66.7) | (31.4) | (1.9) | (98.9) | (1) | (0.1) | |
| Single stands | |||||||||
| Sonneratia apetala | 885 | 37 | 12 | 925 | 7 | 2 | 663 | 253 | 18 |
| (94.8) | (4) | (1.3) | (99) | (0.7) | (0.2) | (71) | (27.1) | (1.9) | |
| Sonneratia caseolaris | 746 | 69 | 128 | 654 | 261 | 28 | 938 | 3 | 2 |
| (79.1) | (7.3) | (13.6) | (69.4) | (27.7) | (3) | (99.5) | (0.3) | (0.2) | |
| Total | |||||||||
| Sonneratia apetala | 2239 | 90 | 34 | 2340 | 19 | 4 | 1589 | 740 | 34 |
| (94.8) | (3.8) | (1.4) | (99) | (0.8) | (0.2) | (67.2) | (31.3) | (1.4) | |
| Sonneratia caseolaris | 1875 | 176 | 285 | 1583 | 699 | 54 | 2315 | 17 | 4 |
| (80.3) | (7.5) | (12.2) | (67.8) | (29.9) | (2.3) | (99.1) | (0.7) | (0.2) | |
| Isolate Sequenced | Number of Isolates Obtained | Sonneratia Host | Associated Disease | GenBank Accession Number (ITS) | Fungal Taxa |
|---|---|---|---|---|---|
| P8 | 34 | S. apetala | Black leaf spot | MZ127510 | Curvulariaaff. tsudae |
| T28 | 39 | S. apetala | Stem dieback | MZ127517 | Neopestalotiopsissp.1 |
| P2 | 11 | S. caseolaris | Pink leaf spot | MZ127508 | Pestalotiopsis aff. humus |
| P4 | 45 | S. caseolaris | Pink leaf spot | MZ127507 | Pestalotiopsissp.1 |
| Y61 | 19 | S. apetala | Black leaf spot | MZ127526 | Pestalotiopsis sp.2 |
| P20 | 16 | S. caseolaris | Pink leaf spot | MZ127512 | Pestalotiopsis aff. neolitseae |
| T27 | 17 | S. caseolaris | Stem dieback | MZ127516 | Pestalotiopsis sp.3 |
| Y6 | 23 | S. apetala | Black leaf spot | MZ127521 | Pestalotiopsis sp.4b |
| Pesta23 | 23 | S. caseolaris | Black leaf spot | MZ127514 | Pestalotiopsissp.4a |
| Host Species | Isolate | Fungal Taxa | Disease Index | Pathogenicity |
|---|---|---|---|---|
| Sonneratia apetala | P8 | Curvularia aff. tsudae | 1.90 | average |
| T28 | Neopestalotiopsis sp.1 | 1.82 | average | |
| P4 | Pestalotiopsis sp.1 | 1.76 | average | |
| Pesta23 | Pestalotiopsis sp.4a | 1.55 | average | |
| Control (PDA) | 0.00 | nil | ||
| Chi-Square | 48.9 | |||
| Sonneratia caseolaris | P8 | Curvularia aff. tsudae | 1.93 | average |
| T28 | Neopestalotiopsis sp.1 | 2.36 | strong | |
| P4 | Pestalotiopsis sp.1 | 1.93 | average | |
| Pesta23 | Pestalotiopsis sp.4a | 1.89 | average | |
| Control (PDA) | 0.00 | nil | ||
| Chi-Square | 53.6 | |||
| p-Value | 0.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, H.T.T.; Hardy, G.E.S.J.; Le, T.V.; Nguyen, H.Q.; Le, D.H.; Nguyen, T.V.; Dell, B. Mangrove Dieback and Leaf Disease in Sonneratia apetala and Sonneratia caseolaris in Vietnam. Forests 2021, 12, 1273. https://doi.org/10.3390/f12091273
Nguyen HTT, Hardy GESJ, Le TV, Nguyen HQ, Le DH, Nguyen TV, Dell B. Mangrove Dieback and Leaf Disease in Sonneratia apetala and Sonneratia caseolaris in Vietnam. Forests. 2021; 12(9):1273. https://doi.org/10.3390/f12091273
Chicago/Turabian StyleNguyen, Huong Thi Thuy, Giles E. St. J. Hardy, Tuat Van Le, Huy Quoc Nguyen, Duc Hoang Le, Thinh Van Nguyen, and Bernard Dell. 2021. "Mangrove Dieback and Leaf Disease in Sonneratia apetala and Sonneratia caseolaris in Vietnam" Forests 12, no. 9: 1273. https://doi.org/10.3390/f12091273
APA StyleNguyen, H. T. T., Hardy, G. E. S. J., Le, T. V., Nguyen, H. Q., Le, D. H., Nguyen, T. V., & Dell, B. (2021). Mangrove Dieback and Leaf Disease in Sonneratia apetala and Sonneratia caseolaris in Vietnam. Forests, 12(9), 1273. https://doi.org/10.3390/f12091273