Utilization of the Hollies (Ilex L. spp.): A Review



Abstract
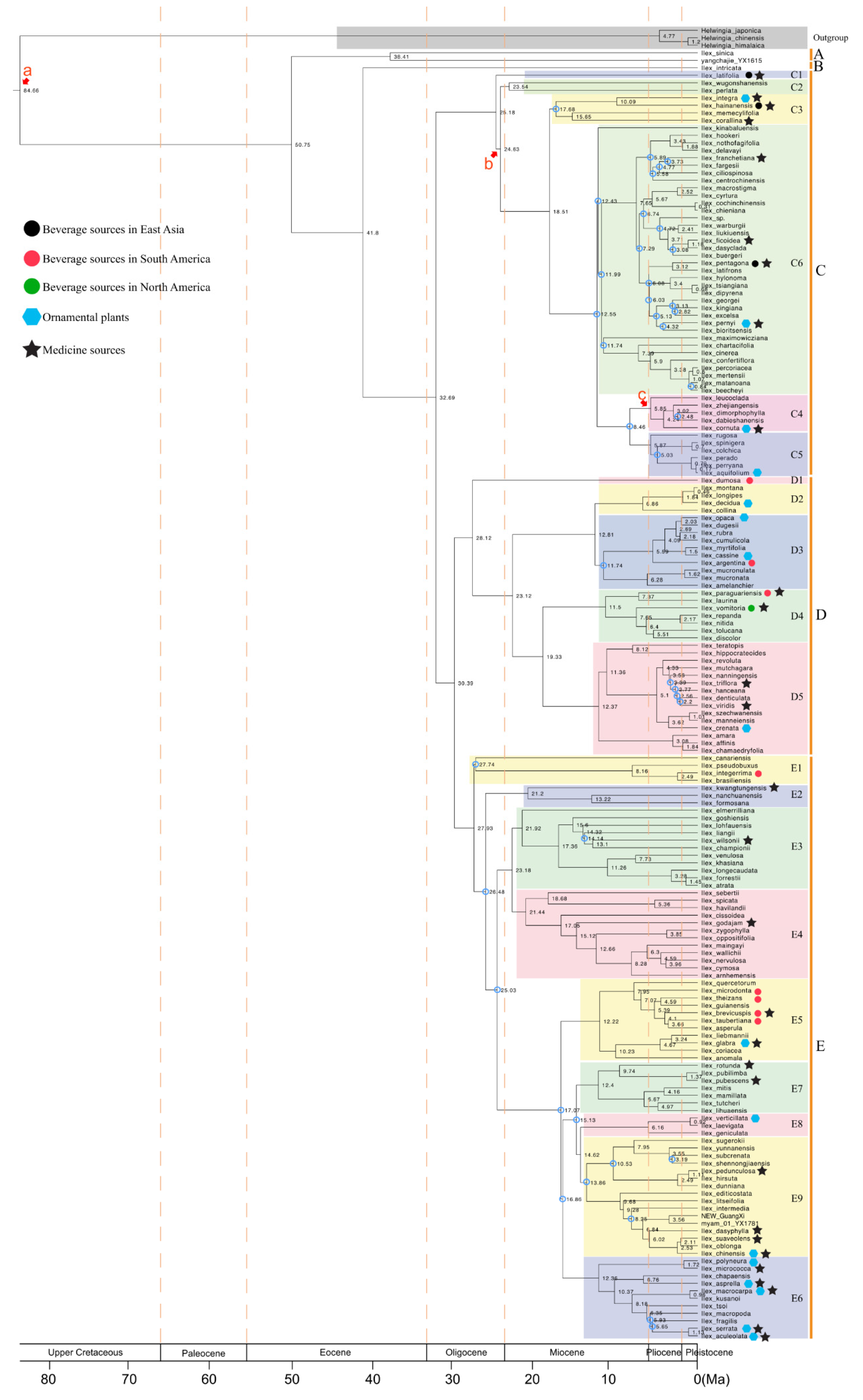
:1. Introduction
2. Hollies Utilized as Sources of Beverages
3. Hollies Utilized as Sources of Medicines
4. Hollies Utilized as Ornamental Plants
5. Hollies Utilized as Honey Plants
6. Other Uses
6.1. Timber
6.2. Minor Uses
7. Future Prospects for Utilization of Ilex Species
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- POWO. Plants of the World Online. Available online: http://www.plantsoftheworldonline.org/ (accessed on 4 November 2021).
- Yao, X.; Song, Y.; Yang, J.B.; Tan, Y.H.; Corlett, R.T. Phylogeny and biogeography of the hollies (Ilex L., Aquifoliaceae). J. Syst. Evol. 2021, 59, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Yao, X.; Lu, Z.; Song, Y.; Hu, X.; Corlett, R.T. A chromosome-scale genome assembly for the holly (Ilex polyneura) provides insights into genomic adaptations to elevation in Southwest China. Hortic. Res. 2021. accepted. [Google Scholar]
- Li, L.; Xu, L.J.; Ma, G.Z.; Dong, Y.M.; Peng, Y.; Xiao, P.G. The large-leaved kudingcha (Ilex latifolia Thunb and Ilex kudingcha CJ Tseng): A traditional Chinese tea with plentiful secondary metabolites and potential biological activities. J. Nat. Med. 2013, 67, 425–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Li, X.; Peng, P.; Peng, J.; Wang, L. Kudingcha resources and rural kudingcha in Sichuan. Fujianchaye 2004, 1, 9–10. [Google Scholar]
- Zhang, D.; Yan, D. Development prospect and domestication cultivation of kudingcha (Ilex pentagona) in mountainous areas of Guizhou province. J. Guizhou Agric. Sci. 2004, 32, 85–87. [Google Scholar]
- Gu, J.; Peng, Y.; Xu, L.; Xiao, W.; Chen, Z.; Xiao, P. Sources investigation and characters identification of kudingcha production. J. Chin. Med. Mater. 2011, 34, 196–199. [Google Scholar]
- Liu, B.; Ma, R.; Zhang, J.; Sun, P.; Yi, R.; Zhao, X. Preventive effect of small-leaved kuding tea (Ligustrum robustum (Roxb.) Bl.) polyphenols on D-galactose-induced oxidative stress and aging in mice. Evid.-Based Complement. Altern. Med. 2019, 2019, 3152324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Zhou, L.; Qiao, Y.; Qi, J.; Fu, G.; Xia, X. Quality evaluation of Ilex asprella based on simultaneous determination of five bioactive components, chlorogenic acid, luteoloside, quercitrin, quercetin, and kaempferol, using UPLC-Q-TOF MS study. J. AOAC Int. 2019, 102, 1414–1422. [Google Scholar] [CrossRef]
- Jiang, S.; Cui, H.; Wu, P.; Liu, Z.; Zhao, Z. Botany, traditional uses, phytochemistry, pharmacology and toxicology of Ilex pubescens Hook et Arn. J. Ethnopharmacol. 2019, 245, 112147. [Google Scholar] [CrossRef]
- Negrin, A.; Long, C.; Motley, T.J.; Kennelly, E.J. LC-MS metabolomics and chemotaxonomy of caffeine-containing holly (Ilex) species and related taxa in the Aquifoliaceae. J. Agric. Food Chem. 2019, 67, 5687–5699. [Google Scholar] [CrossRef]
- Filip, R.; Lopez, P.; Coussio, J.; Ferraro, G. Mate substitutes or adulterants: Study of xanthine content. Phytother. Res. 1998, 12, 129–131. [Google Scholar] [CrossRef]
- Reginatto, F.H.; Athayde, M.L.; Gosmann, G.; Schenkel, E.P. Methylxanthines accumulation in Ilex species-caffeine and theobromine in erva-mate (Ilex paraguariensis) and other Ilex species. J. Braz. Chem. Soc. 1999, 10, 443–446. [Google Scholar] [CrossRef] [Green Version]
- Seyed, M.A.R. Determinación de Arbutina en las Especies de Ilex. Ph.D. Thesis, Universidad de Belgrano, Buenos Aires, Argentina, 2012. [Google Scholar]
- Mesquita, M.; Santos, E.; Kassuya, C.A.; Salvador, M.J. Chimarrão, terere and mate-tea in legitimate technology modes of preparation and consume: A comparative study of chemical composition, antioxidant, anti-inflammatory and anti-anxiety properties of the mostly consumed beverages of Ilex paraguariensis St. Hil. J. Ethnopharmacol. 2021, 279, 114401. [Google Scholar] [PubMed]
- Gottlieb, A.M.; Poggio, L. Quantitative and qualitative genomic characterization of cultivated Ilex L. species. Plant Genet. Resour. 2015, 13, 142–152. [Google Scholar] [CrossRef]
- Maiocchi, M.G.; Gorrales, L.; Cardoso-Schiavi, P.; Serrano, N.; Petenatti, E.M.; Marchevsky, E.J.; del Vitto, L.A. Nutritional parameters of “mate cocido” with milk prepared from two species of Ilex and their commercial mixture for school age population. Rev. Fac. Cienc. Agrar. 2018, 50, 203–216. [Google Scholar]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#search/Mat%C3%A9 (accessed on 4 November 2021).
- Croge, C.P.; Cuquel, F.L.; Pintro, P.T.M. Yerba mate: Cultivation systems, processing and chemical composition. A review. Sci. Agric. 2021, 78, e20190259. [Google Scholar] [CrossRef]
- Magri, E.; Gugelmin, E.K.; Grabarski, F.A.P.; Barbosa, J.Z.; Auler, A.C.; Wendling, I.; Prior, S.A.; Valduga, A.T.; Motta, A.C.V. Manganese hyperaccumulation capacity of Ilex paraguariensis A. St. Hil. and occurrence of interveinal chlorosis induced by transient toxicity. Ecotoxicology 2020, 203, 111010. [Google Scholar] [CrossRef] [PubMed]
- Paiva, D.I.; Cascales, J.; Rosetti, M.E.N.; Scherer, R.A.; Gauchat, M.E.; Gottlieb, A. Unraveling the genetic complexity of a cultivated breeding population of “yerba mate”(Ilex paraguariensis St. Hil.). An. Acad. Bras. Cienc. 2020, 92, e20190113. [Google Scholar] [CrossRef]
- Dueñas, J.F.; Jarrett, C.; Cummins, I.; Logan-Hines, E. Amazonian guayusa (Ilex guayusa Loes.): A historical and ethnobotanical overview. Econ. Bot. 2016, 70, 85–91. [Google Scholar] [CrossRef]
- Erazo-Garcia, M.P.; Guadalupe, J.J.; Rowntree, J.K.; Borja-Serrano, P.; de los Monteros-Silva, N.E.; Torres, M.D.L. Assessing the genetic diversity of Ilex guayusa Loes., a medicinal plant from the Ecuadorian Amazon. Diversity 2021, 13, 182. [Google Scholar] [CrossRef]
- Wise, G.; Negrin, A. A critical review of the composition and history of safe use of guayusa: A stimulant and antioxidant novel food. Crit. Rev. Food Sci. Nutr. 2020, 60, 2393–2404. [Google Scholar] [CrossRef] [PubMed]
- Wainwright, A.E.; Putz, F.E. A misleading name reduces marketability of a healthful and stimulating natural product: A comparative taste test of infusions of a native Florida holly (Ilex vomitoria) and yerba mate (I. paraguariensis). Econ. Bot. 2014, 68, 350–354. [Google Scholar] [CrossRef]
- Folch, C. Ceremony, medicine, caffeinated Tea: Unearthing the forgotten faces of the North American stimulant yaupon (Ilex vomitoria). Comp. Stud. Soc. Hist. 2021, 63, 464–498. [Google Scholar] [CrossRef]
- Yanovsky, E. Food Plants of the North American Indians; United States Department of Agriculture Miscellaneous Publications No. 237; United States Department of Agriculture: Washington, DC, USA, 1936.
- Grieve, M. A Modern Herbal; Harcourt, Brace & Company: New York, NY, USA, 1931. [Google Scholar]
- Yi, F.; Zhao, X.; Peng, Y.; Xiao, P. Genus Ilex L.: Phytochemistry, ethnopharmacology, and pharmacology. Chin. Herb. Med. 2016, 8, 209–230. [Google Scholar] [CrossRef]
- Noureddine, T.; El Husseini, Z.; Nehme, A.; Massih, R.A. Antibacterial activity of Ilex paraguariensis (Yerba Mate) against Gram-positive and Gram-negative bacteria. J. Infect. Dev. Ctries. 2018, 12, 712–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallmann, A.P.; dos Santos, C.V.; da Silva, T.L.; Bandeira, D.M.; Lakoski, L.V.; Rosset, J.; Conceição, L.H.S.M.; Pinto, F.G.S. Yield, phytochemical characterization and evaluation of antibacterial and antioxidante activity of Ilex brevicuspis (Aquifoliaceae) Reissek extracts against Salmonella spp. serotypes of poultry origen. Braz. J. Dev. 2021, 7, 29143–29158. [Google Scholar] [CrossRef]
- Mallmann, A.P.; dos Santos, C.V.; Toledo, A.G.; Lakoski, L.V.; Bandeira, D.M.; Corrêa, J.M.; Pinto, F.G.S. Determination of the phytochemical profile and evaluation of biological activities of leaf extracts of Ilex brevicuspis Reissek. (Aquifoliaceae). Res. Soc. Dev. 2021, 10, e10210313154. [Google Scholar] [CrossRef]
- García-Ruiz, A.; Baenas, N.; Benítez-González, A.M.; Stinco, C.M.; Meléndez-Martínez, A.J.; Moreno, D.A.; Ruales, J. Guayusa (Ilex guayusa L.) new tea: Phenolic and carotenoid composition and antioxidant capacity. J. Sci. Food Agric. 2017, 97, 3929–3936. [Google Scholar] [CrossRef] [PubMed]
- Chóez-Guaranda, I.; Viteri-Espinoza, R.; Barragán-Lucas, A.; Quijano-Avilés, M.; Manzano, P. Effect of solvent-solvent partition on antioxidant activity and GC-MS profile of Ilex guayusa Loes. leaves extract and fractions. Nat. Prod. Res. 2021. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Yang, H.; Sun, W.D.; Wang, J.; Zhang, B.Y.; Shen, Y.J.; Yin, M.Q.; Liu, Y.X.; Liu, C.; Sun, Y. Ethanol extract of Ilex hainanensis Merr. exhibits anti-melanoma activity by induction of G 1/S cell-cycle arrest and apoptosis. Chin. J. Integr. Med. 2018, 24, 47–55. [Google Scholar] [CrossRef]
- Zhuang, H.; Wang, C.; Wang, Y.; Jin, T.; Huang, R.; Lin, Z.; Wang, Y. Native useful vascular plants of China: A checklist and use patterns. Plant Divers. 2021, 43, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Shao, S.Y.; Li, R.F.; Sun, H.; Li, S. New triterpenoid saponins from the leaves of Ilex chinensis and their hepatoprotective activity. Chin. J. Nat. Med. 2021, 19, 376–384. [Google Scholar] [CrossRef]
- Ali, A.; Khalil, A.A.; Khuda, F.; Nazir, N.; Ullah, R.; Bari, A.; Haider, A.; Jamal, S.B.; Ahmad, S.; Khan, Z.; et al. Phytochemical and biological screening of leaf, bark and fruit extracts from Ilex dipyrena Wall. Life 2021, 11, 837. [Google Scholar] [CrossRef] [PubMed]
- Palu, D.; Bighelli, A.; Casanova, J.; Paoli, M. Identification and quantitation of ursolic and oleanolic acids in Ilex aquifolium L. leaf extracts using 13C and 1H-NMR spectroscopy. Molecules 2019, 24, 4413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, X. Resources investigation of indigenous plants in Nanchang and their application in urban landscape. In Proceedings of the 3rd International Conference on Civil, Architecture and Urban Engineering, Xiamen, China, 30 June 2021. [Google Scholar]
- Peterken, G.F.; Lloyd, P.S. Ilex aquifolium L. J. Ecol. 1967, 55, 841–858. [Google Scholar] [CrossRef]
- Tsang, A.C.W.; Corlett, R.T. Reproductive biology of the Ilex species (Aquifoliaceae) in Hong Kong, China. Can. J. Bot. 2005, 83, 1645–1654. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Yang, Y.; Liu, M.; Wang, B.; Li, M.; Chen, Y. Molecular tracing of the origin of six different plant species in bee honey using real-time PCR. J. AOAC Int. 2019, 100, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Bista, S.; Shivakoti, G.P. Honeybee flora at Kabre, Dolakha District. Nep. Agric. Res. J. 2000, 4, 18–25. [Google Scholar] [CrossRef]
- Bareke, T.; Addi, A. Bee flora diversity in different vegetation communities of Gsha-Sayilem forest in Kaffa Zone, south-western Ethiopia. Plants Environ. 2000, 2, 138–148. [Google Scholar] [CrossRef]
- Orwa, C.; Mutua, A.; Kindt, R.; Simons, A.; Jamnadass, R.H. Agroforestry Database 4: A Tree Reference and Selection Guide; World Agroforestry Centre: Nairobi, Kenya, 2010. [Google Scholar]
- Kerkvliet, J.D.; Beerlink, J.G. Pollen analysis of honeys from the coastal plain of Surinam. J. Apic. Res. 1991, 30, 25–31. [Google Scholar] [CrossRef]
- Lieux, M.H. Minor honeybee plants of Louisiana indicated by pollen analysis. Econ. Bot. 1978, 32, 418–432. [Google Scholar] [CrossRef]
- Grelen, H.E. Ilex opaca. In Silvics of North America: 2. Hardwoods. Agriculture Handbook 654; Burns, R.M., Honkala, B.H., Eds.; University of Michigan Library: Ann Arbor, MI, USA, 1990; pp. 379–385. [Google Scholar]
- Baas, P. The wood anatomical range in Ilex (Aquifoliaceae) and its ecological and phylogenetic significance. Blumea 1973, 21, 193–260. [Google Scholar]
- Shupe, T.F.; Aguilar, F.X.; Vlosky, R.P.; Belisle, M.; Chavez, A. Wood properties of selected lesser-used Honduran wood species. J. Trop. For. Sci. 2005, 17, 438–446. [Google Scholar]
- Maroyi, A. Ilex mitis (L.) Radlk. In PROTA (Plant Resources of Tropical Africa/Resources Végétales de l’Afrique Tropicale); Lemmens, R.H.M.J., Louppe, D., Oteng-Amoako, A.A., Eds.; Foundation PROTA: Wageningen, The Netherlands, 2012. [Google Scholar]
- Conn, B.J.; Damas, K.Q. Guide to Trees of Papua New Guinea. Available online: http://www.pngplants.org/PNGtrees (accessed on 10 April 2021).
- Wiselius, S.I. Ilex L. In Plant Resources of South-East Asia No. 5(3): Timber Trees; Lesser-Known, Timbers; Sosef, M.S.M., Hong, L.T., Prawirohatmodjo, S., Eds.; PROSEA Foundation: Bogor, Indonesia, 1998. [Google Scholar]
- Groppo, M. Neotropical Aquifoliaceae. In Neotropikey: Neotropical Flowering Plants; Milliken, W., Klitgaard, B., Baracat, A., Eds.; Royal Botanic Gardens, Kew: London, UK, 2009. [Google Scholar]
- Sensei, K.N. Dyeing and painting using plant pigments. Curtis’s Bot. Mag. 1999, 16, 134–136. [Google Scholar] [CrossRef]
- Zhu, L. Study on the extraction of natural dye from holly and its dyeing on silk fabric. Text. Aux. 2016, 33, 51–53. [Google Scholar]
- Giacomini, F.; Menegazzo, M.A.B.; dos Santos, J.C.O.; Arroyo, P.A.; de Barros, M.A.S.D. Ecofriendly dyeing of silk with extract of yerba mate (Ilex paraguariensis). Text. Res. J. 2017, 87, 829–837. [Google Scholar] [CrossRef]
- Vila, N.T.; Ferreira, A.; da Silva, M.G.; Fernandes, M.; Fiori, S. Surface modification of silk by (DBD) dielectric barrier discharge treatment for dyeing with natural dye yerba mate (Ilex paraguariensis). Procedia Eng. 2017, 200, 170–177. [Google Scholar] [CrossRef]
- Hamel, P.B.; Chiltoskey, M.U. Cherokee Plants and Their Uses—A 400 Year History; Herald Publishing Co. Inc.: Sylva, NC, USA, 1975. [Google Scholar]
- Manen, J.F.; Boulter, M.C.; Naciri-Graven, Y. The complex history of the genus Ilex L. (Aquifoliaceae): Evidence from the comparison of plastid and nuclear DNA sequences and from fossil data. Plant Syst. Evol. 2002, 235, 79–98. [Google Scholar] [CrossRef]
- Lebedev, V.G.; Lebedeva, T.N.; Chernodubov, A.I.; Shestibratov, K.A. Genomic selection for forest tree improvement: Methods, achievements and perspectives. Forests 2020, 11, 1190. [Google Scholar] [CrossRef]
- O’Connor, K.M.; Hayes, B.J.; Hardner, C.M.; Alam, M.; Henry, R.J.; Topp, B.L. Genomic selection and genetic gain for nut yield in an Australian Macadamia breeding population. BMC Genom. 2021, 22, 370. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Beverage | Species | Caffeine Content (mg/g) | Theobromine Content (mg/g) | Theophylline Content (mg/g) | Sources |
|---|---|---|---|---|---|
| kudingcha | Ilex latifolia | ND | ND | ND | [11] |
| Ilex kaushue | ND | ND | ND | [11] | |
| yerba maté | Ilex paraguariensis | 11.86 ± 0.07 | 0.86 ± 0.11 | 0.008 ± 0.001 | [11] |
| Ilex brevicuspis | ND | ND | ND | [12] | |
| Ilex theezans | NQA | ND | NQA | [12] | |
| Ilex microdonta | ND | ND | ND | [13] | |
| Ilex dumosa | NQA | ND | NQA | [12] | |
| Ilex taubertiana | ND | ND | ND | [14] | |
| Ilex pseudobuxus | NQA | ND | 0.6 ± 0.2 | [12] | |
| Ilex integerrima | ND | ND | ND | [14] | |
| Ilex argentina | ND | NQA | ND | [12] | |
| guayusa | Ilex guayusa | 26.94 ± 0.88 | 0.08 ± 0.02 | 0.004 ± 0.001 | [11] |
| yaupon | Ilex vomitoria | 8.44 ± 0.31 | 0.97 ± 0.14 | 0.014 ± 0.002 | [11] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, X.; Zhang, F.; Corlett, R.T. Utilization of the Hollies (Ilex L. spp.): A Review. Forests 2022, 13, 94. https://doi.org/10.3390/f13010094
Yao X, Zhang F, Corlett RT. Utilization of the Hollies (Ilex L. spp.): A Review. Forests. 2022; 13(1):94. https://doi.org/10.3390/f13010094
Chicago/Turabian StyleYao, Xin, Fan Zhang, and Richard T. Corlett. 2022. "Utilization of the Hollies (Ilex L. spp.): A Review" Forests 13, no. 1: 94. https://doi.org/10.3390/f13010094
APA StyleYao, X., Zhang, F., & Corlett, R. T. (2022). Utilization of the Hollies (Ilex L. spp.): A Review. Forests, 13(1), 94. https://doi.org/10.3390/f13010094