
Are Existing Modeling Tools Useful to Evaluate Outcomes in Mangrove Restoration and Rehabilitation Projects? A Minireview



Abstract
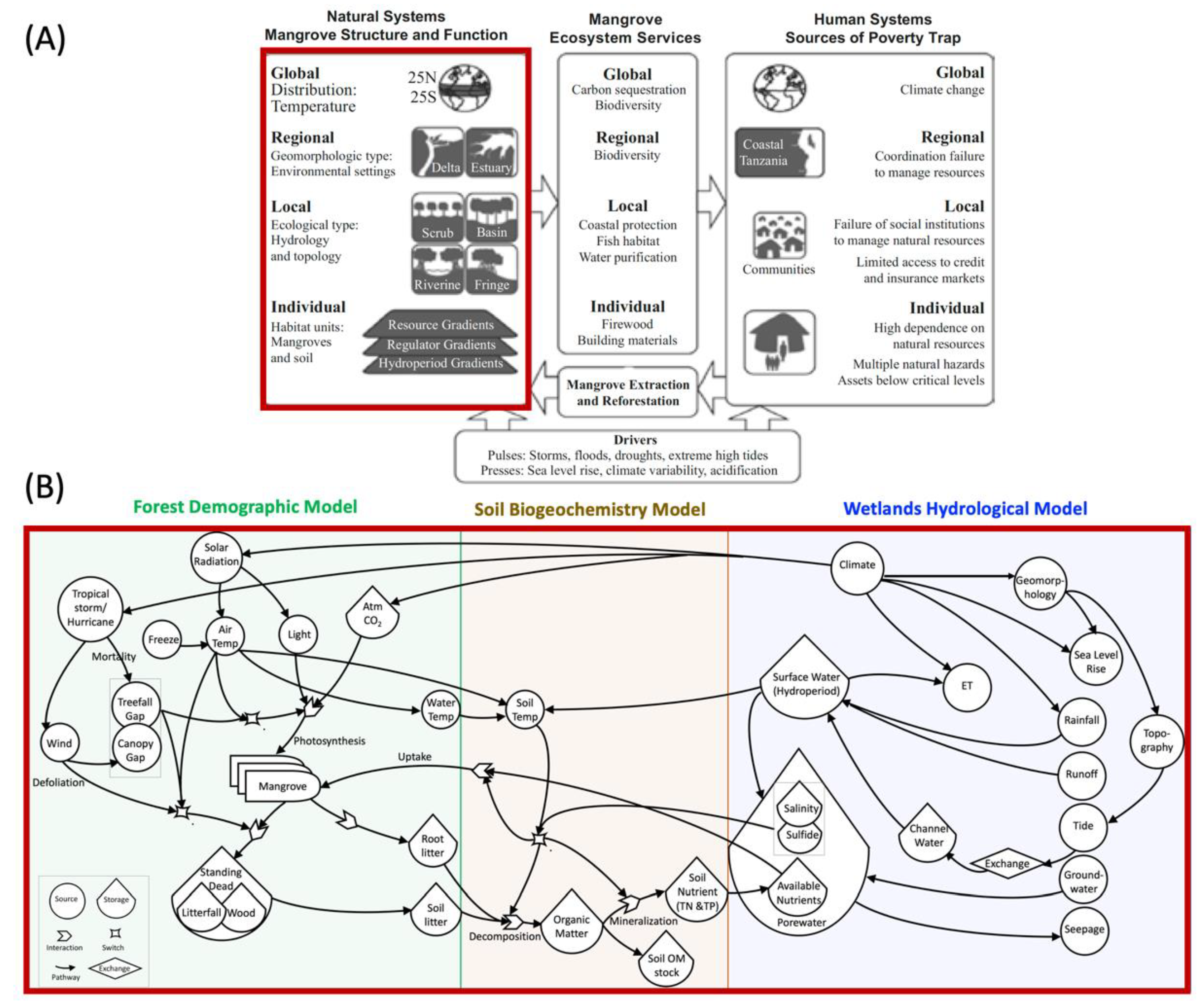
:1. Introduction
2. Materials and Methods
2.1. Literature Review
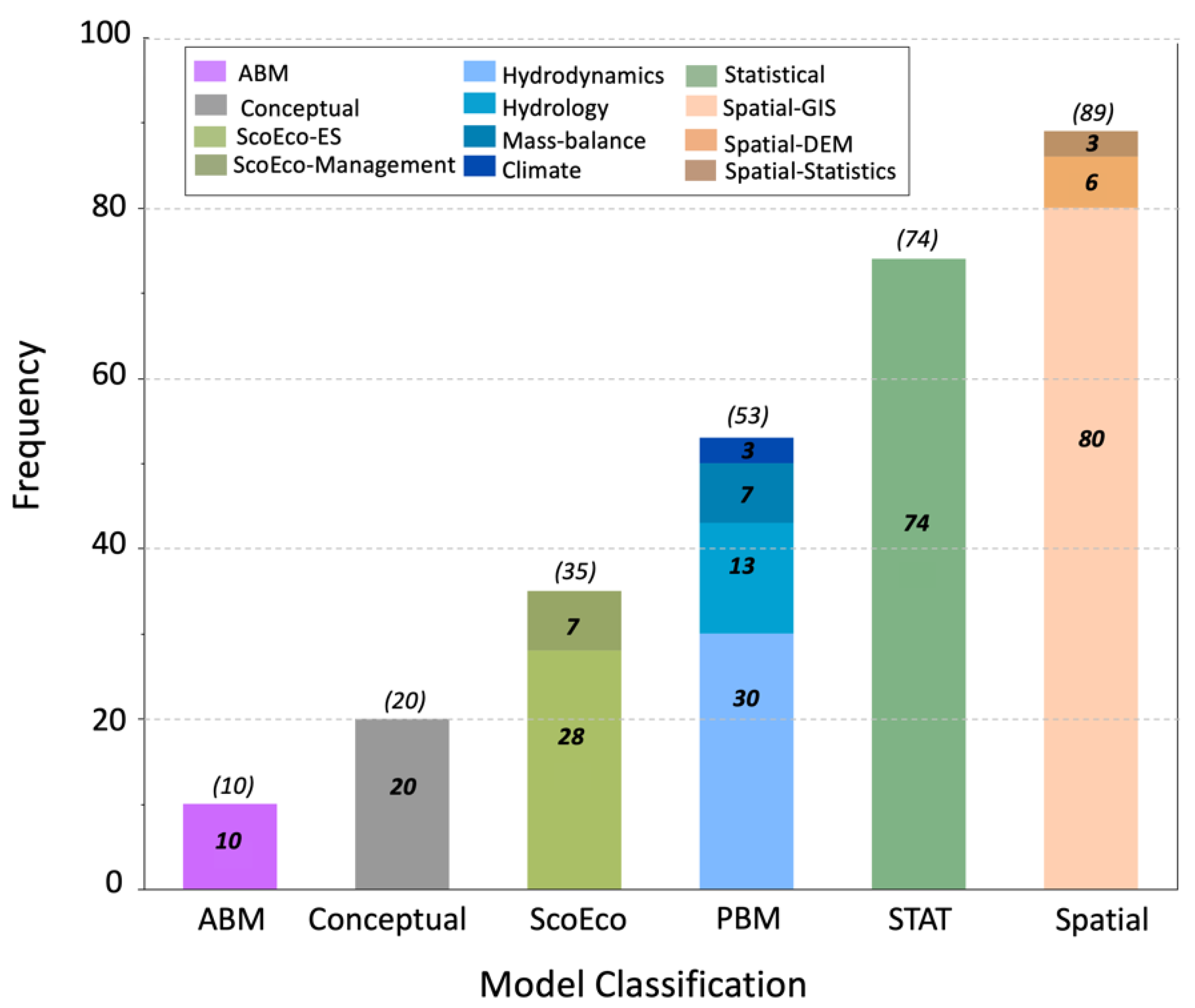
2.2. Model Classification
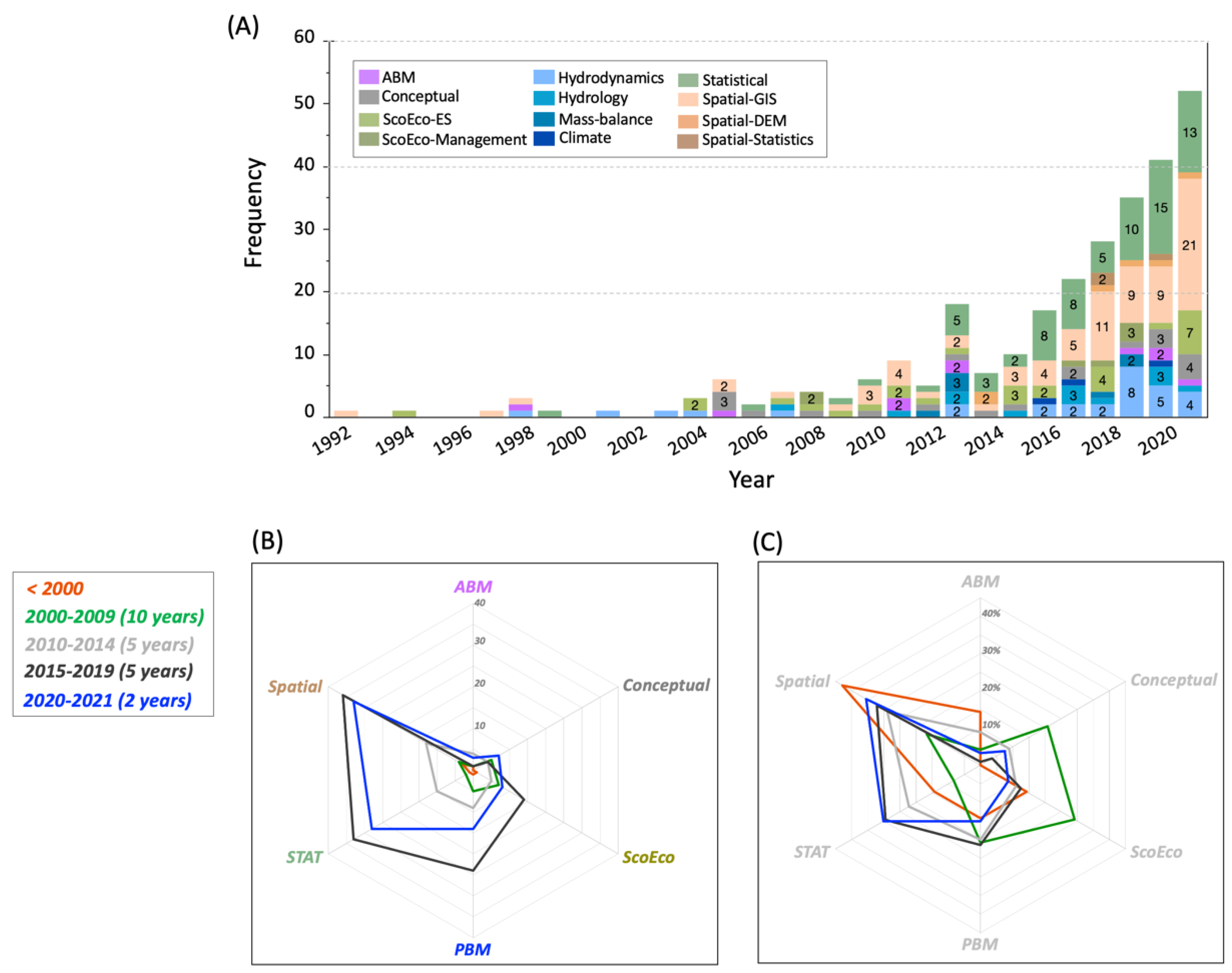
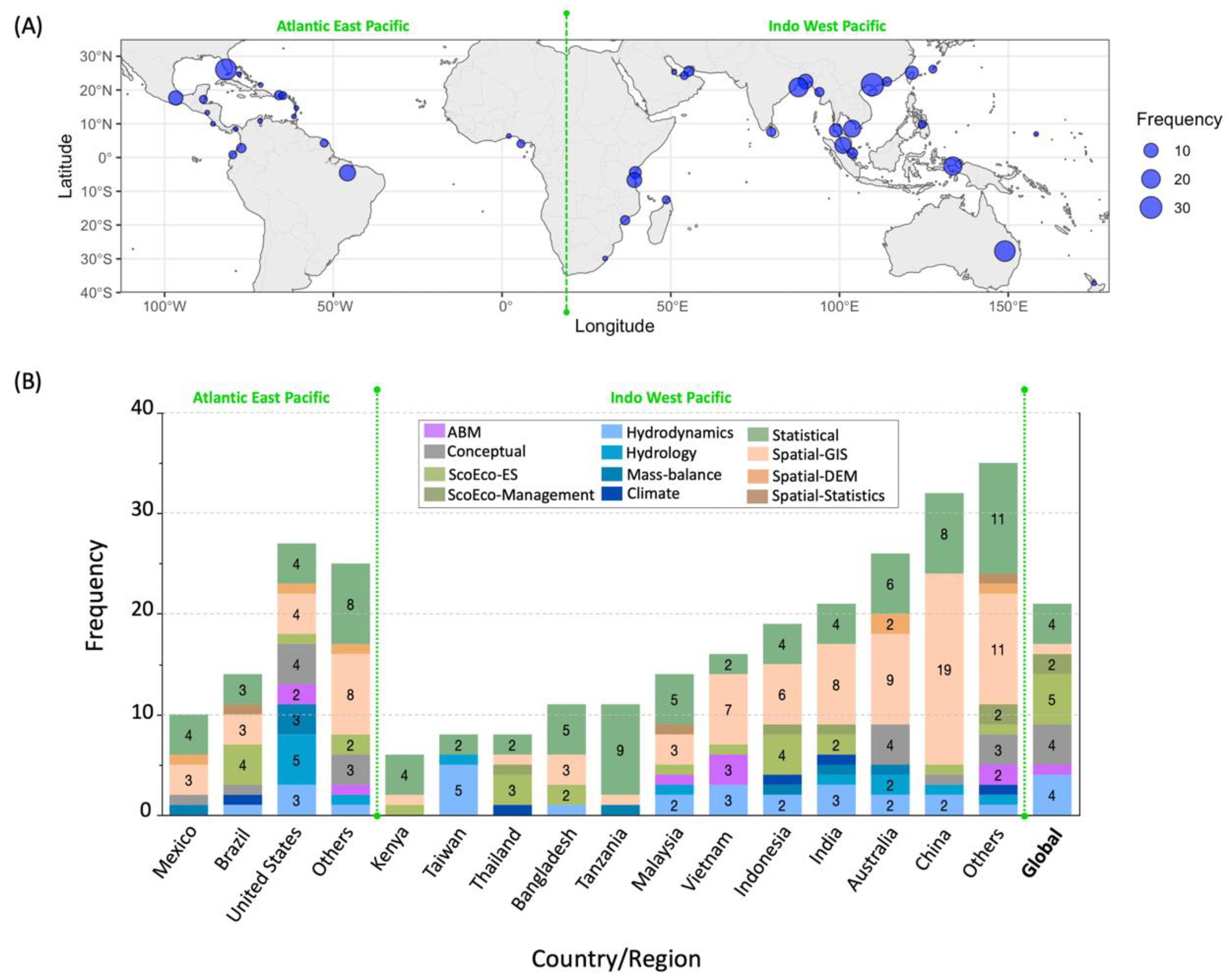
3. Results
4. Discussion
4.1. Mangrove Models Inventory and Utility in R/R Projects
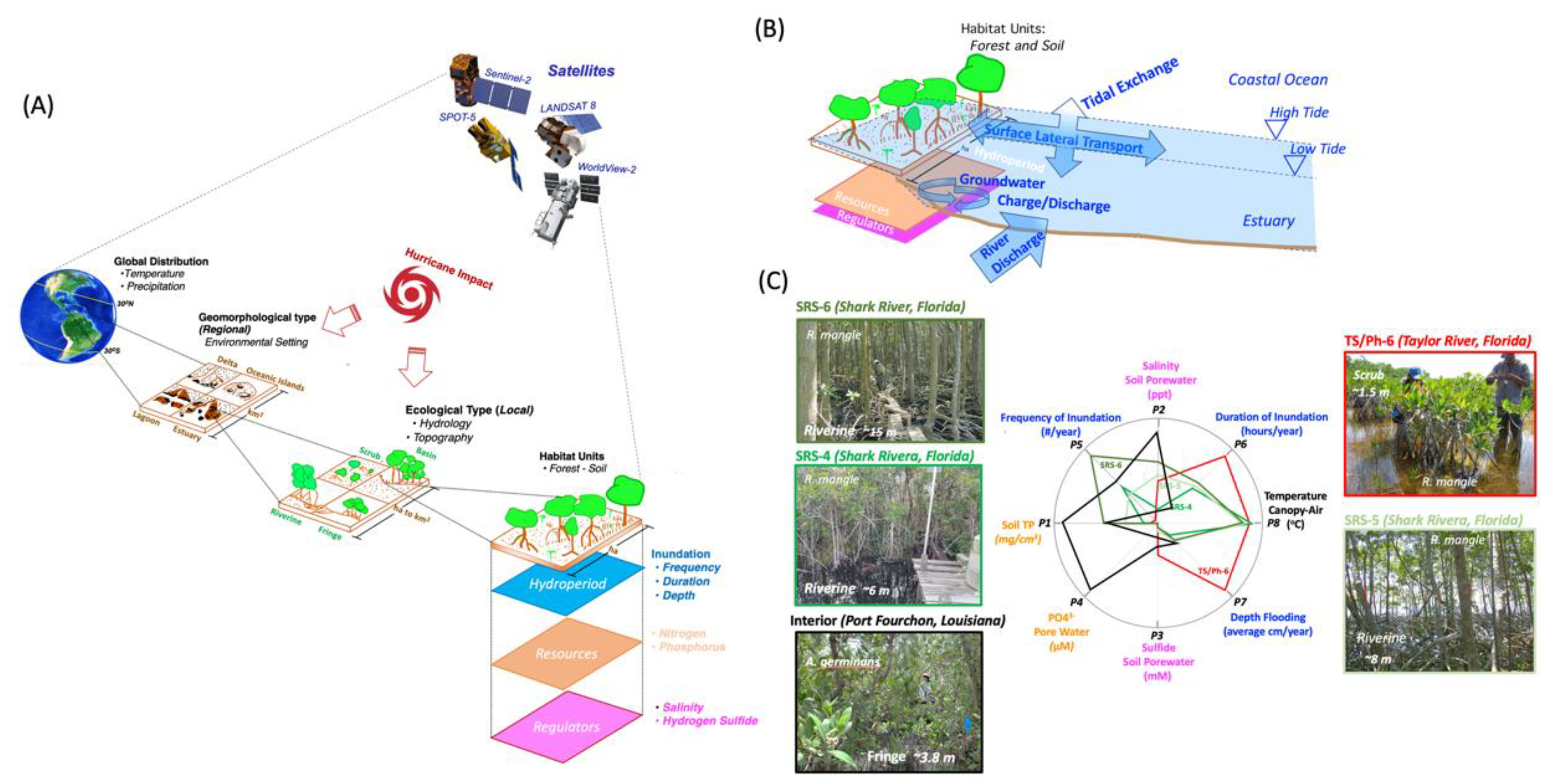
4.2. Agent and Process-Based Models
4.3. Static Spatial and Statistical Model Utility in Biogeochemical Studies
4.4. Implementing and Forecasting the Success of R/R Projects
4.5. A Multi-Modeling Approach Is Needed
5. Future Research Directions and Conclusions
5.1. Explicitly Linking Mangrove Models and ESs
5.2. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gann, G.D.; McDonald, T.; Walder, B.; Aronson, J.; Nelson, C.R.; Jonson, J.; Hallett, J.G.; Eisenberg, C.; Guariguata, M.R.; Liu, J.G.; et al. International principles and standards for the practice of ecological restoration; 2nd ed. Restor. Ecol. 2019, 27, S3–S46. [Google Scholar] [CrossRef]
- Field, C.D. Rehabilitation of mangrove ecosystems: An overview. Mar. Pollut. Bull. 1998, 37, 383–392. [Google Scholar] [CrossRef]
- Ellison, A.M.; Felson, A.J.; Friess, D.A. Mangrove Rehabilitation and Restoration as Experimental Adaptive Management. Front. Mar. Sci. 2020, 7, 19. [Google Scholar] [CrossRef]
- Ewel, K.C.; Twilley, R.R.; Ong, J.E. Different kinds of mangrove forests provide different goods and services. Glob. Ecol. Biogeogr. 1998, 7, 83–94. [Google Scholar] [CrossRef]
- Lugo, A.E. Mangrove Ecosystems: Successional or Steady State. Biotropica 1980, 12, 65–72. [Google Scholar] [CrossRef]
- Twilley, R.R.; Rivera-Monroy, V.H.; Chen, R.H.; Botero, L. Adapting an ecological mangrove model to simulate trajectories in restoration ecology. Mar. Pollut. Bull. 1998, 37, 404–419. [Google Scholar] [CrossRef]
- Bosire, J.O.; Dahdouh-Guebas, F.; Walton, M.; Crona, B.I.; Lewis, R.R.; Field, C.; Kairo, J.G.; Koedam, N. Functionality of restored mangroves: A review. Aquat. Bot. 2008, 89, 251–259. [Google Scholar] [CrossRef] [Green Version]
- López-Portillo, J.; Lewis, R.R.; Saenger, P.; Rovai, A.; Koedam, N.; Dahdouh-Guebas, F.; Agraz-Hernández, C.; Rivera-Monroy, V.H. Mangrove Forest Restoration and Rehabilitation. In Mangrove Ecosystems: A Global Biogeographic Perspective; Rivera-Monroy, V.H., Lee, S.Y., Kristensen, E., Twilley, R.R., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 301–345. [Google Scholar]
- Ellison, A.M. Mangrove Restoration: Do We Know Enough? Restor. Ecol. 2000, 8, 219–229. [Google Scholar] [CrossRef]
- Twilley, R.R.; Rivera-Monroy, V.H. Developing performance measures of mangrove wetlands using simulation models of hydrology, nutrient biogeochemistry, and community dynamics. J. Coastal. Res. 2005, 79–93. Available online: https://www.jstor.org/stable/25736617 (accessed on 17 August 2022).
- Lugo, A.E.; Snedaker, S.C. The Ecology of Mangroves. Annu. Rev. Ecol. Syst. 1974, 5, 39–64. [Google Scholar] [CrossRef]
- Berger, U.; Hildenbrandt, H. A new approach to spatially explicit modelling of forest dynamics: Spacing, ageing and neighbourhood competition of mangrove trees. Ecol. Model. 2000, 132, 287–302. [Google Scholar] [CrossRef]
- Mandal, S.; Ray, S.; Ghosh, P.B. Modelling of the contribution of dissolved inorganic nitrogen (DIN) from litterfall of adjacent mangrove forest to Hooghly-Matla estuary, India. Ecol. Model. 2009, 220, 2988–3000. [Google Scholar] [CrossRef]
- Ray, S.; Mukherjee, J.; Mandal, S. Chapter 13—Modelling nitrogen and carbon cycles in Hooghly estuary along with adjacent mangrove ecosystem. In Developments in Environmental Modelling; Park, Y.-S., Lek, S., Baehr, C., Jørgensen, S.E., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; Volume 27, pp. 289–320. [Google Scholar]
- Jiang, J.; DeAngelis, D.L.; Anderson, G.H.; Smith, T.J. Analysis and Simulation of Propagule Dispersal and Salinity Intrusion from Storm Surge on the Movement of a Marsh-Mangrove Ecotone in South Florida. Estuaries Coasts 2014, 37, 24–35. [Google Scholar] [CrossRef]
- Chen, R.; Twilley, R.R. A gap dynamic model of mangrove forest development along gradients of soil salinity and nutrient resources. J. Ecol. 1998, 86, 37–51. [Google Scholar] [CrossRef]
- Mazda, Y.; Wolanski, E. Hydrodynamics and modeling of water flow in mangrove areas. Coast. Wetl. Integr. Ecosyst. Approach 2009, 8, 231–261. [Google Scholar]
- Yoshikai, M.; Nakamura, T.; Suwa, R.; Sharma, S.; Rollon, R.; Yasuoka, J.; Egawa, R.; Nadaoka, K. Predicting mangrove forest dynamics across a soil salinity gradient using an individual-based vegetation model linked with plant hydraulics. Biogeosciences 2022, 19, 1813–1832. [Google Scholar] [CrossRef]
- Van Maanen, B.; Coco, G.; Bryan, K.R. On the ecogeomorphological feedbacks that control tidal channel network evolution in a sandy mangrove setting. Proc. Math. Phys. Eng. Sci. 2015, 471, 20150115. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.H.; Twilley, R.R. A simulation model of organic matter and nutrient accumulation in mangrove wetland soils. Biogeochemistry 1999, 44, 93–118. [Google Scholar] [CrossRef]
- Dai, Z.H.; Trettin, C.C.; Frolking, S.; Birdsey, R.A. Mangrove carbon assessment tool: Model validation and assessment of mangroves in southern USA and Mexico. Estuar. Coast. Shelf Sci. 2018, 208, 107–117. [Google Scholar] [CrossRef]
- Rivera-Monroy, V.H.; Osland, M.J.; Day, J.W.; Ray, S.; Rovai, A.; Day, R.H.; Mukherjee, J. Advancing Mangrove Macroecology. In Mangrove Ecosystems: A Global Biogeographic Perspective; Rivera-Monroy, V.H., Lee, S.Y., Kristensen, E., Twilley, R.R., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 347–381. [Google Scholar]
- Clarke, P.J. Seeking global generality: A critique for mangrove modellers. Mar. Freshw. Res. 2014, 65, 930–933. [Google Scholar] [CrossRef]
- Botkin, D.B.; Janak, J.F.; Wallis, J.R. Some Ecological Consequences of a Computer Model of Forest Growth. J. Ecol. 1972, 60, 849–872. [Google Scholar] [CrossRef] [Green Version]
- Mitsch, W.J.; Straškraba, M.; Jorgensen, S.E. Wetland Modelling; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Shugart, H.H.; Foster, A.; Wang, B.; Druckenbrod, D.; Ma, J.Y.; Lerdau, M.; Saatchi, S.; Yang, X.; Yan, X.D. Gap models across micro- to mega-scales of time and space: Examples of Tansley’s ecosystem concept. For. Ecosyst. 2020, 7, 18. [Google Scholar] [CrossRef]
- Barbier, E.B.; Hacker, S.D.; Kennedy, C.; Koch, E.W.; Stier, A.C.; Silliman, B.R. The value of estuarine and coastal ecosystem services. Ecol. Monogr. 2011, 81, 169–193. [Google Scholar] [CrossRef]
- Taillardat, P.; Friess, D.A.; Lupascu, M. Mangrove blue carbon strategies for climate change mitigation are most effective at the national scale. Biol. Lett. 2018, 14, 20180251. [Google Scholar] [CrossRef] [Green Version]
- Rovai, A.S.; Twilley, R.R.; Castañeda-Moya, E.; Riul, P.; Cifuentes-Jara, M.; Manrow-Villalobos, M.; Horta, P.A.; Simonassi, J.C.; Fonseca, A.L.; Pagliosa, P.R. Global controls on carbon storage in mangrove soils. Nat. Clim. Chang. 2018, 8, 534–538. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Reef, R. Variable Impacts of Climate Change on Blue Carbon. One Earth 2020, 3, 195–211. [Google Scholar] [CrossRef]
- Hamilton, S.E.; Friess, D.A. Global carbon stocks and potential emissions due to mangrove deforestation from 2000 to 2012. Nat. Clim. Chang. 2018, 8, 240. [Google Scholar] [CrossRef] [Green Version]
- Uchida, E.; Rivera-Monroy, V.H.; Ates, S.A.; Castañeda-Moya, E.; Gold, A.J.; Guilfoos, T.; Hernandez, M.F.; Lokina, R.; Mangora, M.M.; Midway, S.R.; et al. Collaborative Research Across Boundaries: Mangrove Ecosystem Services and Poverty Traps as a Coupled Natural-Human System. In Collaboration across Boundaries for Social-Ecological Systems Science; Perz, S.G., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 115–152. [Google Scholar]
- Lugo, A.E.; Medina, E. Mangrove forests. In Encyclopedia of Natural Resources-Land; Taylor and Francis: New York, NY, USA, 2014; Volume 1, pp. 343–352. [Google Scholar]
- Osland, M.J.; Feher, L.C.; Lopez-Portillo, J.; Day, R.H.; Suman, D.O.; Menendez, J.M.G.; Rivera-Monroy, V.H. Mangrove forests in a rapidly changing world: Global change impacts and conservation opportunities along the Gulf of Mexico coast. Estuar. Coast. Shelf Sci. 2018, 214, 120–140. [Google Scholar] [CrossRef]
- Friess, D.A.; Krauss, K.W.; Horstman, E.M.; Balke, T.; Bouma, T.J.; Galli, D.; Webb, E.L. Are all intertidal wetlands naturally created equal? Bottlenecks, thresholds and knowledge gaps to mangrove and saltmarsh ecosystems. Biol. Rev. Camb. Philos. Soc. 2012, 87, 346–366. [Google Scholar] [CrossRef]
- Brown, S.; Lugo, A.E. Rehabilitation of Tropical Lands: A Key to Sustaining Development. Restor. Ecol. 1994, 2, 97–111. [Google Scholar] [CrossRef]
- Yan, X.D.; Shugart, H.H. FAREAST: A forest gap model to simulate dynamics and patterns of eastern Eurasian forests. J. Biogeogr. 2005, 32, 1641–1658. [Google Scholar] [CrossRef]
- Fischer, R.; Bohn, F.; de Paula, M.D.; Dislich, C.; Groeneveld, J.; Gutierrez, A.G.; Kazmierczak, M.; Knapp, N.; Lehmann, S.; Paulick, S.; et al. Lessons learned from applying a forest gap model to understand ecosystem and carbon dynamics of complex tropical forests. Ecol. Model. 2016, 326, 124–133. [Google Scholar] [CrossRef]
- Giri, C.; Ochieng, E.; Tieszen, L.L.; Zhu, Z.; Singh, A.; Loveland, T.; Masek, J.; Duke, N. Status and distribution of mangrove forests of the world using earth observation satellite data. Glob. Ecol. Biogeogr. 2011, 20, 154–159. [Google Scholar] [CrossRef]
- Bunting, P.; Rosenqvist, A.; Lucas, R.M.; Rebelo, L.-M.; Hilarides, L.; Thomas, N.; Hardy, A.; Itoh, T.; Shimada, M.; Finlayson, C.M. The Global Mangrove Watch—A New 2010 Global Baseline of Mangrove Extent. Remote Sens. 2018, 10, 1669. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.H.; Twilley, R.R. Patterns of mangrove forest structure and soil nutrient dynamics along the Shark River estuary, Florida. Estuaries 1999, 22, 955–970. [Google Scholar] [CrossRef]
- Danielson, T.M.; Rivera-Monroy, V.H.; Castaneda-Moya, E.; Briceno, H.; Travieso, R.; Marx, B.D.; Gaiser, E.; Farfan, L.M. Assessment of Everglades mangrove forest resilience: Implications for above-ground net primary productivity and carbon dynamics. For. Ecol. Manag. 2017, 404, 115–125. [Google Scholar] [CrossRef]
- Rivera-Monroy, V.H.; Danielson, T.M.; Castaneda-Moya, E.; Marx, B.D.; Travieso, R.; Zhao, X.C.; Gaiser, E.E.; Farfan, L.M. Long-term demography and stem productivity of Everglades mangrove forests (Florida, USA): Resistance to hurricane disturbance. For. Ecol. Manag. 2019, 440, 79–91. [Google Scholar] [CrossRef]
- Odum, H.T. Systems Ecology; An Introduction; 1983. Available online: https://www.osti.gov/biblio/5545893 (accessed on 17 August 2022).
- Ouyang, X.; Lee, S.Y.; Connolly, R.M. Structural equation modelling reveals factors regulating surface sediment organic carbon content and CO2 efflux in a subtropical mangrove. Sci. Total Environ. 2017, 578, 513–522. [Google Scholar] [CrossRef] [Green Version]
- Duke, N.C. Mangrove Floristics and Biogeography Revisited: Further Deductions from Biodiversity Hot Spots, Ancestral Discontinuities, and Common Evolutionary Processes. In Mangrove Ecosystems: A Global Biogeographic Perspective: Structure, Function, and Services; Rivera-Monroy, V.H., Lee, S.Y., Kristensen, E., Twilley, R.R., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 17–53. [Google Scholar]
- Hsu, M.H.; Kuo, A.Y.; Kuo, J.T.; Liu, W.C. Modeling estuarine hydrodynamics and salinity for wetland restoration. J. Environ. Sci. Health Part A 1998, 33, 891–921. [Google Scholar] [CrossRef]
- Lewis, R.R., 3rd; Milbrandt, E.C.; Brown, B.; Krauss, K.W.; Rovai, A.S.; Beever, J.W., 3rd; Flynn, L.L. Stress in mangrove forests: Early detection and preemptive rehabilitation are essential for future successful worldwide mangrove forest management. Mar. Pollut. Bull. 2016, 109, 764–771. [Google Scholar] [CrossRef]
- Kurniawan, A.; Hasan, G.M.J.; Ooi, S.K.; Kit, L.W.; Loh, L.L.; Bayen, S. Understanding Hydrodynamic Flow Characteristics in a Model Mangrove Ecosystem in Singapore. APCBEE Procedia 2014, 10, 286–291. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.C.; Rivera-Monroy, V.H.; Wang, H.Q.; Xue, Z.G.; Tsai, C.F.; Willson, C.S.; Castaneda-Moya, E.; Twilley, R.R. Modeling soil porewater salinity in mangrove forests (Everglades, Florida, USA) impacted by hydrological restoration and a warming climate. Ecol. Model. 2020, 436, 109292. [Google Scholar] [CrossRef]
- Bathmann, J.; Peters, R.; Reef, R.; Berger, U.; Walther, M.; Lovelock, C.E. Modelling mangrove forest structure and species composition over tidal inundation gradients: The feedback between plant water use and porewater salinity in an arid mangrove ecosystem. Agric. For. Meteorol. 2021, 308, 10. [Google Scholar] [CrossRef]
- Kamil, E.A.; Takaijudin, H.; Hashim, A.M. Mangroves As Coastal Bio-Shield: A Review of Mangroves Performance in Wave Attenuation. Civ. Eng. J. 2021, 7, 1964–1981. [Google Scholar] [CrossRef]
- Menéndez, P.; Losada, I.J.; Torres-Ortega, S.; Narayan, S.; Beck, M.W. The Global Flood Protection Benefits of Mangroves. Sci. Rep. 2020, 10, 4404. [Google Scholar] [CrossRef] [Green Version]
- McIvor, A.L.; Möller, I.; Spencer, T.; Spalding, M. Reduction of Wind and Swell Waves by Mangroves; 2012; p. 27. Available online: http://www.naturalcoastalprotection.org/documents/reduction-of-wind-and-swell-wavesby-mangroves (accessed on 17 August 2022).
- Berger, U.; Rivera-Monroy, V.H.; Doyle, T.W.; Dahdouh-Guebas, F.; Duke, N.C.; Fontalvo-Herazo, M.L.; Hildenbrandt, H.; Koedam, N.; Mehlig, U.; Piou, C.; et al. Advances and limitations of individual-based models to analyze and predict dynamics of mangrove forests: A review. Aquat. Bot. 2008, 89, 260–274. [Google Scholar] [CrossRef] [Green Version]
- Berger, U.; Adams, M.; Grimm, V.; Hildenbrandt, H. Modelling secondary succession of neotropical mangroves: Causes and consequences of growth reduction in pioneer species. Perspect. Plant Ecol. 2006, 7, 243–252. [Google Scholar] [CrossRef]
- Fontalvo-Herazo, M.L.; Piou, C.; Vogt, J.; Saint-Paul, U.; Berger, U. Simulating harvesting scenarios towards the sustainable use of mangrove forest plantations. Wetl. Ecol. Manag. 2011, 19, 397–407. [Google Scholar] [CrossRef]
- Kautz, M.; Berger, U.; Stoyan, D.; Vogt, J.; Khan, N.I.; Diele, K.; Saint-Paul, U.; Triet, T.; Nam, V.N. Desynchronizing effects of lightning strike disturbances on cyclic forest dynamics in mangrove plantations. Aquat. Bot. 2011, 95, 173–181. [Google Scholar] [CrossRef]
- Khan, M.N.I.; Sharma, S.; Berger, U.; Koedam, N.; Dahdouh-Guebas, F.; Hagihara, A. How do tree competition and stand dynamics lead to spatial patterns in monospecific mangroves? Biogeosciences 2013, 10, 2803–2814. [Google Scholar] [CrossRef] [Green Version]
- Vogt, J.; Kautz, M.; Fontalvo Herazo, M.L.; Triet, T.; Walther, D.; Saint-Paul, U.; Diele, K.; Berger, U. Do canopy disturbances drive forest plantations into more natural conditions?—A case study from Can Gio Biosphere Reserve, Viet Nam. Glob. Planet. Chang. 2013, 110, 249–258. [Google Scholar] [CrossRef]
- Jenke, M.; Pranchai, A.; Peters, R.; Berger, U. Silvicultural options for the transformation of even-aged Rhizophora apiculata stands to irregular and diverse mangroves—A simulation experiment. Estuar. Coast. Shelf Sci. 2021, 248, 106886. [Google Scholar] [CrossRef]
- Grueters, U.; Seltmann, T.; Schmidt, H.; Horn, H.; Pranchai, A.; Vovides, A.G.; Peters, R.; Vogt, J.; Dahdouh-Guebas, F.; Berger, U. The mangrove forest dynamics model mesoFON. Ecol. Model. 2014, 291, 28–41. [Google Scholar] [CrossRef]
- Grueters, U.; Ibrahim, M.R.; Satyanarayana, B.; Dahdouh-Guebas, F. Individual-based modeling of mangrove forest growth: MesoFON—Recent calibration and future direction. Estuar. Coast. Shelf Sci. 2019, 227, 106302. [Google Scholar] [CrossRef] [Green Version]
- Peters, R.; Vovides, A.G.; Luna, S.; Gruters, U.; Berger, U. Changes in allometric relations of mangrove trees due to resource availability—A new mechanistic modelling approach. Ecol. Model. 2014, 283, 53–61. [Google Scholar] [CrossRef]
- Peters, R.; Walther, M.; Lovelock, C.; Jiang, J.; Berger, U. The interplay between vegetation and water in mangroves: New perspectives for mangrove stand modelling and ecological research. Wetl. Ecol. Manag. 2020, 28, 697–712. [Google Scholar] [CrossRef]
- Wimmler, M.C.; Bathmann, J.; Peters, R.; Jiang, J.; Walther, M.; Lovelock, C.E.; Berger, U. Plant-soil feedbacks in mangrove ecosystems: Establishing links between empirical and modelling studies. Trees 2021, 35, 1423–1438. [Google Scholar] [CrossRef]
- Bathmann, J.; Peters, R.; Naumov, D.; Fischer, T.; Berger, U.; Walther, M. The MANgrove-GroundwAter feedback model (MANGA)—Describing belowground competition based on first principles. Ecol. Model. 2020, 420, 11. [Google Scholar] [CrossRef]
- Teh, S.Y.; Koh, H.L.; DeAngelis, D.L.; Turtora, M. Interaction between salinity intrusion and vegetation succession: A modeling approach. Theor. Appl. Mech. Lett. 2013, 3, 032001. [Google Scholar] [CrossRef] [Green Version]
- Bridgham, S.D.; Cadillo-Quiroz, H.; Keller, J.K.; Zhuang, Q. Methane emissions from wetlands: Biogeochemical, microbial, and modeling perspectives from local to global scales. Glob. Chang. Biol. 2013, 19, 1325–1346. [Google Scholar] [CrossRef]
- Luo, Z.K.; Sun, O.J.; Wang, E.L.; Ren, H.; Xu, H.L. Modeling Productivity in Mangrove Forests as Impacted by Effective Soil Water Availability and Its Sensitivity to Climate Change Using Biome-BGC. Ecosystems 2010, 13, 949–965. [Google Scholar] [CrossRef]
- Gauci, V.; Matthews, E.; Dise, N.; Walter, B.; Koch, D.; Granberg, G.; Vile, M. Sulfur pollution suppression of the wetland methane source in the 20th and 21st centuries. Proc. Natl. Acad. Sci. USA 2004, 101, 12583–12587. [Google Scholar] [CrossRef]
- Rosentreter, J.A.; Maher, D.T.; Erler, D.V.; Murray, R.H.; Eyre, B.D. Methane emissions partially offset “blue carbon” burial in mangroves. Sci. Adv. 2018, 4, eaao4985. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.D.; Sun, C.Y.; Qin, Z.C. Drought-Induced Salinity Enhancement Weakens Mangrove Greennouse Gas Cycling. J. Geophys. Res. Biogeosci. 2021, 126, 18. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Feller, I.C.; Reef, R.; Hickey, S.; Ball, M.C. Mangrove dieback during fluctuating sea levels. Sci. Rep. 2017, 7, 1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maina, J.M.; Bosire, J.O.; Kairo, J.G.; Bandeira, S.O.; Mangora, M.M.; Macamo, C.; Ralison, H.; Majambo, G. Identifying global and local drivers of change in mangrove cover and the implications for management. Glob. Ecol. Biogeogr. 2021, 30, 2057–2069. [Google Scholar] [CrossRef]
- Wang, L.; Jia, M.; Yin, D.; Tian, J. A review of remote sensing for mangrove forests: 1956–2018. Remote Sens. Environ. 2019, 231, 111223. [Google Scholar] [CrossRef]
- Arifanti, V.B.; Sidik, F.; Mulyanto, B.; Susilowati, A.; Wahyuni, T.; Subarno; Yulianti; Yuniarti, N.; Aminah, A.; Suita, E.; et al. Challenges and Strategies for Sustainable Mangrove Management in Indonesia: A Review. Forests 2022, 13, 695. [Google Scholar] [CrossRef]
- Rovai, A.S.; Twilley, R.R.; Castaneda-Moya, E.; Midway, S.R.; Friess, D.A.; Trettin, C.C.; Bukoski, J.J.; Stovall, A.E.L.; Pagliosa, P.R.; Fonseca, A.L.; et al. Macroecological patterns of forest structure and allometric scaling in mangrove forests. Glob. Ecol. Biogeogr. 2021, 30, 1000–1013. [Google Scholar] [CrossRef]
- Twilley, R.R.; Castañeda-Moya, E.; Rivera-Monroy, V.H.; Rovai, A. Productivity and Carbon Dynamics in Mangrove Wetlands. In Mangrove Ecosystems: A Global Biogeographic Perspective: Structure, Function, and Services; Rivera-Monroy, V.H., Lee, S.Y., Kristensen, E., Twilley, R.R., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 113–162. [Google Scholar]
- Rodríguez-Zúñiga, M.T.; Troche-Souza, C.; Cruz-López, M.I.; Rivera-Monroy, V.H. Development and Structural Organization of Mexico’s Mangrove Monitoring System (SMMM) as a Foundation for Conservation and Restoration Initiatives: A Hierarchical Approach. Forests 2022, 13, 621. [Google Scholar] [CrossRef]
- Zhao, X.C.; Rivera-Monroy, V.H.; Li, C.Y.; Vargas-Lopez, I.A.; Rohli, R.V.; Xue, Z.G.; Castaneda-Moya, E.; Coronado-Molina, C. Temperature across Vegetation Canopy-Water-Soil Interfaces Is Modulated by Hydroperiod and Extreme Weather in Coastal Wetlands. Front. Mar. Sci. 2022, 9, 21. [Google Scholar] [CrossRef]
- Maher, D.T.; Call, M.; Santos, I.R.; Sanders, C.J. Beyond burial: Lateral exchange is a significant atmospheric carbon sink in mangrove forests. Biol. Lett. 2018, 14, 4. [Google Scholar] [CrossRef]
- Regnier, P.; Resplandy, L.; Najjar, R.G.; Ciais, P. The land-to-ocean loops of the global carbon cycle. Nature 2022, 603, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Rivera-Monroy, V.H.; Farfan, L.M.; Briceno, H.; Castaneda-Moya, E.; Travieso, R.; Gaiser, E.E. Tropical cyclones cumulatively control regional carbon fluxes in Everglades mangrove wetlands (Florida, USA). Sci. Rep. 2021, 11, 13927. [Google Scholar] [CrossRef]
- Adame, M.F.; Zakaria, R.M.; Fry, B.; Chong, V.C.; Then, Y.H.A.; Brown, C.J.; Lee, S.Y. Loss and recovery of carbon and nitrogen after mangrove clearing. Ocean Coast. Manag. 2018, 161, 117–126. [Google Scholar] [CrossRef]
- Dontis, E.E.; Radabaugh, K.R.; Chappel, A.R.; Russo, C.E.; Moyer, R.P. Carbon Storage Increases with Site Age as Created Salt Marshes Transition to Mangrove Forests in Tampa Bay, Florida (USA). Estuaries Coasts 2020, 43, 1470–1488. [Google Scholar] [CrossRef]
- Bouillon, S.; Borges, A.V.; Castaneda-Moya, E.; Diele, K.; Dittmar, T.; Duke, N.C.; Kristensen, E.; Lee, S.Y.; Marchand, C.; Middelburg, J.J.; et al. Mangrove production and carbon sinks: A revision of global budget estimates. Glob. Biogeochem. Cy. 2008, 22, 12. [Google Scholar] [CrossRef] [Green Version]
- Ward, N.D.; Megonigal, J.P.; Bond-Lamberty, B.; Bailey, V.L.; Butman, D.; Canuel, E.A.; Diefenderfer, H.; Ganju, N.K.; Goni, M.A.; Graham, E.B.; et al. Representing the function and sensitivity of coastal interfaces in Earth system models. Nat. Commun. 2020, 11, 2458. [Google Scholar] [CrossRef]
- Twilley, R.R.; Rivera-Monroy, V.H.; Rovai, A.S.; Castañeda-Moya, E.; Davis, S. Mangrove Biogeochemistry at Local to Global Scales Using Ecogeomorphic Approaches. In Coastal Wetlands; Perillo, G.M.E., Wolanski, E., Cahoon, D.R., Hopkinson, C.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 717–785. [Google Scholar]
- Twilley, R.R.; Chen, R.H. A water budget and hydrology model of a basin mangrove forest in Rookery Bay, Florida. Mar. Freshw. Res. 1998, 49, 309–323. [Google Scholar] [CrossRef]
- Castañeda-Moya, E.; Twilley, R.R.; Rivera-Monroy, V.H.; Marx, B.D.; Coronado-Molina, C.; Ewe, S.M. Patterns of root dynamics in mangrove forests along environmental gradients in the Florida Coastal Everglades, USA. Ecosystems 2011, 14, 1178–1195. [Google Scholar] [CrossRef]
- Medina-Calderon, J.H.; Mancera-Pineda, J.E.; Castaneda-Moya, E.; Rivera-Monroy, V.H. Hydroperiod and Salinity Interactions Control Mangrove Root Dynamics in a Karstic Oceanic Island in the Caribbean Sea (San Andres, Colombia). Front. Mar. Sci. 2021, 7, 598132. [Google Scholar] [CrossRef]
- Feller, I.C.; Lovelock, C.E.; Berger, U.; McKee, K.L.; Joye, S.B.; Ball, M.C. Biocomplexity in mangrove ecosystems. Annu. Rev. Mar. Sci. 2010, 2, 395–417. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.A.; Rose, K.A.; de Mutsert, K.; Sable, S.; Ainsworth, C.; Brady, D.C.; Townsend, H. Using Multiple Ecological Models to Inform Environmental Decision-Making. Front. Mar. Sci. 2021, 8, 20. [Google Scholar] [CrossRef]
- Proisy, C.; Degenne, P.; Anthony, E.J.; Berger, U.; Blanchard, E.; Fromard, F.; Gardel, A.; Olagoke, A.; Santos, V.; Walcker, R.; et al. A Multiscale Simulation Approach for Linking Mangrove Dynamics to Coastal Processes using Remote Sensing Observations. J. Coast. Res. 2016, 75, 810–814. [Google Scholar] [CrossRef]
- Mukherjee, J.; Bhowmick, A.R.; Ghosh, P.B.; Ray, S. Impact of environmental factors on the dependency of litter biomass in carbon cycling of Hooghly estuary, India. Ecol. Inform. 2019, 51, 193–200. [Google Scholar] [CrossRef]
- Box, G.E.P. Science and Statistics. J. Am. Stat. Assoc. 1976, 71, 791–799. [Google Scholar] [CrossRef]
- Fulton, E.A.; Blanchard, J.L.; Melbourne-Thomas, J.; Plagányi, É.E.; Tulloch, V.J.D. Where the Ecological Gaps Remain, a Modelers’ Perspective. Front. Ecol. Evol. 2019, 7, 424. [Google Scholar] [CrossRef] [Green Version]
- Castaneda-Moya, E.; Rivera-Monroy, V.H.; Chambers, R.M.; Zhao, X.; Lamb-Wotton, L.; Gorsky, A.; Gaiser, E.E.; Troxler, T.G.; Kominoski, J.S.; Hiatt, M. Hurricanes fertilize mangrove forests in the Gulf of Mexico (Florida Everglades, USA). Proc. Natl. Acad. Sci. USA 2020, 117, 4831–4841. [Google Scholar] [CrossRef]
- Bayraktarov, E.; Brisbane, S.; Hagger, V.; Smith, C.S.; Wilson, K.A.; Lovelock, C.E.; Gillies, C.; Steven, A.D.L.; Saunders, M.I. Priorities and Motivations of Marine Coastal Restoration Research. Front. Mar. Sci. 2020, 7, 484. [Google Scholar] [CrossRef]
- Su, J.; Friess, D.A.; Gasparatos, A. A meta-analysis of the ecological and economic outcomes of mangrove restoration. Nat. Commun. 2021, 12, 5050. [Google Scholar] [CrossRef] [PubMed]
- Barbier, E.B. The protective service of mangrove ecosystems: A review of valuation methods. Mar. Pollut. Bull. 2016, 109, 676–681. [Google Scholar] [CrossRef]
- Himes-Cornell, A.; Grose, S.O.; Pendleton, L. Mangrove Ecosystem Service Values and Methodological Approaches to Valuation: Where Do We Stand? Front. Mar. Sci. 2018, 5, 376. [Google Scholar] [CrossRef]
- Robertson, M.D.; Midway, S.R.; West, L.; Tillya, H.; Rivera-Monroy, V.H. Fishery characteristics in two districts of coastal Tanzania. Ocean Coast. Manag. 2018, 163, 254–268. [Google Scholar] [CrossRef]
- Shinnaka, T.; Sano, M.; Ikejima, K.; Tongnunui, P.; Horinouchi, M.; Kurokura, H. Effects of mangrove deforestation on fish assemblage at Pak Phanang Bay, southern Thailand. Fish. Sci. 2007, 73, 862–870. [Google Scholar] [CrossRef]
- Rivera-Monroy, V.H.; Twilley, R.R.; Mancera, E.; Alcantara-Eguren, A.; Castañeda-Moya, E.; Casas- Monroy, O.; Reyes, F.; Restrepo, J.; Perdomo, L.; Campos, E.; et al. Adventures and misfortunes in Macondo: Rehabilitation of the Cienaga Grande de Santa Marta Lagoon Complex, Colombia. Ecotropicos 2006, 19, 72–93. [Google Scholar]
- Lee, S.Y.; Hamilton, S.; Barbier, E.B.; Primavera, J.; Lewis, R.R., 3rd. Better restoration policies are needed to conserve mangrove ecosystems. Nat. Ecol. Evol. 2019, 3, 870–872. [Google Scholar] [CrossRef] [PubMed]
- Nerem, R.S.; Beckley, B.D.; Fasullo, J.T.; Hamlington, B.D.; Masters, D.; Mitchum, G.T. Climate-change–driven accelerated sea-level rise detected in the altimeter era. Proc. Natl. Acad. Sci. USA 2018, 115, 2022–2025. [Google Scholar] [CrossRef] [Green Version]
- Chambers, R.M.; Gorsky, A.L.; Castañeda-Moya, E.; Rivera-Monroy, V.H. Evaluating a Steady-State Model of Soil Accretion in Everglades Mangroves (Florida, USA). Estuaries Coasts 2021, 44, 1469–1476. [Google Scholar] [CrossRef]
- Saintilan, N.; Khan, N.S.; Ashe, E.; Kelleway, J.J.; Rogers, K.; Woodroffe, C.D.; Horton, B.P. Thresholds of mangrove survival under rapid sea level rise. Science 2020, 368, 1118. [Google Scholar] [CrossRef] [PubMed]
- Törnqvist, T.E.; Cahoon, D.R.; Morris, J.T.; Day, J.W. Coastal Wetland Resilience, Accelerated Sea-Level Rise, and the Importance of Timescale. AGU Adv. 2021, 2, e2020AV000334. [Google Scholar] [CrossRef]
- Matthews, H.D.; Wynes, S. Current global efforts are insufficient to limit warming to 1.5 °C. Science 2022, 376, 1404–1409. [Google Scholar] [CrossRef]
- Groves, D.; Panis, T.; Wilson, M. 2023 Coastal Master Plan: Planning Tool Overview. Version I 2021, 51. Available online: https://coastal.la.gov/wp-content/uploads/2021/12/PlanningToolOverview__Oct2021.pdf (accessed on 17 August 2022).
- LoSchiavo, A.J.; Best, R.G.; Burns, R.E.; Gray, S.; Harwell, M.C.; Hines, E.B.; McLean, A.R.; St. Clair, T.; Traxler, S.; Vearil, J.W. Lessons Learned from the First Decade of Adaptive Management in Comprehensive Everglades Restoration. Ecol. Soc. 2013, 18, 4. [Google Scholar] [CrossRef]
- Farley, S.S.; Dawson, A.; Goring, S.J.; Williams, J.W. Situating Ecology as a Big-Data Science: Current Advances, Challenges, and Solutions. BioScience 2018, 68, 563–576. [Google Scholar] [CrossRef]
- Ladeau, S.L.; Han, B.A.; Rosi-Marshall, E.J.; Weathers, K.C. The Next Decade of Big Data in Ecosystem Science. Ecosystems 2017, 20, 274–283. [Google Scholar] [CrossRef] [Green Version]
- Simard, M.; Fatoyinbo, L.; Smetanka, C.; Rivera-Monroy, V.H.; Castaneda-Moya, E.; Thomas, N.; Van der Stocken, T. Mangrove canopy height globally related to precipitation, temperature and cyclone frequency. Nat. Geosci. 2019, 12, 40. [Google Scholar] [CrossRef] [Green Version]
- Adame, M.F.; Connolly, R.M.; Turschwell, M.P.; Lovelock, C.E.; Fatoyinbo, T.; Lagomasino, D.; Goldberg, L.A.; Holdorf, J.; Friess, D.A.; Sasmito, S.D.; et al. Future carbon emissions from global mangrove forest loss. Glob. Chang. Biol. 2021, 27, 2856–2866. [Google Scholar] [CrossRef] [PubMed]
- Sanderman, J.; Hengl, T.; Fiske, G.; Solvik, K.; Adame, M.F.; Benson, L.; Bukoski, J.J.; Carnell, P.; Cifuentes-Jara, M.; Donato, D.; et al. A global map of mangrove forest soil carbon at 30 m spatial resolution. Environ. Res. Lett. 2018, 13, 055002. [Google Scholar] [CrossRef]
- Lugo, A.E.; Medina, E.; McGinley, K. Issues and challenges of Mangrove conservation in the Anthropocene. Madera y Bosques 2014, 20, 11–38. [Google Scholar] [CrossRef] [Green Version]
- De Lacerda, L.D.; Ferreira, A.C.; Ward, R.; Borges, R. Editorial: Mangroves in the Anthropocene: From local change to global challenge. Front. For. Glob. Chang. 2022, 5. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Type | Sub-Type | Principle | Features |
|---|---|---|---|
| Conceptual model | • Conceptual models use a list of state variables and forcing functions in an ecosystem to describe and illustrate the interactions among state variables and associated processes. | • Since this tool is created to present the abstraction of reality in a system, these processes can be deployed at a relatively higher complexity level regardless of the final model version/development. | |
| • In some cases, prior knowledge and research products are included in the conceptual model and diagram; the model can be directly used to translate science concepts to management issues for management decisions and to communicate with government agencies and the public. | |||
| Agent-based model (ABM) | • ABM tracks the life history of individual trees (tree establishment, growth, and mortality). | • ABM captures the emergent forest properties and spatial patterns at the population level based on species-specific tree properties. • ABM requires larger computation resources as the selected modeling spatial scale increases. • ABMs can assess mangrove forest structure and carbon storage with a spatially explicit model implementation. | |
| Process-based model (PBM) | Hydrodynamic model | • PBM calculates the water dynamic movements by solving differential equations over time and space. • PBM highlights hydrodynamic processes that depict drivers and state variables that control ground surface materials flux and interactions with adjacent ecosystems (e.g., freshwater marsh, salt marsh, and estuary) | • PBM incorporates mass and momentum conservation as well as drag force and the eddy viscosity caused by complex mangrove tree structure; thus, it underscores the vegetation influence on water movements. • PBMs are commonly applied to assess the role of mangrove wetlands in coastal protection, especially coastlines threatened by sea-level rise and other extreme weather/climate events (e.g., tropical cyclones and tsunamis) |
| Hydrological model | • Hydrological and mass-balance models are considered two different types, yet both share common attributes of the mass-balance approach that ignore specific dynamic momentum of water movement across the system boundaries. • These models only consider the differences in the amount of material at the initial and at the end of a simulation time step. | • Models focus on water and material exchanges driven by system physical processes, such as water level, salinity, nutrient, and others (e.g., propagule dispersion). • Models can be used to investigate the impacts of SLR and flooding in coastal regions. | |
| Mass-balance (“Box”) model | • Models highlight energy or materials transport or exchange between mangrove wetlands and their adjacent system by considering the mangroves as a closed “box”. • Models are commonly used to assess carbon and energy fluxes across mangrove forests. | ||
| Climate model | • Global/regional climate model outputs/products are used in mangrove modeling research in either a direct or indirect way. | • The climate model output can be used as initial boundary conditions or as an independent input to initialize other models. • Some studies directly analyzed the climate model output to inform mangrove R/R activities. | |
| Statistical model | • The statistical model is an empirical model based on observation/sampling datasets. • Models include traditional statical analysis (i.e., regression and principal component analysis) and advanced machine learning (e.g., Structure Equation model and Artificial Neural Network). | • This model is the most widely used, especially regression, not only in mangrove research but also in other ecological studies. • Statistical models are widely applied to assess forest distribution, defining allometric relationships (structure, carbon) and other ecological interactions. This model can be used directly to provide management alternatives. | |
| Spatial model | GIS-based model | • Model mainly uses remotely sensing products. | • The spatial model largely depends on the Geographical Information System (GIS) and other geographical and spatial surveys. • Models are commonly used to investigate the spatial, and sometimes temporal, changes in mangrove distribution, extent, and some other structural attributes (e.g., biomass). • Models are used to map mangrove carbon and nutrient spatial distribution. • Models are applied to a wide range of spatial scales, from local to regional and global scales. |
| DEM-based model | • Models involving the application of digital elevation model (DEM) products. | ||
| Statistics-based model | • This group uses some other spatial or statistical analysis techniques (e.g., Kriging). | ||
| Socioeconomic/management model (ScoEco) | ScoEco-ES | • ScoEco models produce comprehensive evaluations based on statistical analysis, expert-based knowledge, and other survey methods (questionnaire and interview) aiming to guide and support decision-making and to plan mangrove R/R projects. | • Human activities and interferences are usually considered or summarized as statistical indexes to represent the human disturbance level. • Models are developed to evaluate the value of mangrove ecosystem services (ESs) to inform management plans by state/federal agencies. |
| ScoEco-management |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivera-Monroy, V.H.; Zhao, X.; Wang, H.; Xue, Z.G. Are Existing Modeling Tools Useful to Evaluate Outcomes in Mangrove Restoration and Rehabilitation Projects? A Minireview. Forests 2022, 13, 1638. https://doi.org/10.3390/f13101638
Rivera-Monroy VH, Zhao X, Wang H, Xue ZG. Are Existing Modeling Tools Useful to Evaluate Outcomes in Mangrove Restoration and Rehabilitation Projects? A Minireview. Forests. 2022; 13(10):1638. https://doi.org/10.3390/f13101638
Chicago/Turabian StyleRivera-Monroy, Victor H., Xiaochen Zhao, Hongqing Wang, and Zuo George Xue. 2022. "Are Existing Modeling Tools Useful to Evaluate Outcomes in Mangrove Restoration and Rehabilitation Projects? A Minireview" Forests 13, no. 10: 1638. https://doi.org/10.3390/f13101638
APA StyleRivera-Monroy, V. H., Zhao, X., Wang, H., & Xue, Z. G. (2022). Are Existing Modeling Tools Useful to Evaluate Outcomes in Mangrove Restoration and Rehabilitation Projects? A Minireview. Forests, 13(10), 1638. https://doi.org/10.3390/f13101638