
Tissue Structure Changes of Aquilaria sinensis Xylem after Fungus Induction


Abstract
:1. Introduction
2. Materials and Methods
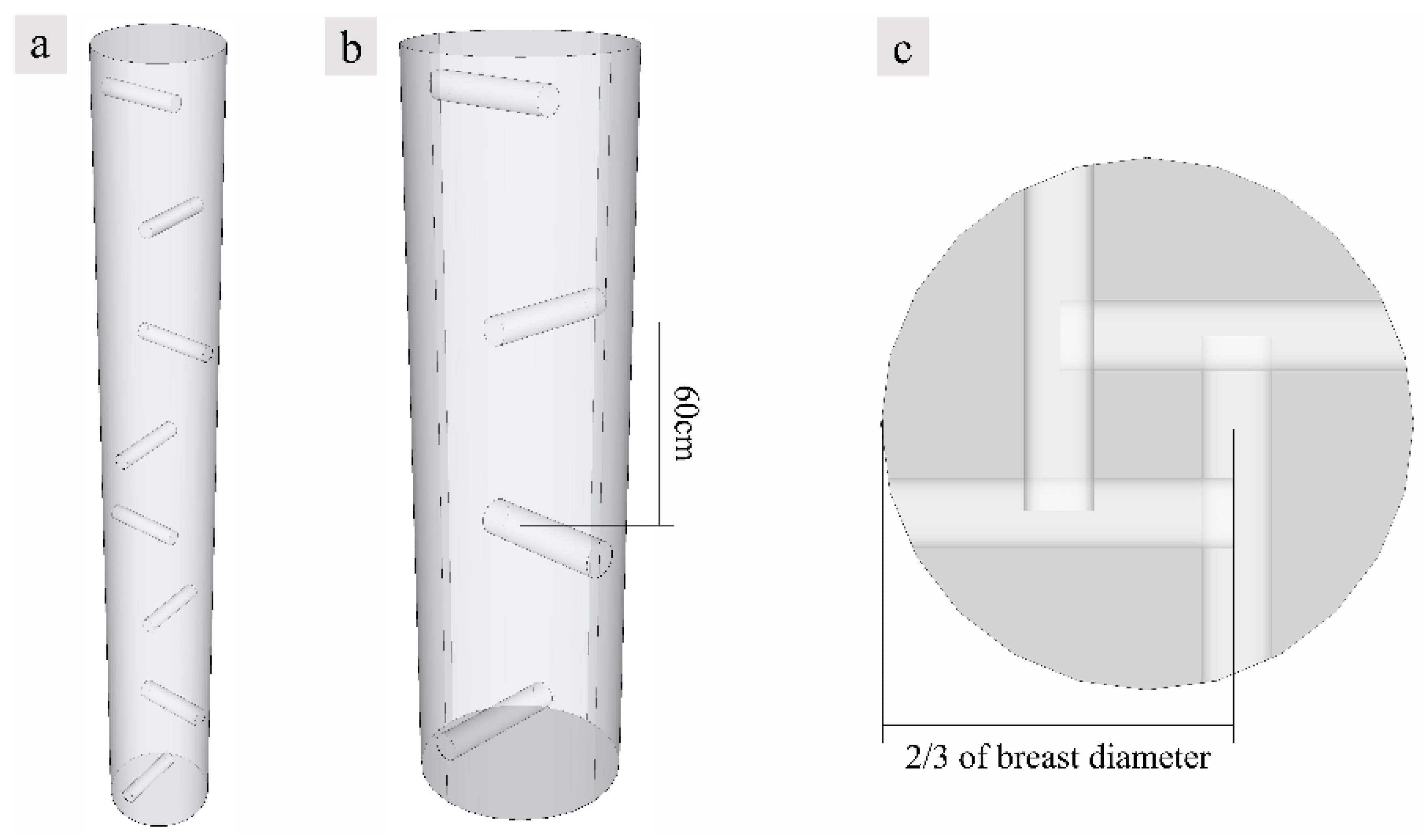
2.1. Fungus Induction Operation and Wood Collection
2.2. Observation of Xylem Tissue Structure
2.3. Scanning Electron Microscope (SEM)
2.4. Data Statistics and Analysis
3. Results
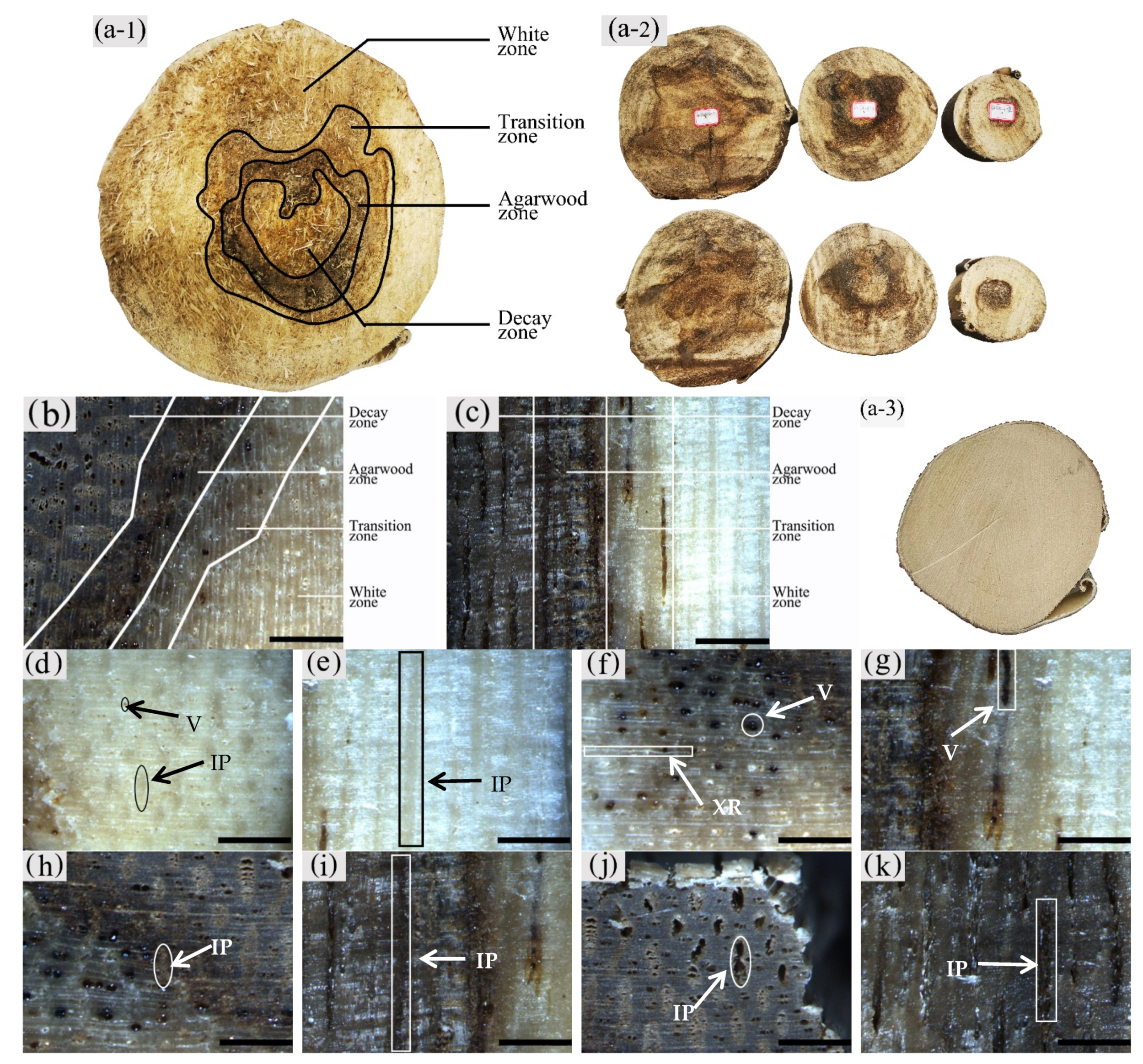
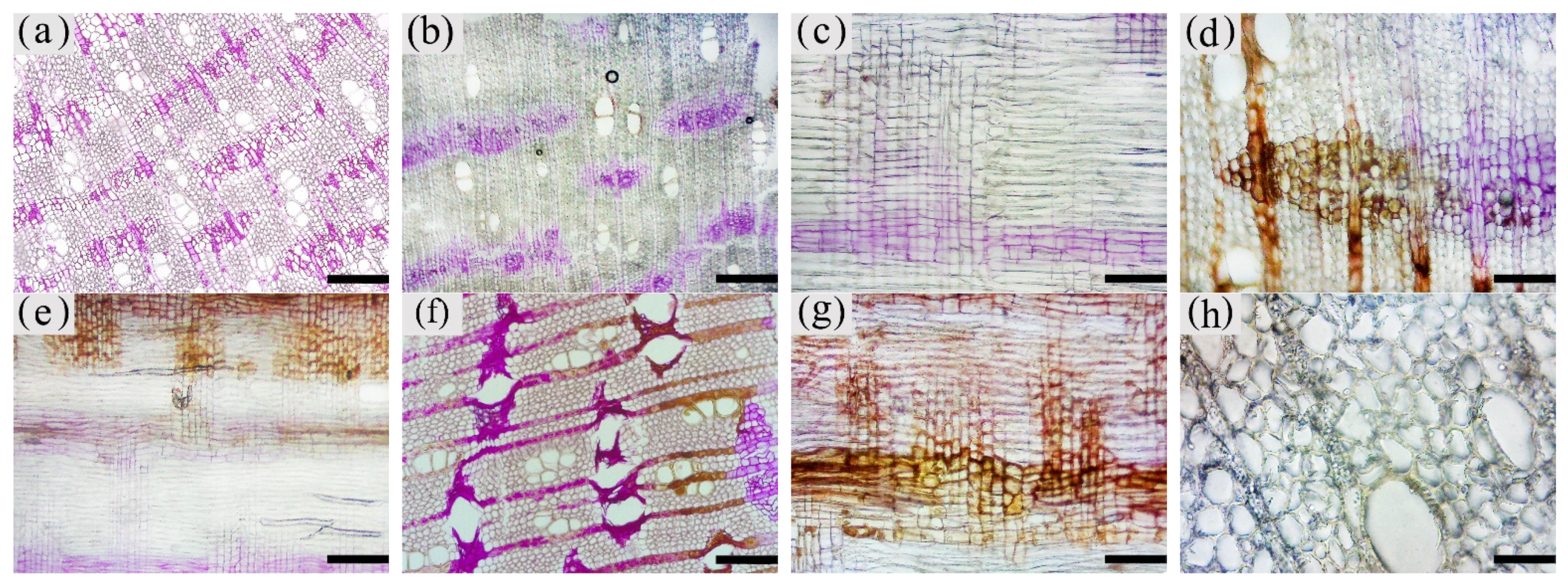
3.1. Macrostructural Differences at Different Zones of the Xylem
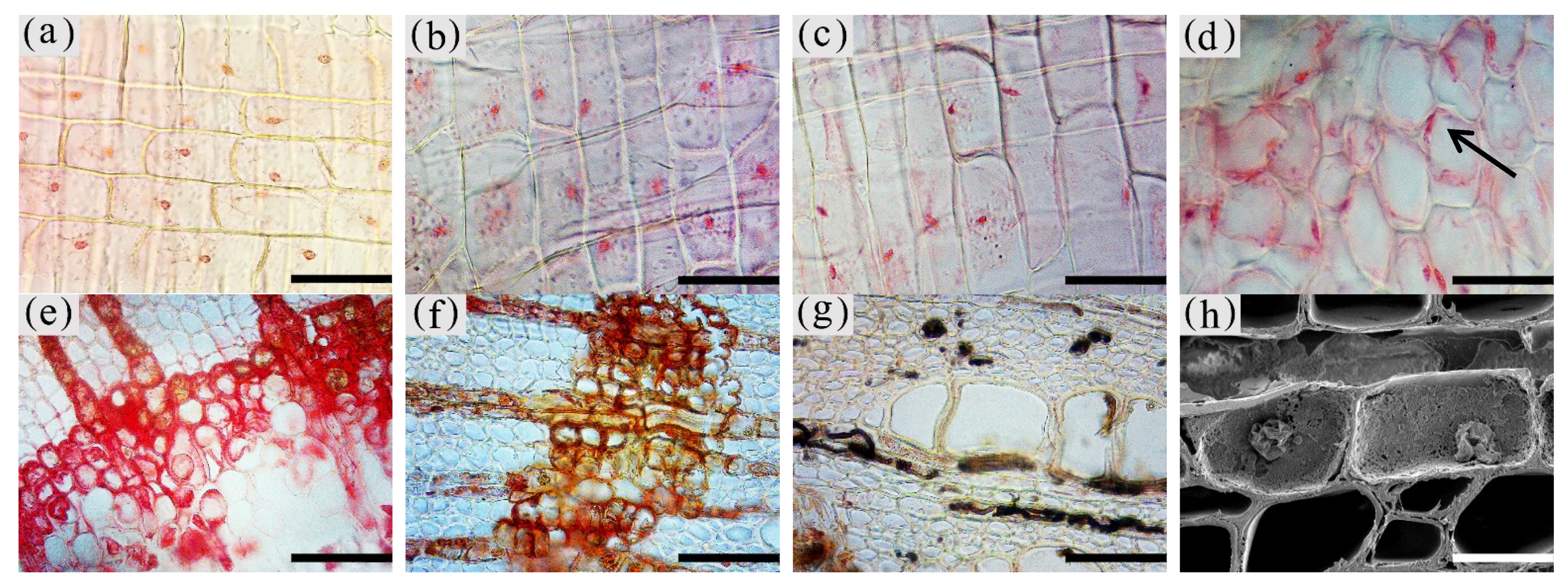
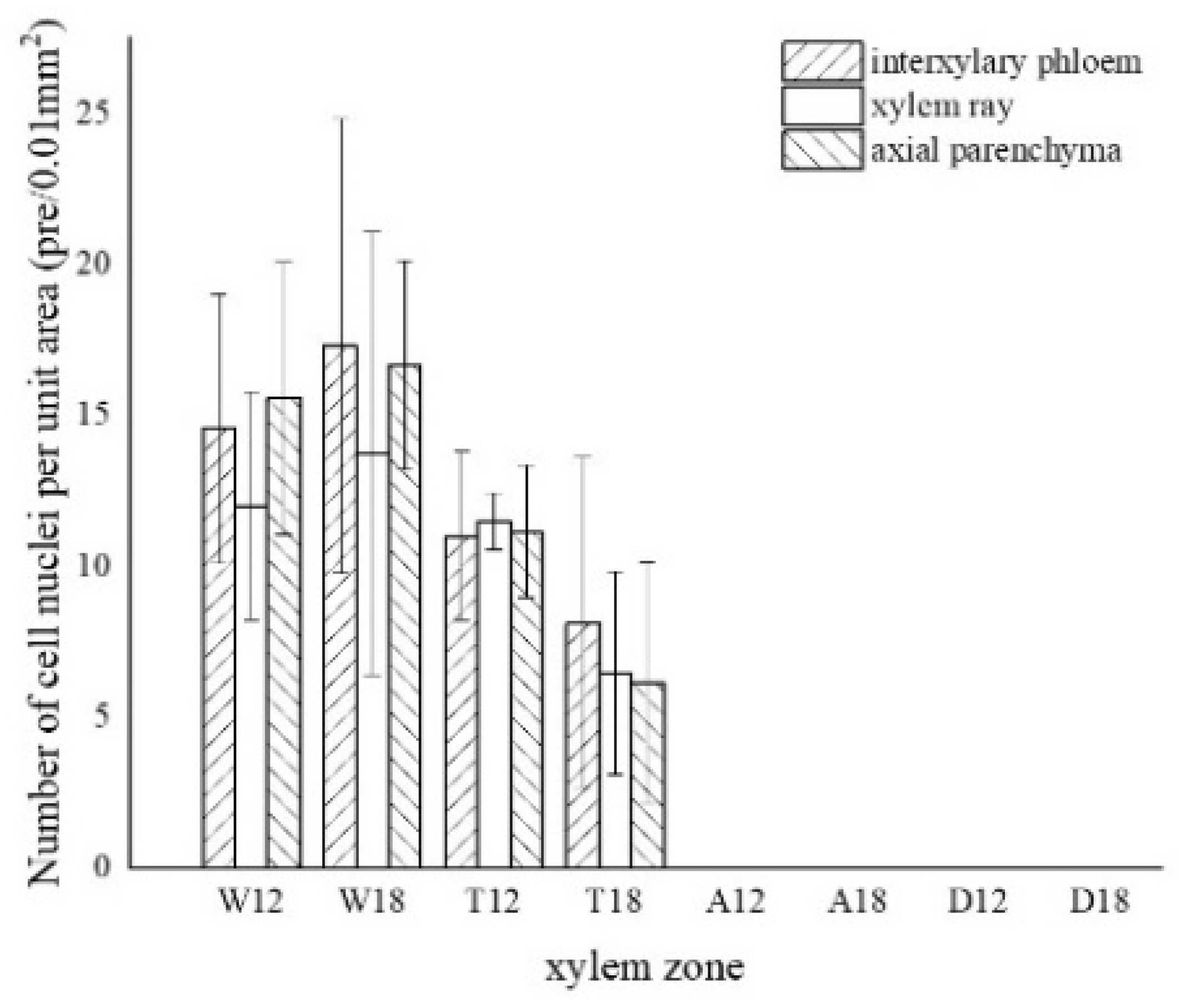
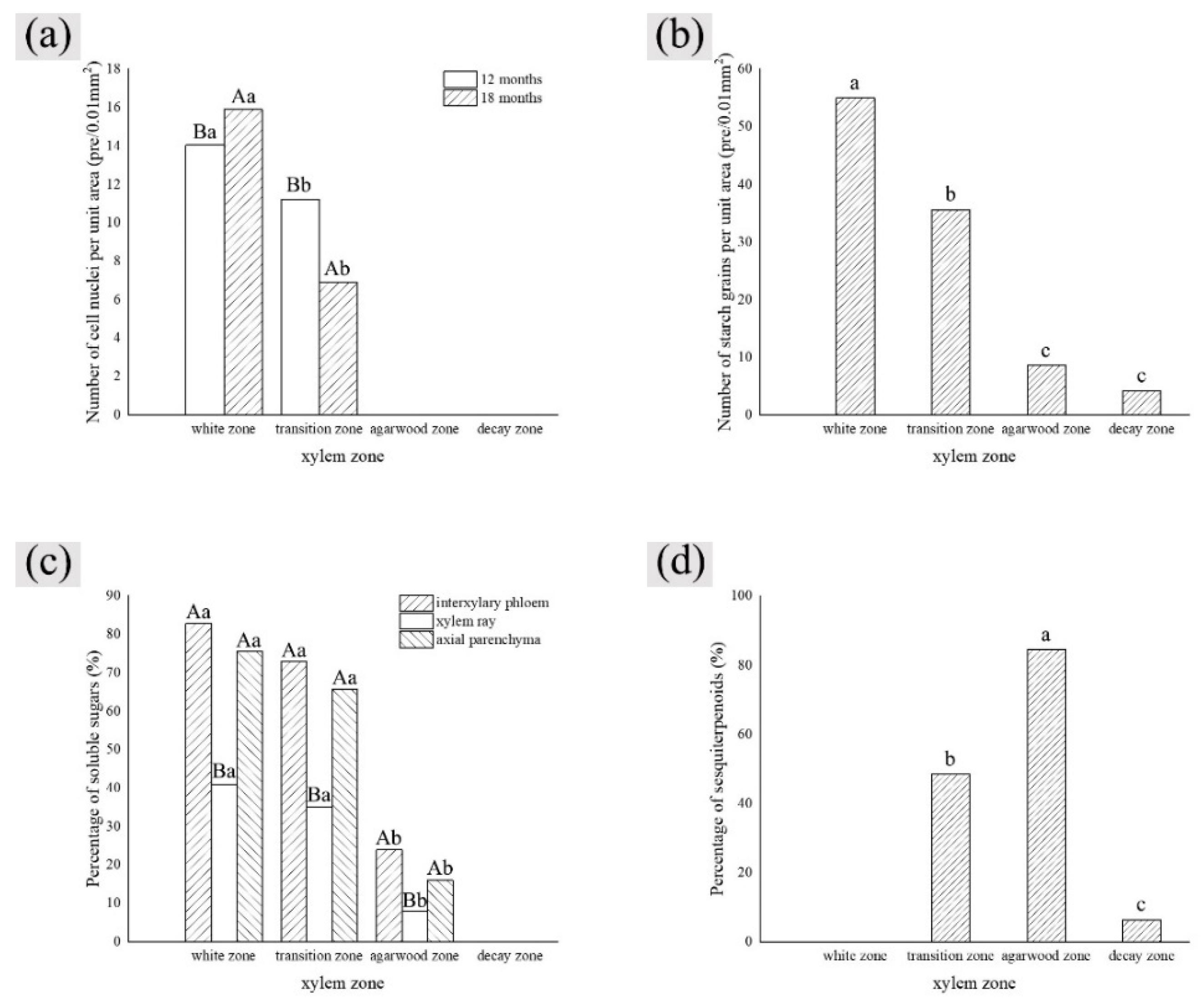
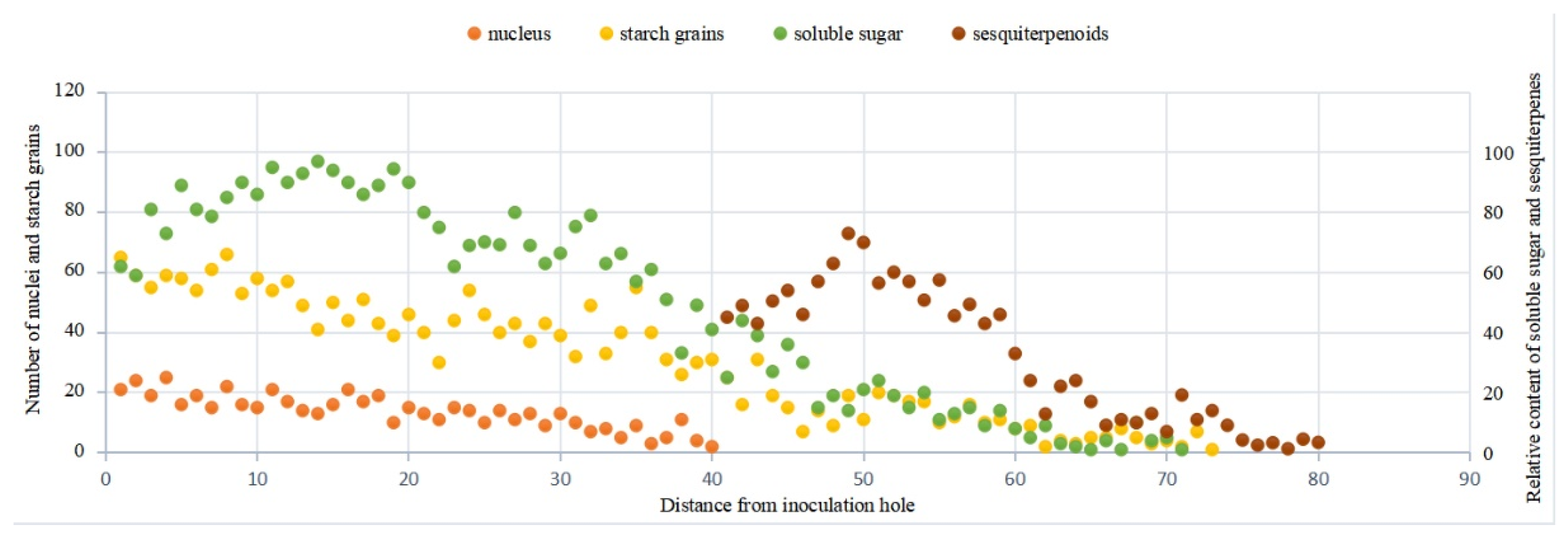
3.2. Observation on the Nucleus of Parenchyma Cells in the Xylem of A. sinensis
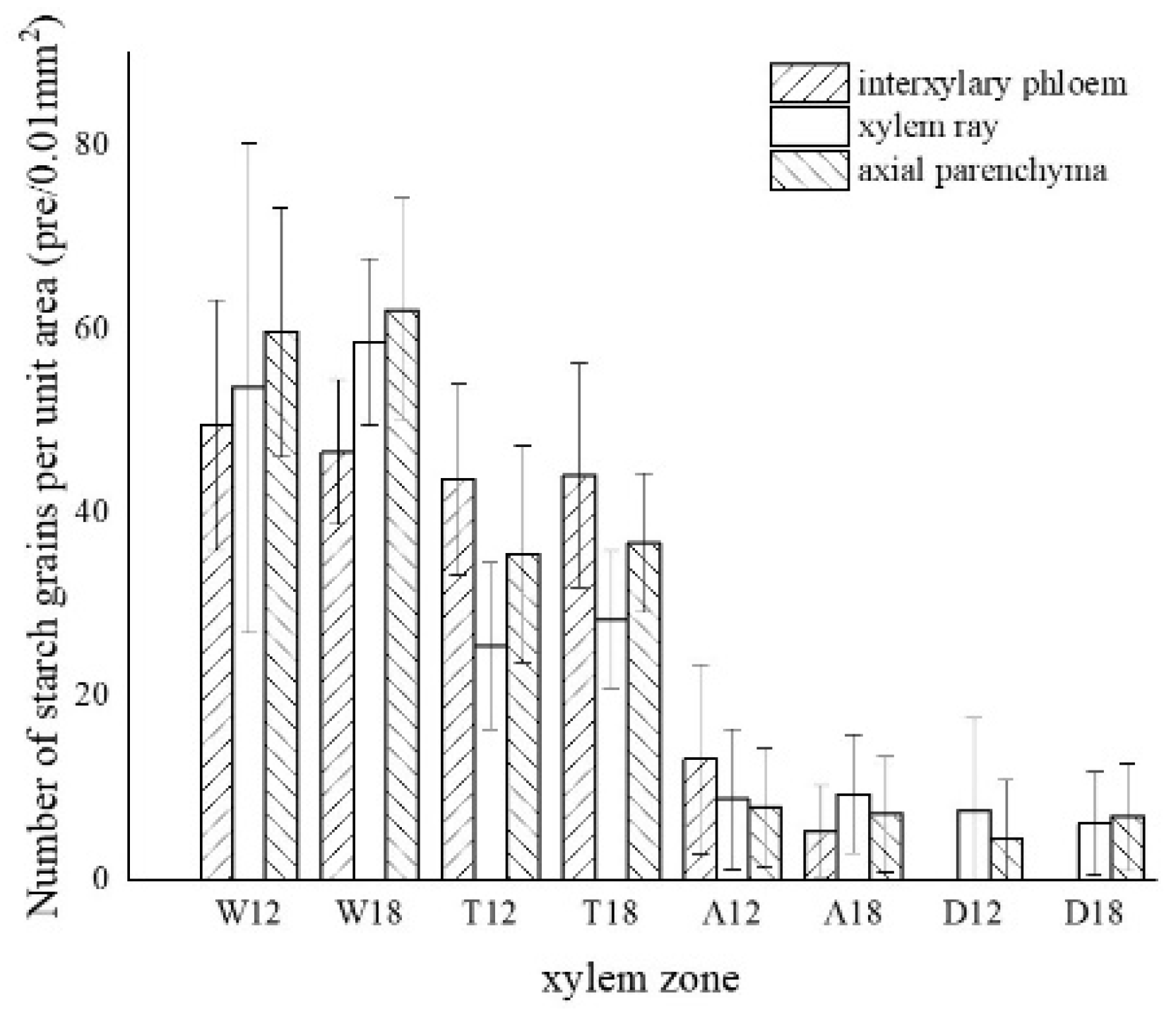
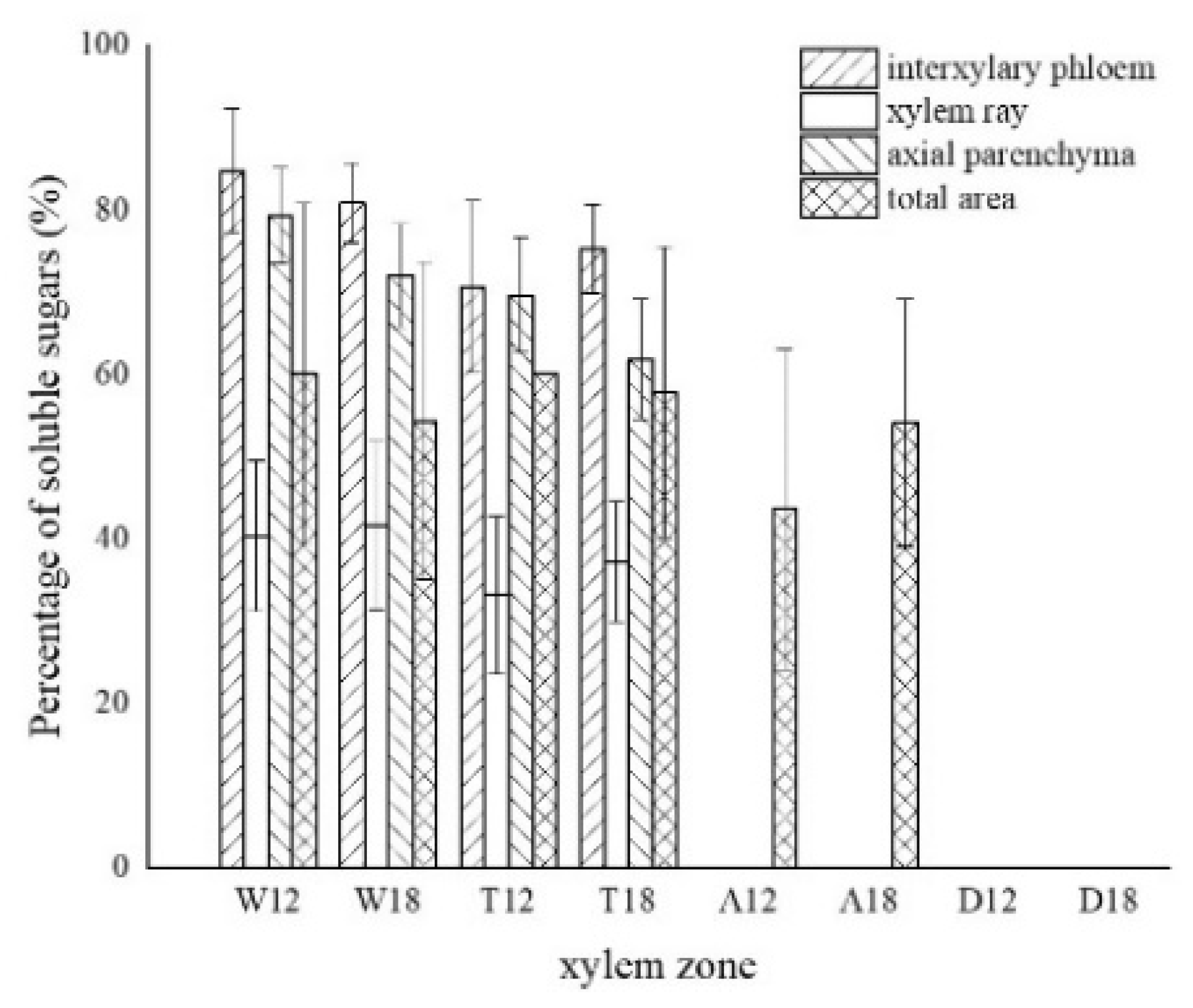
3.3. Distribution of Sugars in the Xylem of A. sinensis
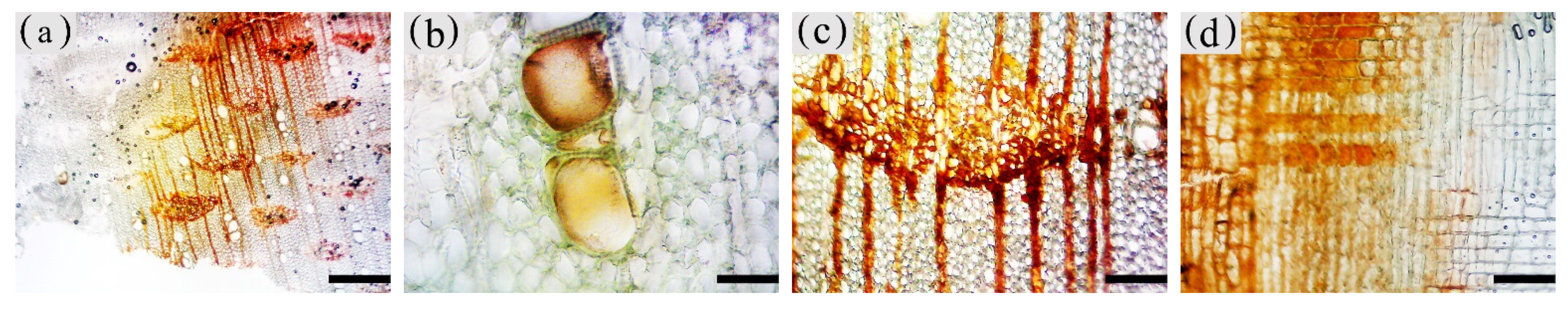
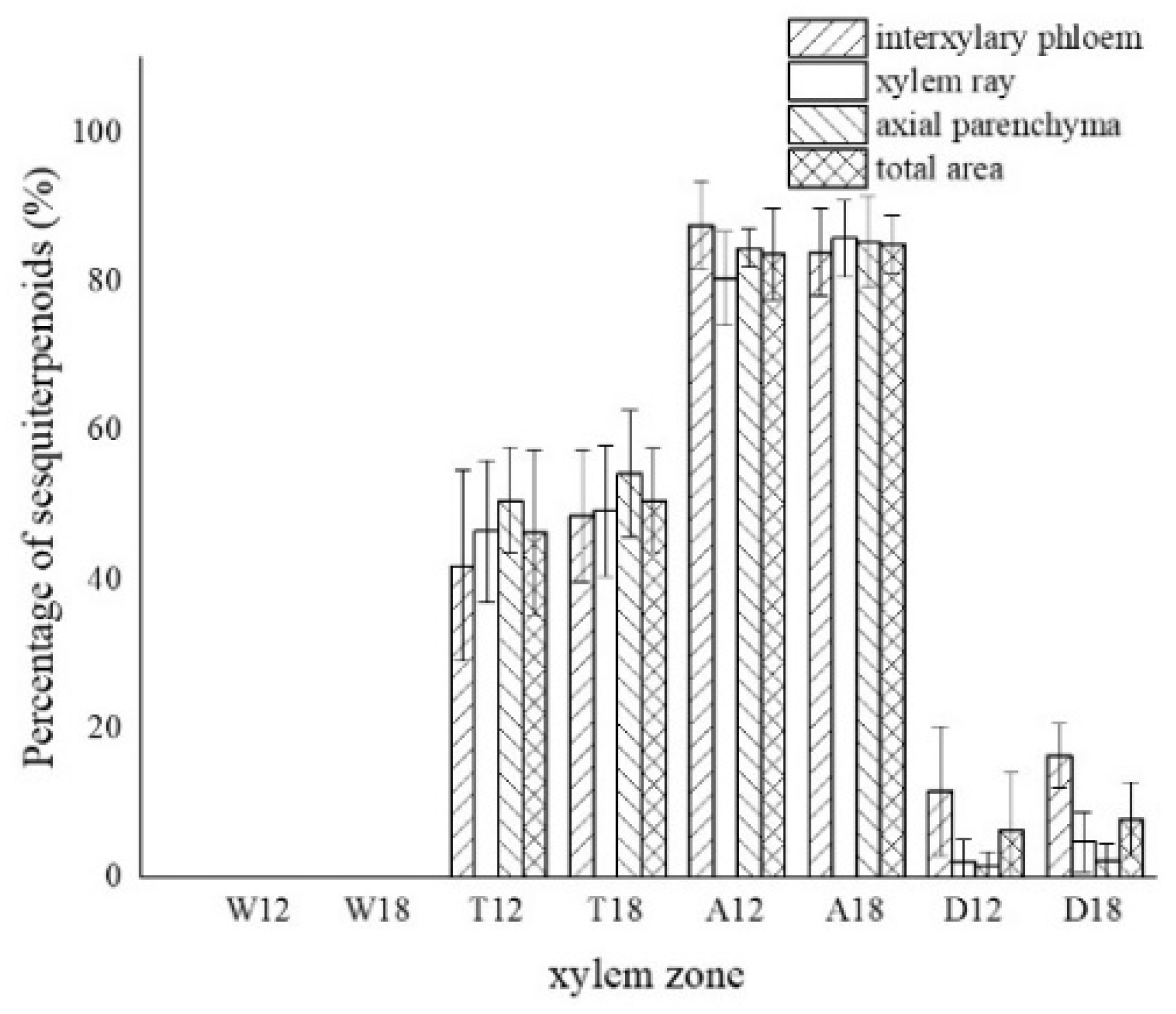
3.4. Distribution of Sesquiterpenoids in the Xylem of A. sinensis
3.5. Production of Inclusions in the Xylem of A. sinensis Parenchyma Cell
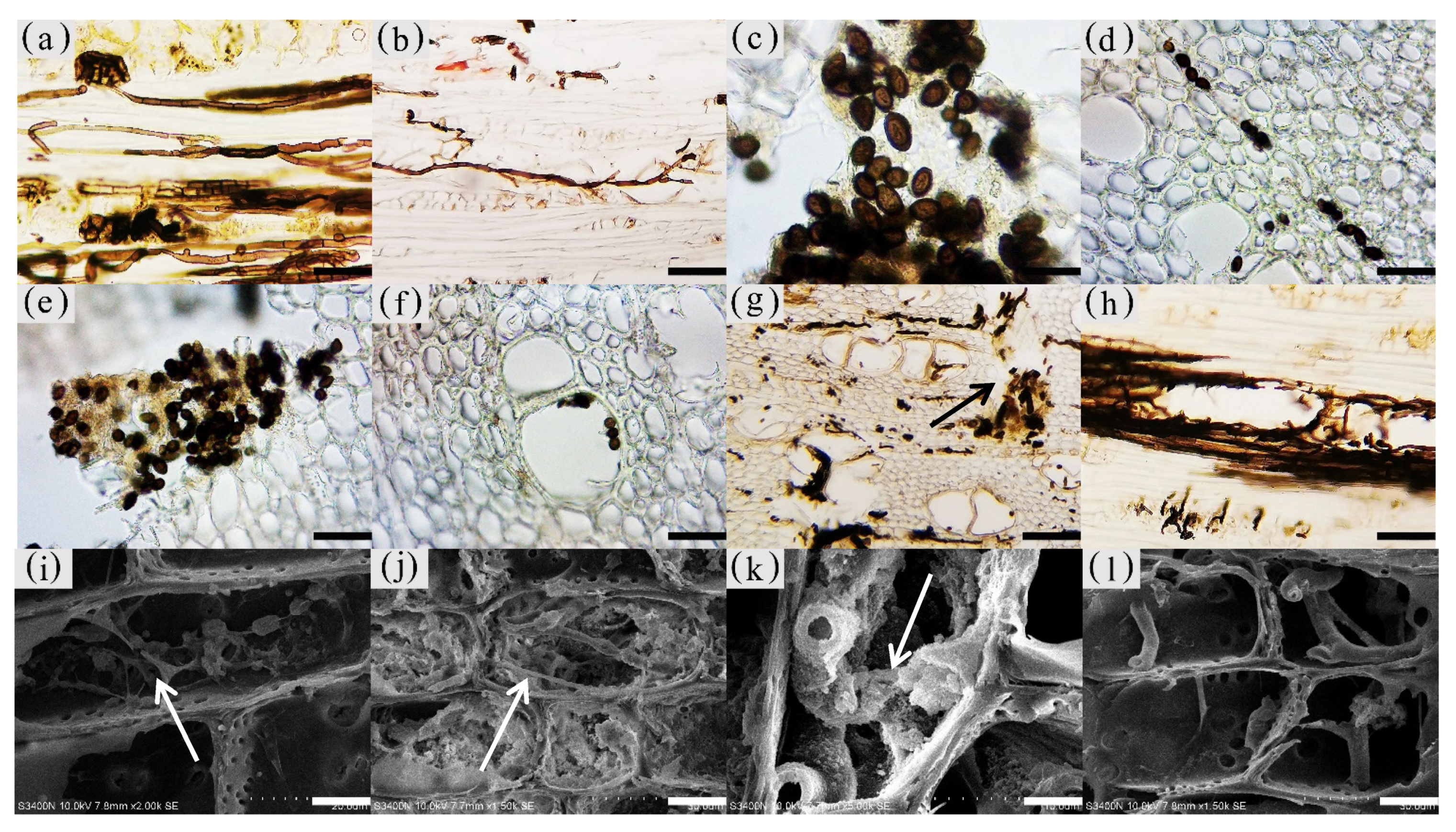
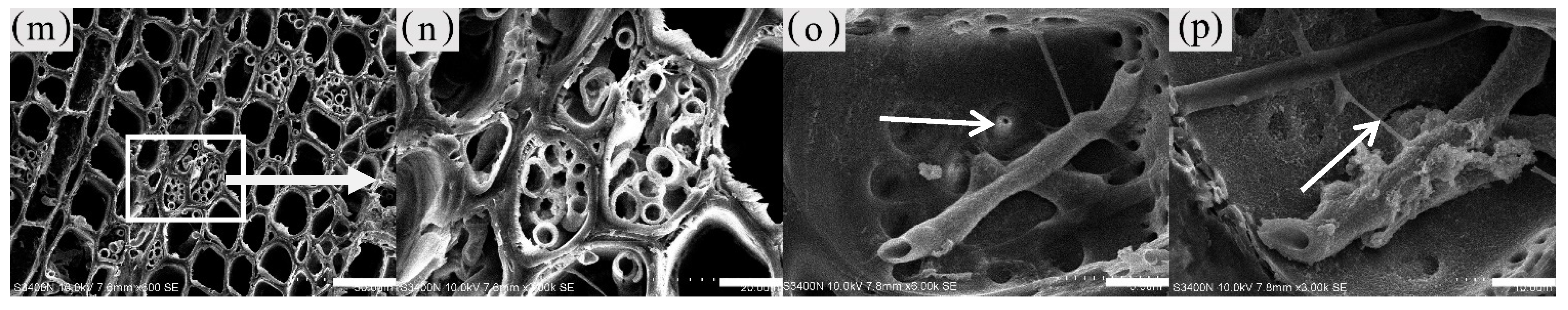
3.6. Distribution and Morphology of Fungal Hyphae and Propagules in the Xylem of A. sinensis
3.7. Variance Analysis of Factors and Xylem Structure
4. Discussion
4.1. Structural Changes in the Xylem of A. sinensis during Agarwood Formation
4.2. Relationship between Fungi and Inclusions Production
4.3. Structural Differences of Different Induction Methods
4.4. Differences in Xylem Structure at Different Inoculation Times
5. Conclusions
- The interxylary phloem was significantly related to cell extinction, sugar conversion, and generation of characteristic agarwood products during agarwood formation. Among all types of parenchyma cells, the interxylary phloem had the fastest response to the agar-wood process and was the first region of agarwood generation;
- Compared with the 12 months of inoculation in A. sinensis, the xylem of the 18 months of inoculation was more vigorous. There were no significant differences between the 12 and 18 months of inoculation in terms of sugars and agarwood characteristic products. In production, harvesting after 12 months of inoculation can improve harvesting efficiency;
- The fungus entered the A. sinensis, expanded outward along the vessels and xylem rays, aggregated in the interxylary phloem and xylem rays, and induced the production of agarwood. In agarwood-bearing trees, fungal propagules and hyphae were mainly distributed in the decay zone.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, Z.; Lu, A.; Tang, Y.; Chen, Z.; Li, D. The Families and Genera of Angiosperms in China—A Comprehensive Analysis; China Science Publishing & Media Ltd.: Beijing, China, 2003. [Google Scholar]
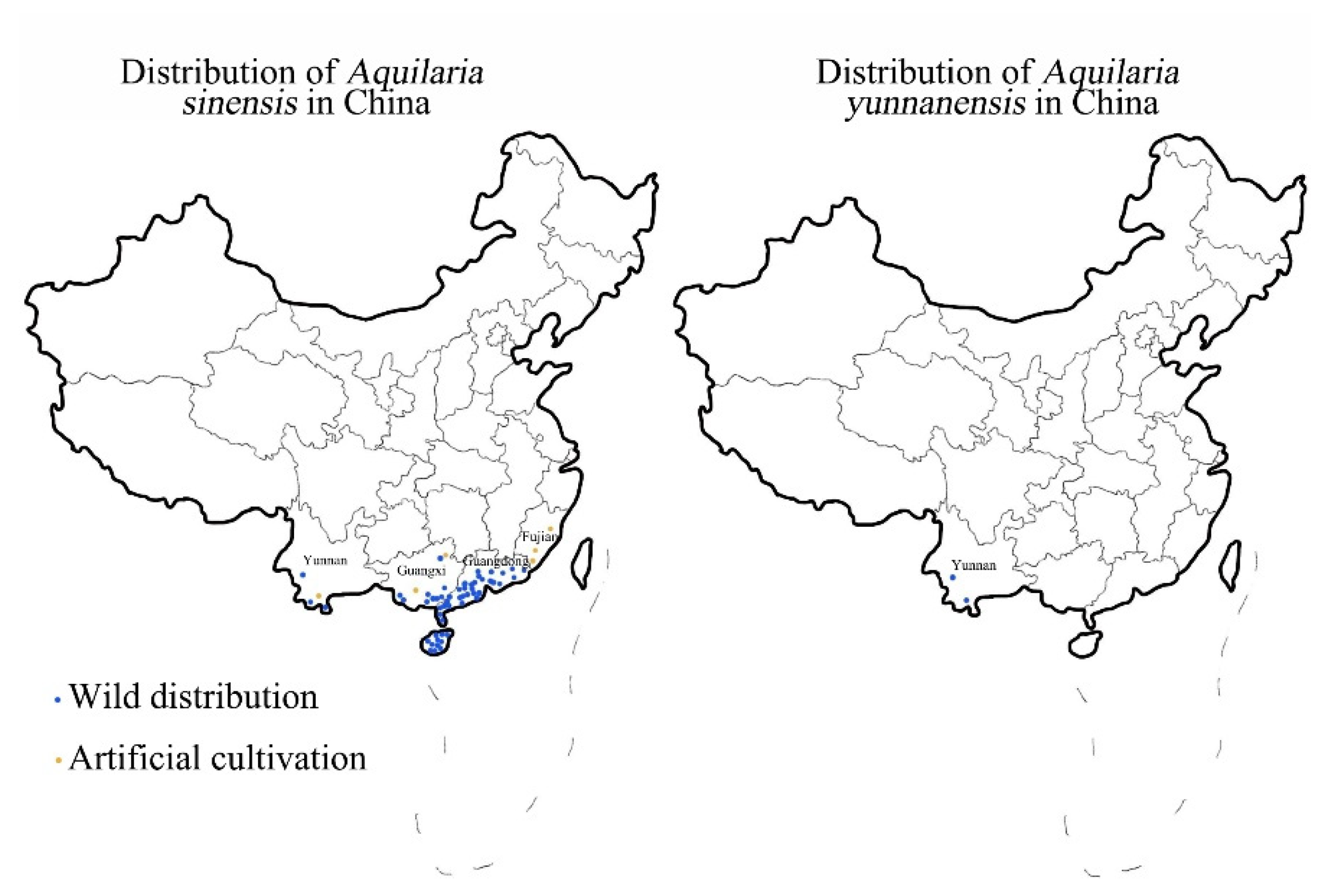
- Li, L.; Shou, H.; Ma, Q. Distribution of Aquilaria sinensis (Thymelaeaceae). J. Anhui Agri. Sci. 2012, 40, 9254–9256. [Google Scholar]
- National Key Protection of Wild Plants List (the First Batch); Communique of the State Council of the People’s Republic of China: Beijing, China, 1999; pp. 39–47.
- Convention on International Trade in Endangered Species of Wild Fauna and Flora. J. Int. Wildl. Law AMP Policy 2005, 8, 229.
- Chen, Y.; Liu, H.; Heinen, J. Challenges in the Conservation of an Over-Harvested Plant Species with High Socioeconomic Values. Sustainability 2019, 11, 4194. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chen, H.; Yang, Y.; Zhang, Z.; Wei, J.; Meng, H.; Chen, W.; Feng, J.; Gan, B.; Chen, X.; et al. Whole-tree Agarwood-Inducing Technique: An Efficient Novel Technique for Producing High-Quality Agarwood in Cultivated Aquilaria sinensis Trees. Molecules 2013, 18, 3086–3106. [Google Scholar] [CrossRef]
- Wu, Z. Studies on the Agarwood Forming Mechanism and Quality Evaluation of Aquilaria sinensis by Biologically Induced Method; Guangzhou University of Chinese Medicine: Guangzhou, China, 2015. [Google Scholar]
- Ei, W.; Zuo, W.; Yang, D.; Dong, W.; Dai, H. Advances in the Mechanism, Artificial Agarwood-induction Techniques and Chemical Constituents of Artificial Agarwood Production. Chin. J. Trop. Crops 2013, 34, 2513–2520. [Google Scholar]
- Liu, Y. Research on Quality Characteristics of Agarwood Produced by Whole-Tree Agarwood-Inducing Technique; Peking Union Medical College: Beijing, China, 2018. [Google Scholar]
- Yun, Y.; Feng, J.; Zhu, J.; Liu, Y.; Chen, D.; Wei, J. Quality of Agarwood Produced by Whole Tree Agarwood-inducing Technique (Agar-Wit) from China and Southeast Asian Countries. Chin. Pharm. J. 2019, 54, 1988–1994. [Google Scholar]
- Chen, X.; Sui, C.; Liu, Y.; Yang, Y.; Liu, P.; Zhang, Z.; Wei, J. Agarwood Formation Induced by Fermentation Liquid of Lasiodiplodia theobromae, the Dominating Fungus in Wounded Wood of Aquilaria sinensis. Curr. Microbiol. 2017, 74, 460–468. [Google Scholar] [CrossRef]
- Chen, X.; Liu, Y.; Yang, Y.; Yang, Y.; Liu, P.; Zhang, Z.; Wei, J. Trunk surface agarwood-inducing technique with Rigidoporus vinctus: An efficient novel method for agarwood production. PLoS ONE 2018, 13, e198111. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, R.; Jong, P.L.; Kamziah, A.K. Fungal inoculation induces agarwood in young Aquilaria malaccensis trees in the nursery. J. For. Res. 2014, 25, 201–204. [Google Scholar] [CrossRef]
- Faizal, A.; Esyanti, R.R.; Aulianisa, E.N.; Santoso, E.; Turjaman, M. Formation of agarwood from Aquilaria malaccensis in response to inoculation of local strains of Fusarium solani. Trees 2017, 31, 189–197. [Google Scholar] [CrossRef]
- Zhang, Z.; Han, X.; Wei, J.; Xue, J.; Yang, Y.; Liang, L.; Li, X.; Guo, Q.; Xu, Y.; Gao, Z. Compositions and Antifungal Activities of Essential Oils from Agarwood of Aquilaria sinensis (Lour.) Gilg Induced by Lasiodiplodia theobromae (Pat.) Griffon. & Maubl. J. Braz. Chem. Soc. 2014, 25, 20–26. [Google Scholar] [CrossRef]
- Liu, P.; Zhang, X.; Yang, Y.; Xue, J.; Yang, Y.; Liang, L.; Li, X.; Guo, Q.; Xu, Y.; Gao, Z. Interxylary phloem and xylem rays are the structural foundation of agarwood resin formation in the stems of Aquilaria sinensis. Trees 2019, 33, 533–542. [Google Scholar] [CrossRef]
- Tabata, Y.; Widjaja, E.; Mulyaningsih, T.; Parman, I.; Wiriadinata, H.; Mandang, Y.I.; Itoh, T. Structural survey and artificial induction of aloeswood. Wood Res. Bull. Wood Res. Inst. Kyoto Univ. 2003, 90, 11–90. [Google Scholar]
- Fahn, A. The Development of the Secondary Body in Plants with Interxylary Phloem. In Xylorama; Springer: Birkhäuser, Basel, 1985. [Google Scholar]
- Carlquist, S. Interxylary phloem: Diversity and functions. Brittonia 2013, 65, 477–495. [Google Scholar] [CrossRef]
- He, H.; Pan, Z.; Liang, Y.; Luo, P.; Qiu, J. Anatomy comparative study of Aquilaria sinensis in different agarwood induced methods. J. For. Eng. 2019, 4, 54–59. [Google Scholar]
- Li, M.; Shen, H.; He, H.; Qiu, J. Anatomic Structure and Extracted Chemical Component Content of Aquilaria sinensis. J. Southwest For. Univ. 2017, 37, 208–213. [Google Scholar]
- Wang, J.; Wang, Y.; Jinling, Y.; Wei, L.I.; Dong, W.; Wenli, M.; Haofu, D. Comparison of the Anatomy Structure and Chemical Compositions of Agarwoods from Two Kinds of Aquilaria sinensis. SCIENTIA SILVAE SINICAE 2019, 55, 146–154. [Google Scholar]
- Keisuke, O.; Hideo, K.; Hiroo, F. Direct Evidence of Active and Rapid Nuclear Degradation Triggered by Vacuole Rupture during Programmed Cell Death in Zinnia. Plant Physiol. 2001, 125, 615–626. [Google Scholar]
- Janssen, T.A.J.; Hölttä, T.; Fleischer, K.; Naudts, K.; Dolman, H. Wood allocation trade-offs between fiber wall, fiber lumen, and axial parenchyma drive drought resistance in neotropical trees. Plant Cell Environ. 2020, 43, 965–980. [Google Scholar] [CrossRef] [Green Version]
- Sheng, M.; Yunlin, F.; Yingjian, L.; Penglian, W.; Zhigao, L. The formation and quality evaluation of agarwood induced by the fungi in Aquilaria sinensis. Ind. Crops Prod. 2021, 173, 114129. [Google Scholar]
- Ma, S.; Qiao, M.; Fu, Y.; Wei, P.; Li, Y.; Liu, Z. Comparative Analysis of Biological Activity of Artificial and Wild Agarwood. Forests 2021, 12, 1532. [Google Scholar] [CrossRef]
- Kang, Z. Ultrastructure of Plant Pathogenic Fungi; China Science & Technology Press: Beijing, China, 1995. [Google Scholar]
- Sbrana, C.; Fortuna, P.; Giovannetti, M. Plugging into the network: Belowground connections between germlings and extraradical mycelium of arbuscular mycorrhizal fungi. Mycologia 2011, 103, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Z.; Jia-Qi, G.; Yuan, Y.; Quan, Y.; Liang-Yun, Z.; Juan, L.; Lu-Qi, H. Analysis of bacteria distribution characteristics in different layers of agarwood based on Hiseq sequencing. China J. Chin. Mater. Med. 2020, 45, 2374–2381. [Google Scholar]
- Kuo, T.; Huang, H.; Hsu, C. Mass spectrometry imaging guided molecular networking to expedite discovery and structural analysis of agarwood natural products. Anal. Chim. Acta 2019, 1080, 95–103. [Google Scholar] [CrossRef]
- Zhidong, L.; Shuang, Z.; Dailin, L.; Xiaoxia, G.; Yanmin, Z.; Yu, C. Evaluation of three different artificial agarwood-inducing methods from Aquilaria sinensis using antimicrobial activity. Pak. J. Pharm. Sci. 2019, 32, 905–910. [Google Scholar]
- Nussinovitch, A. Plant Gum Exudates of the World:Sources, Distribution, Properties, and Applications; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Latt, C.R.; Nair, P.K.R.; Kang, B.T. Reserve carbohydrate levels in the boles and structural roots of five multipurpose tree species in a seasonally dry tropical climate. For. Ecol. Manag. 2001, 146, 145–158. [Google Scholar] [CrossRef]
- Zhang, X. Studies on Relationship between Wound-Induced Defense Response and Agarwood Formation in Aquilaria sinensis. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2013. [Google Scholar]
- Justin, S.; Lihan, S.; ElvisSulang, M.; Chiew, T. Formulated microbial consortium as inoculant for agarwood induction. J. Trop. For. Sci. 2020, 32, 161–169. [Google Scholar] [CrossRef]
- Gu, L.; Zheng, K.; Liu, Y.; Ma, H.; Xiao, Z. Two Kinds of Fungi Effect on the Tissue Structure of Aquilaria sinensis Xylem. J. West China For. Sci. 2018, 47, 141–144. [Google Scholar]
- Juan, L.; Xiang, Z.; Jian, Y.; Junhui, Z.; Yuan, Y.; Chao, J.; Xiulian, C.; Luqi, H. Agarwood wound locations provide insight into the association between fungal diversity and volatile compounds in Aquilaria sinensis. R. Soc. Open Sci. 2019, 6, 190211. [Google Scholar]
- Zhang, M. Apreliminary Study on the of Lsolation and Identification of Endophytic Bacteria in White Fragrant and Distribution. Master’s Thesis, Guangzhou University of Chinese Medicine, Guangzhou, China, 2017. [Google Scholar]
- Song, X.; Huang, G.; Wang, X.; Song, J.; Zhang, Q.; Liang, K.; Zhou, Z. Resistance Capability of Aquilaria sinensis under Combinations of Inorganic Salts, Hormones and Fungus Induction. Bull. Bot. Res. 2019, 39, 505–513. [Google Scholar]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Xylem Zone | Parenchyma Cells Type | Inoculation Times |
|---|---|---|---|
| number of nuclei | 0.000 ** | 0.403 | 0.048 * |
| number of starch grains | 0.000 ** | 0.592 | 0.947 |
| soluble sugar color area | 0.000 ** | 0.005 ** | 0.933 |
| sesquiterpene color area | 0.000 ** | 0.429 | 0.385 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Qiao, M.; Fu, Y.; Wei, P.; Li, Y.; Liu, Z. Tissue Structure Changes of Aquilaria sinensis Xylem after Fungus Induction. Forests 2022, 13, 43. https://doi.org/10.3390/f13010043
Liu Y, Qiao M, Fu Y, Wei P, Li Y, Liu Z. Tissue Structure Changes of Aquilaria sinensis Xylem after Fungus Induction. Forests. 2022; 13(1):43. https://doi.org/10.3390/f13010043
Chicago/Turabian StyleLiu, Yuxin, Mengji Qiao, Yunlin Fu, Penglian Wei, Yingjian Li, and Zhigao Liu. 2022. "Tissue Structure Changes of Aquilaria sinensis Xylem after Fungus Induction" Forests 13, no. 1: 43. https://doi.org/10.3390/f13010043
APA StyleLiu, Y., Qiao, M., Fu, Y., Wei, P., Li, Y., & Liu, Z. (2022). Tissue Structure Changes of Aquilaria sinensis Xylem after Fungus Induction. Forests, 13(1), 43. https://doi.org/10.3390/f13010043