

Environmental Factors Driving the Transpiration of a Betula platyphylla Sukaczev Forest in a Semi-arid Region in North China during Different Hydrological Years
 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
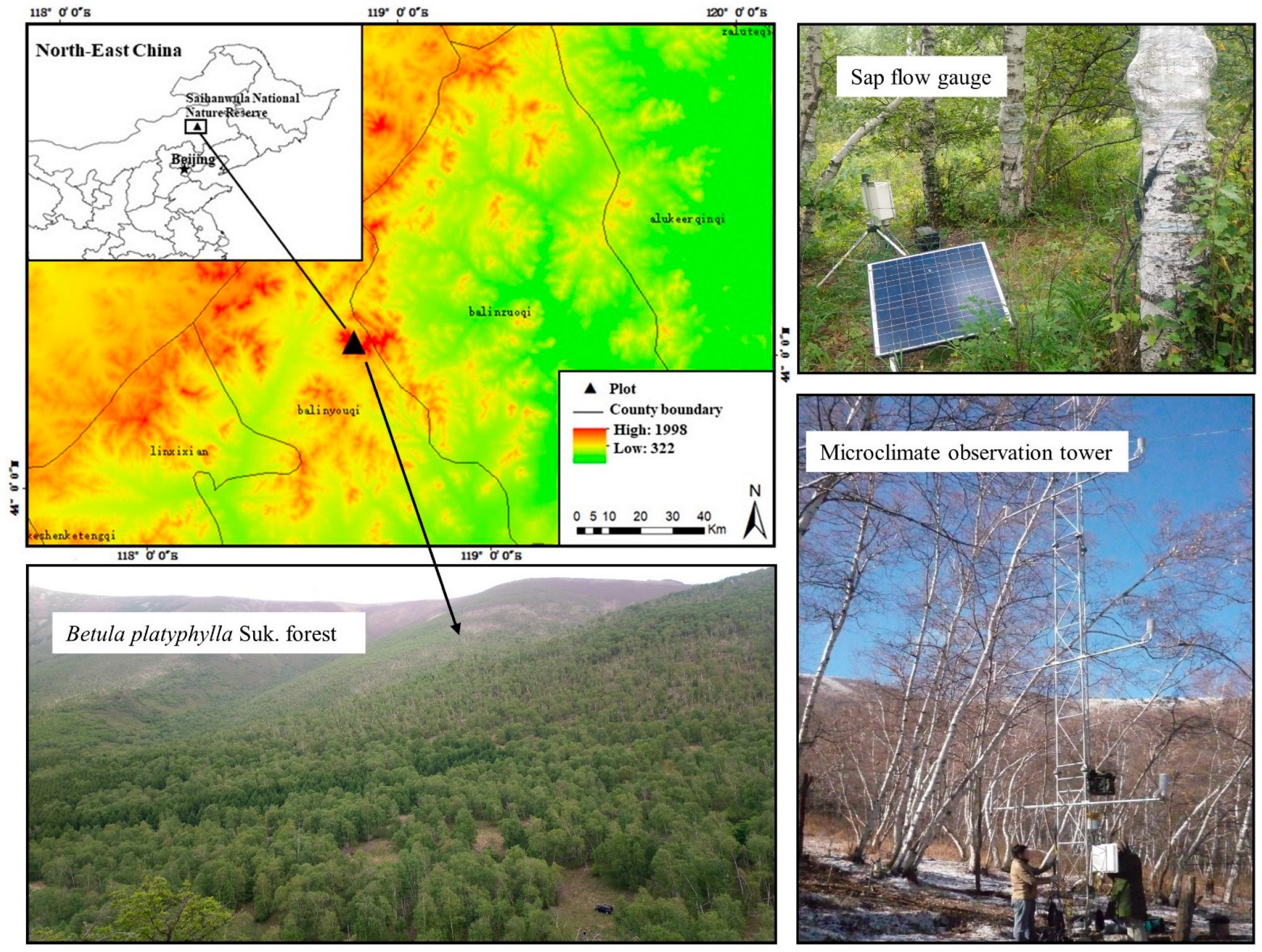
2.1. Study Area and Sample Trees
2.2. Sapwood Area Measurement
2.3. Division of Hydrological Years
2.4. Meteorological Measurements and Soil Moisture
2.5. Sap Flow Estimation Methods
2.5.1. Sap Flux Density
2.5.2. Tree Transpiration Calculation
2.5.3. Stand Transpiration Calculation
2.6. Model Structure and Statistical Assessment
2.7. Determination of the Contributions of , and to
3. Results
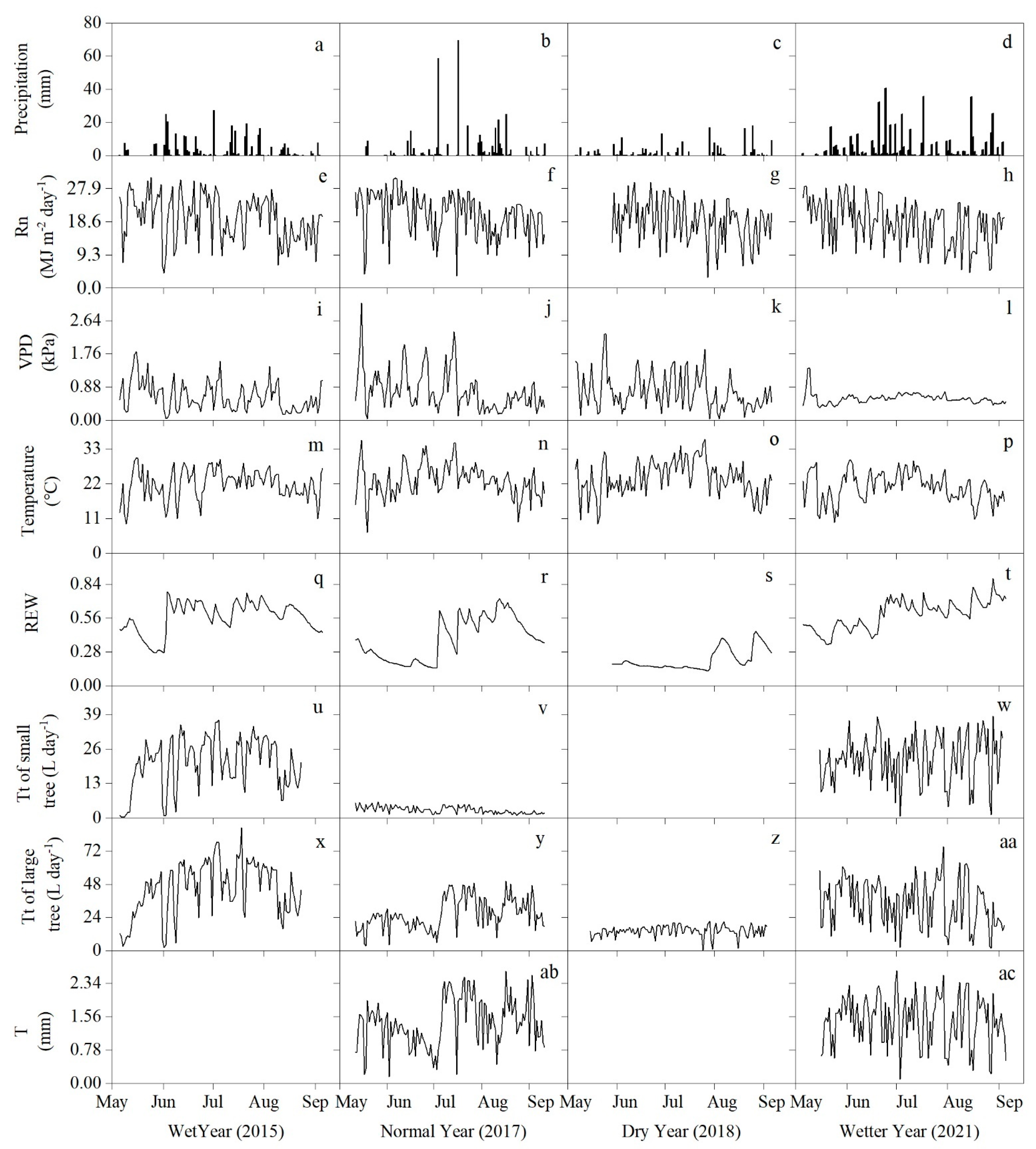
3.1. Environmental Characteristics of the Growing Season in Different Hydrological Years
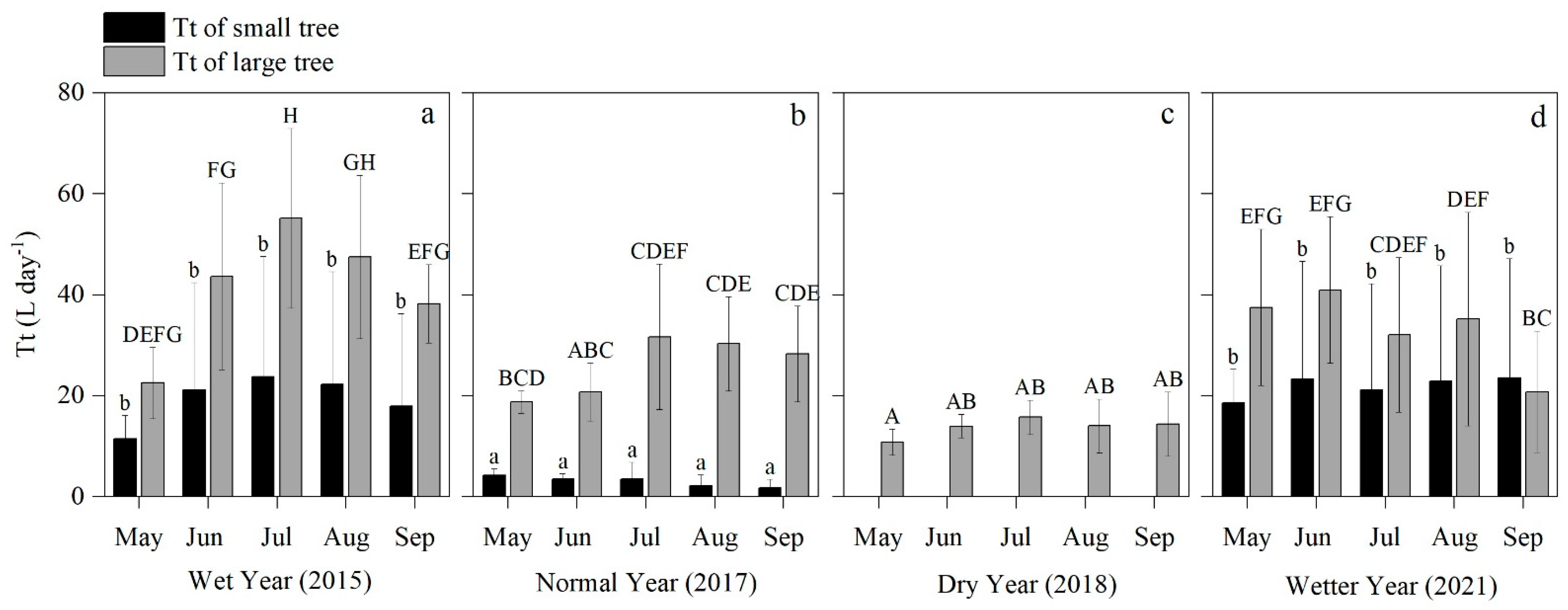
3.2. Variation of Tt and T in Different Hydrological Years
3.3. Response of T to ,
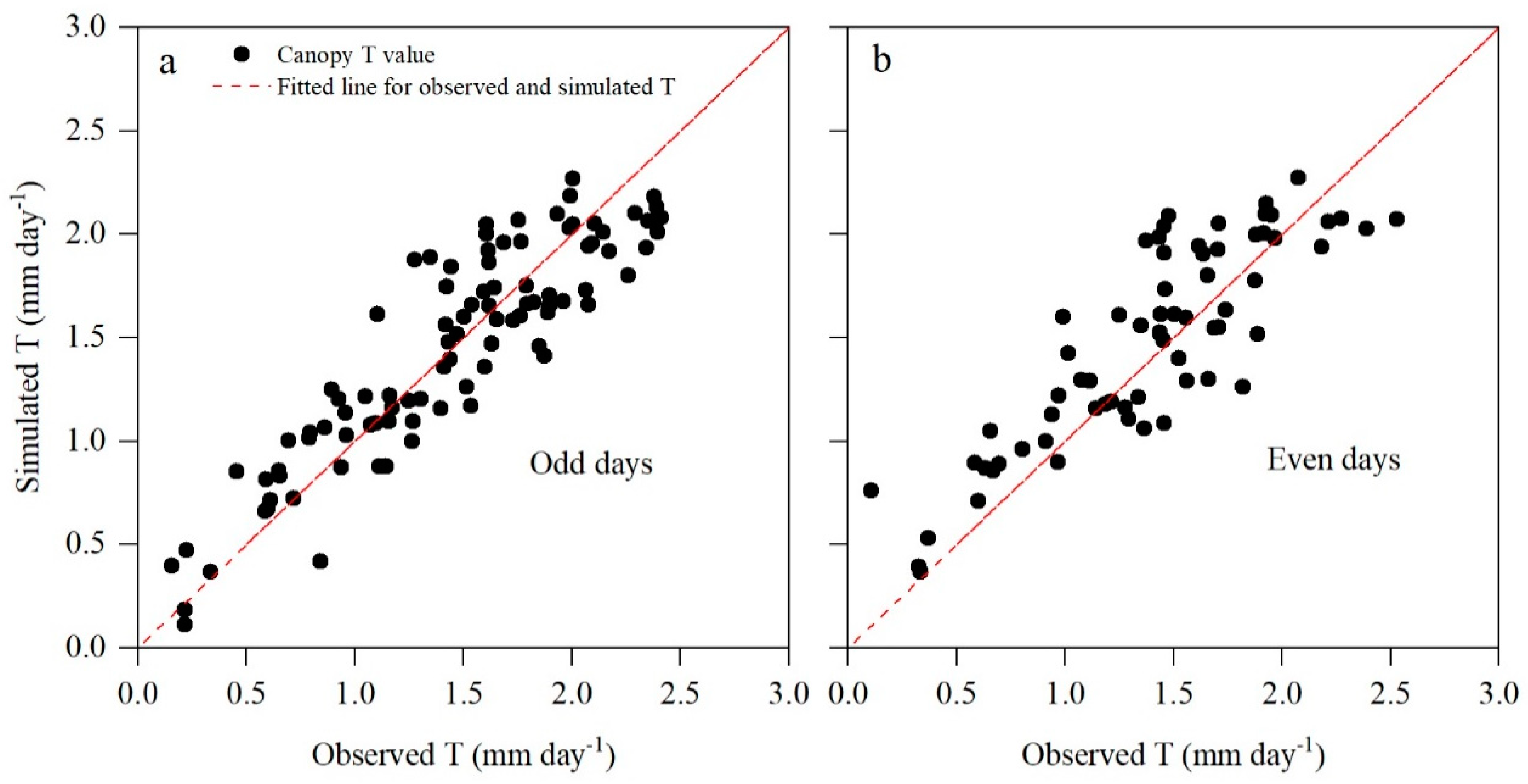
3.4. Construction and Validation of the T Model
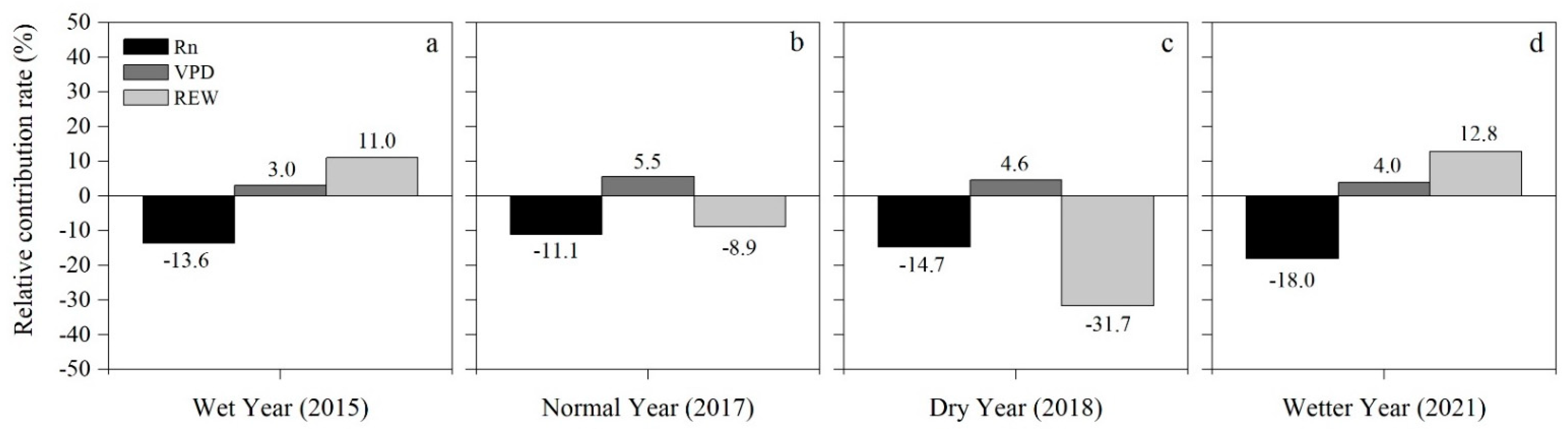
3.5. Relative Effects of and on T
4. Discussion
4.1. Effects of Drought on Transpiration
4.2. Assessing T Responses to Different Hydrological Years
4.3. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clark, J.S.; Iverson, L.; Woodall, C.W.; Allen, C.D.; Bell, D.M.; Bragg, D.C.; D’Amato, A.W.; Davis, F.W.; Hersh, M.H.; Ibanez, I.; et al. The impacts of increasing drought on forest dynamics, structure, and biodiversity in the United States. Glob. Chang. Biol. 2016, 22, 2329–2352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weed, A.S.; Ayres, M.P.; Hicke, J.A. Consequences of climate change for biotic disturbances in North American forests. Ecol. Monogr. 2013, 83, 441–470. [Google Scholar] [CrossRef]
- Adams, H.D.; Macalady, A.; Breshears, D.D.; Allen, C.D. Climate-induced tree mortality: Earth system consequences for carbon, energy, and water exchanges. AGU Fall Meet. Abstr. 2010, 91, 153–154. [Google Scholar] [CrossRef]
- Boisvenue, C.; Running, S.W. Impacts of climate change on natural forest productivity—Evidence since the middle of the 20th century. Glob. Chang. Biol. 2006, 12, 862–882. [Google Scholar] [CrossRef]
- Zeng, N.; Yao, H.; Zhou, M.; Zhao, P.; Dech, J.P.; Zhang, B.; Lu, X. Species-specific determinants of mortality and recruitment in the forest-steppe ecotone of northeast China. For. Chron. 2016, 92, 336–344. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Liu, H.; Hampe, A. Hydraulic adaptability promotes tree life spans under climate dryness. Glob. Ecol. Biogeogr. 2021, 31, 51–61. [Google Scholar] [CrossRef]
- Chang, X.; Zhao, W.; Liu, H.; Wei, X.; Liu, B.; He, Z. Qinghai spruce (Picea crassifolia) forest transpiration and canopy conductance in the upper Heihe River Basin of arid northwestern China. Agric. For. Meteorol. 2014, 198–199, 209–220. [Google Scholar] [CrossRef]
- Ungar, E.D.; Rotenberg, E.; Raz-Yaseef, N.; Cohen, S.; Yakir, D.; Schiller, G. Transpiration and annual water balance of Aleppo pine in a semiarid region: Implications for forest management. For. Ecol. Manag. 2013, 298, 39–51. [Google Scholar] [CrossRef]
- Forner, A.; Aranda, I.; Granier, A.; Valladares, F. Differential impact of the most extreme drought event over the last half century on growth and sap flow in two coexisting Mediterranean trees. Plant Ecol. 2014, 215, 703–719. [Google Scholar] [CrossRef] [Green Version]
- Granier, A.; Biron, P.; Lemoine, D.G. Water balance, transpiration and canopy conductance in two beech stands. Agric. For. Meteorol. 2000, 100, 291–308. [Google Scholar] [CrossRef]
- Steppe, K.; Vandegehuchte, M.W.; Tognetti, R.; Mencuccini, M. Sap flow as a key trait in the understanding of plant hydraulic functioning. Tree Physiol 2015, 35, 341–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tie, Q.; Hu, H.; Tian, F.; Guan, H.; Lin, H. Environmental and physiological controls on sap flow in a subhumid mountainous catchment in North China. Agric. For. Meteorol. 2017, 240–241, 46–57. [Google Scholar] [CrossRef]
- Kunert, N.; Schwendenmann, L.; Hölscher, D. Seasonal dynamics of tree sap flux and water use in nine species in Panamanian forest plantations. Agric. For. Meteorol. 2010, 150, 411–419. [Google Scholar] [CrossRef]
- Granier, A.; Bréda, N. Modelling canopy conductance and stand transpiration of an oak forest from sap flow measurements. Ann. Des Sci. For. 1996, 53, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Phillips, N.G.; Ryan, M.G.; Bond, B.J.; McDowell, N.G.; Hinckley, T.M.; Cermak, J. Reliance on stored water increases with tree size in three species in the Pacific Northwest. Tree Physiol. 2003, 23, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Ewers, B.E.; Mackay, D.S.; Tang, J.; Bolstad, P.V.; Samanta, S. Intercomparison of sugar maple (Acer saccharum Marsh.) stand transpiration responses to environmental conditions from the Western Great Lakes Region of the United States. Agric. For. Meteorol. 2008, 148, 231–246. [Google Scholar] [CrossRef]
- Bretfeld, M.; Ewers, B.E.; Hall, J.S. Plant water use responses along secondary forest succession during the 2015–2016 El Nino drought in Panama. New Phytol. 2018, 219, 885–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, S.; Wang, Y.-L.; Kume, T.; Zhang, J.-G.; Otsuki, K.; Yamanaka, N.; Liu, G.-B. Sapflow characteristics and climatic responses in three forest species in the semiarid Loess Plateau region of China. Agric. For. Meteorol. 2011, 151, 1–10. [Google Scholar] [CrossRef]
- Oogathoo, S.; Houle, D.; Duchesne, L.; Kneeshaw, D. Vapour pressure deficit and solar radiation are the major drivers of transpiration of balsam fir and black spruce tree species in humid boreal regions, even during a short-term drought. Agric. For. Meteorol. 2020, 291, 108063. [Google Scholar] [CrossRef]
- Llorens, P.; Poyatos, R.; Latron, J.; Delgado, J.; Oliveras, I.; Gallart, F. A multi-year study of rainfall and soil water controls on Scots pine transpiration under Mediterranean mountain conditions. Hydrol. Process. 2010, 24, 3053–3064. [Google Scholar] [CrossRef]
- Mitchell, P.J.; Benyon, R.G.; Lane, P.N.J. Responses of evapotranspiration at different topographic positions and catchment water balance following a pronounced drought in a mixed species eucalypt forest, Australia. J. Hydrol. 2012, 440–441, 62–74. [Google Scholar] [CrossRef]
- da Silva Sallo, F.; Sanches, L.; de Morais Dias, V.R.; da Silva Palácios, R.; de Souza Nogueira, J. Stem water storage dynamics of Vochysia divergens in a seasonally flooded environment. Agric. For. Meteorol. 2017, 232, 566–575. [Google Scholar] [CrossRef]
- Wang, H.; Tetzlaff, D.; Dick, J.J.; Soulsby, C. Assessing the environmental controls on Scots pine transpiration and the implications for water partitioning in a boreal headwater catchment. Agric. For. Meteorol. 2017, 240–241, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Lagergren, F.; Lindroth, A. Transpiration response to soil moisture in pine and spruce trees in Sweden. Agric. For. Meteorol. 2002, 112, 67–85. [Google Scholar] [CrossRef]
- Granier, A.; Breda, N.; Biron, P.; Villette, S. A lumped water balance model to evaluate duration and intensity of drought constraints in forest stands. Ecol. Model. 1999, 116, 269–283. [Google Scholar] [CrossRef]
- Al-Yahyai, R.; Schaffer, B.; Davies, F.S. Physiological Response of Carambola Trees to Soil Water Depletion in Krome Soils. HortScience 2004, 39, 857B-857. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Yu, P.; Wang, Y.; Wang, B.; Yu, Y.; Wang, X.; Liu, Z.; Liu, X.; Wang, S.; Xiong, W. The Variation in Water Consumption by Transpiration of Qinghai Spruce among Canopy Layers in the Qilian Mountains, Northwestern China. Forests 2020, 11, 845. [Google Scholar] [CrossRef]
- Li, Z.; Yu, P.; Wang, Y.; Webb, A.A.; He, C.; Wang, Y.; Yang, L. A model coupling the effects of soil moisture and potential evaporation on the tree transpiration of a semi-arid larch plantation. Ecohydrology 2016, 10, e1764. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Wang, S.; Yuan, G.; Yang, Y.; Cao, M. Evapotranspiration of a tropical rain forest in Xishuangbanna, southwest China. Hydrol. Process. 2010, 24, 2405–2416. [Google Scholar] [CrossRef]
- Huang, J.; Zhou, Y.; Yin, L.; Wenninger, J.; Zhang, J.; Hou, G.; Zhang, E.; Uhlenbrook, S. Climatic controls on sap flow dynamics and used water sources of Salix psammophila in a semi-arid environment in northwest China. Environ. Earth Sci. 2014, 73, 289–301. [Google Scholar] [CrossRef]
- Thomas, F.M.; Foetzki, A.; Arndt, S.K.; Bruelheide, H.; Gries, D.; Li, X.; Zeng, F.; Zhang, X.; Runge, M. Water use by perennial plants in the transition zone between river oasis and desert in NW China. Basic Appl. Ecol. 2006, 7, 253–267. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, Y.; Tian, A.; Webb, A.A.; Yu, P.; Xiong, W.; Xu, L.; Wang, Y. Modeling the Response of Daily Evapotranspiration and its Components of a Larch Plantation to the Variation of Weather, Soil Moisture, and Canopy Leaf Area Index. J. Geophys. Res. Atmos. 2018, 123, 7354–7374. [Google Scholar] [CrossRef]
- Asbjornsen, H.; Campbell, J.L.; Jennings, K.A.; Vadeboncoeur, M.A.; McIntire, C.; Templer, P.H.; Phillips, R.P.; Bauerle, T.L.; Dietze, M.C.; Frey, S.D.; et al. Guidelines and considerations for designing field experiments simulating precipitation extremes in forest ecosystems. Methods Ecol. Evol. 2018, 9, 2310–2325. [Google Scholar] [CrossRef] [Green Version]
- Vilhar, U. Comparison of drought stress indices in beech forests: A modelling study. Iforest Biogeosci. For. 2016, 9, 635–642. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xiong, W.; Yu, P.; Shen, Z.; Guo, M.; Guan, W.; Ma, C.; Ye, B.; Guo, H. Study on the evapotranspiration of forest and vegetation in dryland. Sci. Soil Water Conserv. 2006, 4, 19–26. [Google Scholar] [CrossRef]
- Whitley, R.; Zeppel, M.; Armstrong, N.; Macinnis-Ng, C.; Yunusa, I.; Eamus, D. A modified Jarvis-Stewart model for predicting stand-scale transpiration of an Australian native forest. Plant Soil 2007, 305, 35–47. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, P.G. The interpretation of the variations in leaf water potential and stomatal conductance found in canopies in the field. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1997, 273, 593–610. [Google Scholar] [CrossRef]
- Yu, S.; Guo, J.; Liu, Z.; Wang, Y.; Ma, J.; Li, J.; Liu, F. Assessing the Impact of Soil Moisture on Canopy Transpiration Using a Modified Jarvis-Stewart Model. Water 2021, 13, 2720. [Google Scholar] [CrossRef]
- Petzold, R.; Schwärzel, K.; Feger, K.-H. Transpiration of a hybrid poplar plantation in Saxony (Germany) in response to climate and soil conditions. Eur. J. For. Res. 2010, 130, 695–706. [Google Scholar] [CrossRef]
- Li, H. Under Saihanwula Nature Reserve Different Vegetation Types in the Comparative Study of Soil Physical and Chemical Properties. Master’s Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2010. [Google Scholar]
- Campbell, G.S.; Norman, J. An Introduction to Environmental Biophysics; Springer: New York, NY, USA, 1998. [Google Scholar]
- Qin, J. Drought and Salinity Resistant Physiology and Water Consumption Characteristics of Main Plantation Tree Species in High-Cold Region of Loess Plateau; Beijing Forestry University: Beijing, China, 2011. [Google Scholar]
- Granier, A. Une nouvelle methode pour la mesure dy flux de seve brute dans le trons des arbres. Ann. Sci. 1985, 22, 193–200. [Google Scholar] [CrossRef]
- Samuelson, L.J.; Stokes, T.A.; Ramirez, M.R.; Mendonca, C.C. Drought tolerance of a Pinus palustris plantation. For. Ecol. Manag. 2019, 451, 117557. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Z.; Guo, J.; Wang, Y.; Ma, J.; Yu, S.; Yu, P.; Xu, L. Estimate canopy transpiration in larch plantations via the interactions among reference evapotranspiration, leaf area index, and soil moisture. For. Ecol. Manag. 2021, 481, 118749. [Google Scholar] [CrossRef]
- Granier, A.; Loustau, D.; Bréda, N. A generic model of forest canopy conductance dependent on climate, soil water availability and leaf area index. Ann. For. Sci. 2000, 57, 755–765. [Google Scholar] [CrossRef]
- Loritz, R.; Bassiouni, M.; Hildebrandt, A.; Hassler, S.K.; Zehe, E. Leveraging sap flow data in a catchment-scale hybrid model to improve soil moisture and transpiration estimates. Hydrol. Earth Syst. Sci. 2022, 26, 4757–4771. [Google Scholar] [CrossRef]
- She, D.; Xia, Y.; Shao, M.; Peng, S.; Yu, S. Transpiration and canopy conductance of Caragana korshinskii trees in response to soil moisture in sand land of China. Agrofor. Syst. 2012, 87, 667–678. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, Y.; Wang, Y.; Guo, J.; Yu, P.; Wang, L.; Yu, S.; Liu, F. Separating meteorological condition and soil moisture controls on the variation in stand evapotranspiration of a larch plantation during three hydrological years. Glob. Ecol. Conserv. 2021, 27, e01548. [Google Scholar] [CrossRef]
- Schmidt, U.; Thöni, H.; Kaupenjohann, M. Using a boundary line approach to analyze N2O flux data from agricultural soils. Nutr. Cycl. Agroecosyst. 2000, 57, 119–129. [Google Scholar] [CrossRef]
- Ma, J.; Guo, J.; Liu, Z.; Wang, Y.; Zhang, Z. Diurnal variations of stand transpiration of Larix principis-rupprechtii forest and its response to environmental factors in Liupan Mountains of northwestern China. J. Beijing For. Univ. 2020, 42, 1–11. [Google Scholar] [CrossRef]
- Wilby, R.L. Uncertainty in water resource model parameters used for climate change impact assessment. Hydrol. Process. 2005, 19, 3201–3219. [Google Scholar] [CrossRef]
- Mitchell, P.L. Misuse of regression for empirical validation of models. Agric. Syst. 1997, 54, 313–326. [Google Scholar] [CrossRef]
- Rudiyanto; Minasny, B.; Setiawan, B.I.; Saptomo, S.K.; McBratney, A.B. Open digital mapping as a cost-effective method for mapping peat thickness and assessing the carbon stock of tropical peatlands. Geoderma 2018, 313, 25–40. [Google Scholar] [CrossRef]
- Whitley, R.; Taylor, D.; Macinnis-Ng, C.; Zeppel, M.; Yunusa, I.; O’Grady, A.; Froend, R.; Medlyn, B.; Eamus, D. Developing an empirical model of canopy water flux describing the common response of transpiration to solar radiation and VPD across five contrasting woodlands and forests. Hydrol. Process. 2013, 27, 1133–1146. [Google Scholar] [CrossRef]
- Zhang, K.; Kimball, J.S.; Nemani, R.R.; Running, S.W.; Hong, Y.; Gourley, J.J.; Yu, Z. Vegetation Greening and Climate Change Promote Multidecadal Rises of Global Land Evapotranspiration. Sci. Rep. 2015, 5, 15956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knutti, R.; Rogelj, J.; Plattner, G.-K.; Sedláček, J.; Allen, S.K.; Stocker, T.F. IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar] [CrossRef] [Green Version]
- Duan, H.; Duursma, R.A.; Huang, G.; Smith, R.A.; Choat, B.; O’Grady, A.P.; Tissue, D.T. Elevated [CO2] does not ameliorate the negative effects of elevated temperature on drought-induced mortality in Eucalyptus radiata seedlings. Plant Cell Environ. 2014, 37, 1598–1613. [Google Scholar] [CrossRef] [PubMed]
- Adams, H.D.; Guardiola, M.; Barron-Gafford, G.; Villegas, J.C.; Breshears, D.D.; Zou, C.B.; Troch, P.A.; Huxman, T.E. Temperature sensitivity of drought-induced tree mortality portends increased regional die-off under global-change-type drought. Proc. Natl. Acad. Sci. USA 2009, 106, 7063–7066. [Google Scholar] [CrossRef] [Green Version]
- David, T.S.; Henriques, M.O.; Kurz-Besson, C.; Nunes, J.; Valente, F.; Vaz, M.; Pereira, J.S.; Siegwolf, R.; Chaves, M.M.; Gazarini, L.C.; et al. Water-use strategies in two co-occurring Mediterranean evergreen oaks: Surviving the summer drought. Tree Physiol. 2007, 27, 793–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawson, T. Determining water use by trees and forests from isotopic, energy balance and transpiration analyses: The roles of tree size and hydraulic lift. Tree Physiol. 1996, 16, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Meinzer, F.C.; Andrade, J.L.; Goldstein, G.; Holbrook, N.M.; Cavelier, J.; Wright, S.J. Partitioning of soil water among canopy trees in a seasonally dry tropical forest. Oecologia 1999, 121, 293–301. [Google Scholar] [CrossRef]
- Sinacore, K.; Asbjornsen, H.; Hernandez-Santana, V.; Hall, J.S. Differential and dynamic water regulation responses to El Niño for monospecific and mixed species planted forests. Ecohydrology 2020, 13, e2238. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, Z.; Li, Z.; Tang, J.; Caldwell, P.; Zhang, W. Biophysical control of whole tree transpiration under an urban environment in Northern China. J. Hydrol. 2011, 402, 388–400. [Google Scholar] [CrossRef]
- Han, C.; Chen, N.; Zhang, C.; Liu, Y.; Khan, S.; Lu, K.; Li, Y.; Dong, X.; Zhao, C. Sap flow and responses to meteorological about the Larix principis-rupprechtii plantation in Gansu Xinlong mountain, northwestern China. For. Ecol. Manag. 2019, 451, 117519. [Google Scholar] [CrossRef]
- Fried, J.S.; Torn, M.S.; Mills, E. The Impact of Climate Change on Wildfire Severity: A Regional Forecast for Northern California. Clim. Chang. 2004, 64, 169–191. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Sample Tree No. | Tree Height (m) | Diameter at Breast Height (cm) | Sapwood Thickness (cm) | Sapwood Area (cm2) |
|---|---|---|---|---|---|
| 2015, 2017 | 1 | 14.5 | 15 | 3.06 | 85.06 |
| 2 | 13.6 | 17.6 | 3.67 | 121.58 | |
| 3 | 15 | 19.1 | 4.03 | 145.97 | |
| 4 | 15.2 | 21 | 4.49 | 180.44 | |
| 2018 | 5 | 11.3 | 14.1 | 2.85 | 74.07 |
| 6 | 15.1 | 17.4 | 3.62 | 118.52 | |
| 7 | 14.5 | 19.9 | 4.22 | 159.99 | |
| 8 | 16.2 | 21.3 | 4.56 | 186.25 | |
| 2021 | 9 | 15.8 | 14.3 | 2.90 | 76.44 |
| 10 | 13.2 | 15.5 | 3.17 | 91.53 | |
| 11 | 14 | 20.9 | 4.46 | 178.52 | |
| 12 | 15.6 | 22.7 | 4.90 | 214.73 |
| Year | Hydrological Years | Annual Precipitation (mm) | Percentage Difference from the Multi-Year Average |
|---|---|---|---|
| 2015 | Wet year | 445 | +18.5% |
| 2017 | Normal year | 394 | +5% |
| 2018 | Dry year | 223 | −40.7% |
| 2021 | Wetter year | 561 | +49.3% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Zhao, P.; Zhou, M.; Liu, Z.; Yao, H.; Wei, J.; Shu, Y.; Li, J.; Xiang, C.; Zhou, L. Environmental Factors Driving the Transpiration of a Betula platyphylla Sukaczev Forest in a Semi-arid Region in North China during Different Hydrological Years. Forests 2022, 13, 1729. https://doi.org/10.3390/f13101729
Wu Y, Zhao P, Zhou M, Liu Z, Yao H, Wei J, Shu Y, Li J, Xiang C, Zhou L. Environmental Factors Driving the Transpiration of a Betula platyphylla Sukaczev Forest in a Semi-arid Region in North China during Different Hydrological Years. Forests. 2022; 13(10):1729. https://doi.org/10.3390/f13101729
Chicago/Turabian StyleWu, Yiheng, Pengwu Zhao, Mei Zhou, Zebin Liu, Huaxia Yao, Jiangsheng Wei, Yang Shu, Jiamei Li, Changlin Xiang, and Liwen Zhou. 2022. "Environmental Factors Driving the Transpiration of a Betula platyphylla Sukaczev Forest in a Semi-arid Region in North China during Different Hydrological Years" Forests 13, no. 10: 1729. https://doi.org/10.3390/f13101729
APA StyleWu, Y., Zhao, P., Zhou, M., Liu, Z., Yao, H., Wei, J., Shu, Y., Li, J., Xiang, C., & Zhou, L. (2022). Environmental Factors Driving the Transpiration of a Betula platyphylla Sukaczev Forest in a Semi-arid Region in North China during Different Hydrological Years. Forests, 13(10), 1729. https://doi.org/10.3390/f13101729