
Trade-Offs between Economic Gains and Carbon Stocks across a Range of Management Alternatives in Boreal Forests


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Study Area and Chronosequence Data
2.2. Carbon Stocks
2.3. Forest Management Alternatives
2.4. Economic Gains
2.5. Statistical Trade-Off Analysis
3. Results
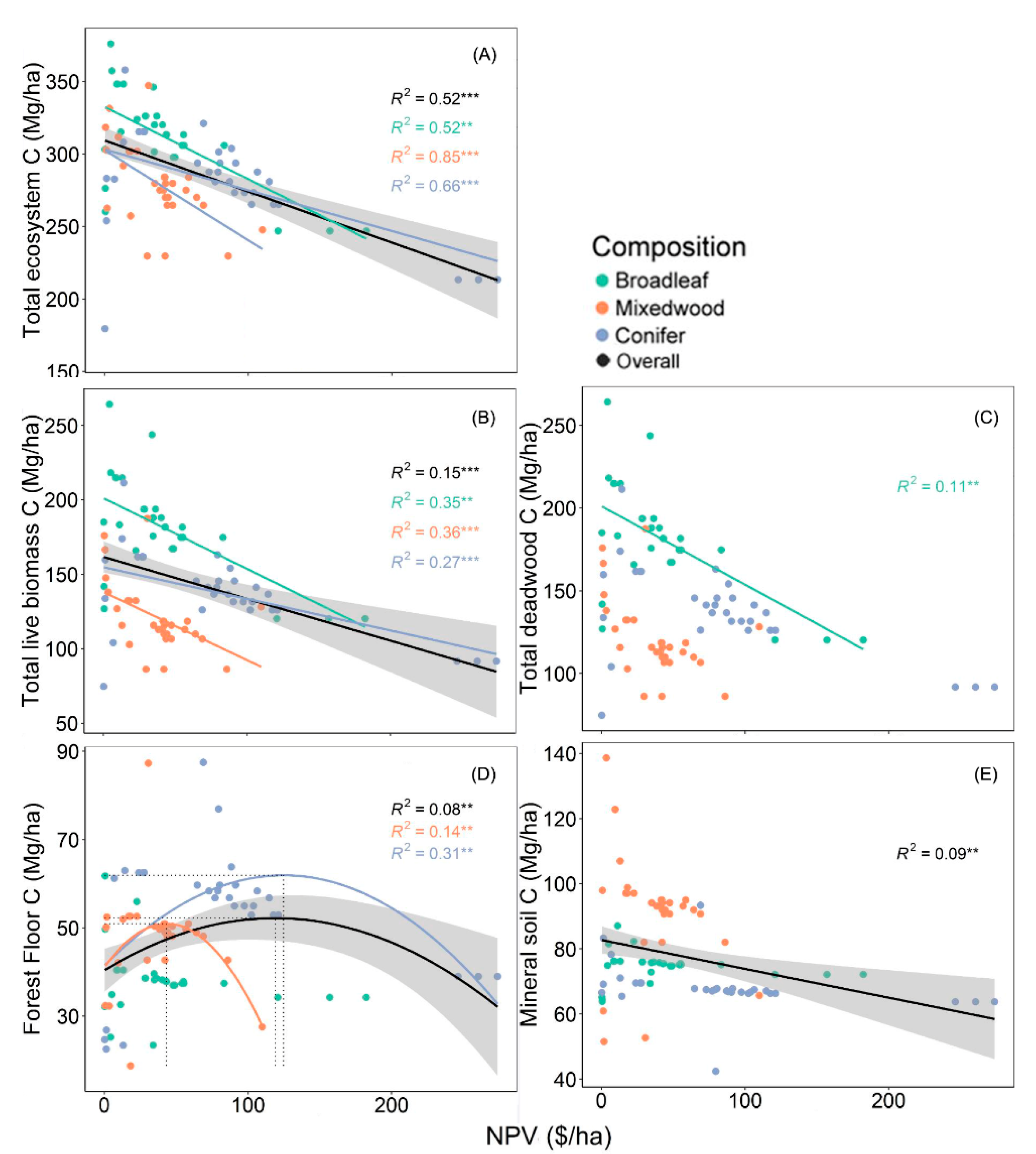
3.1. Relationship between Economic Gains and Carbon Stocks
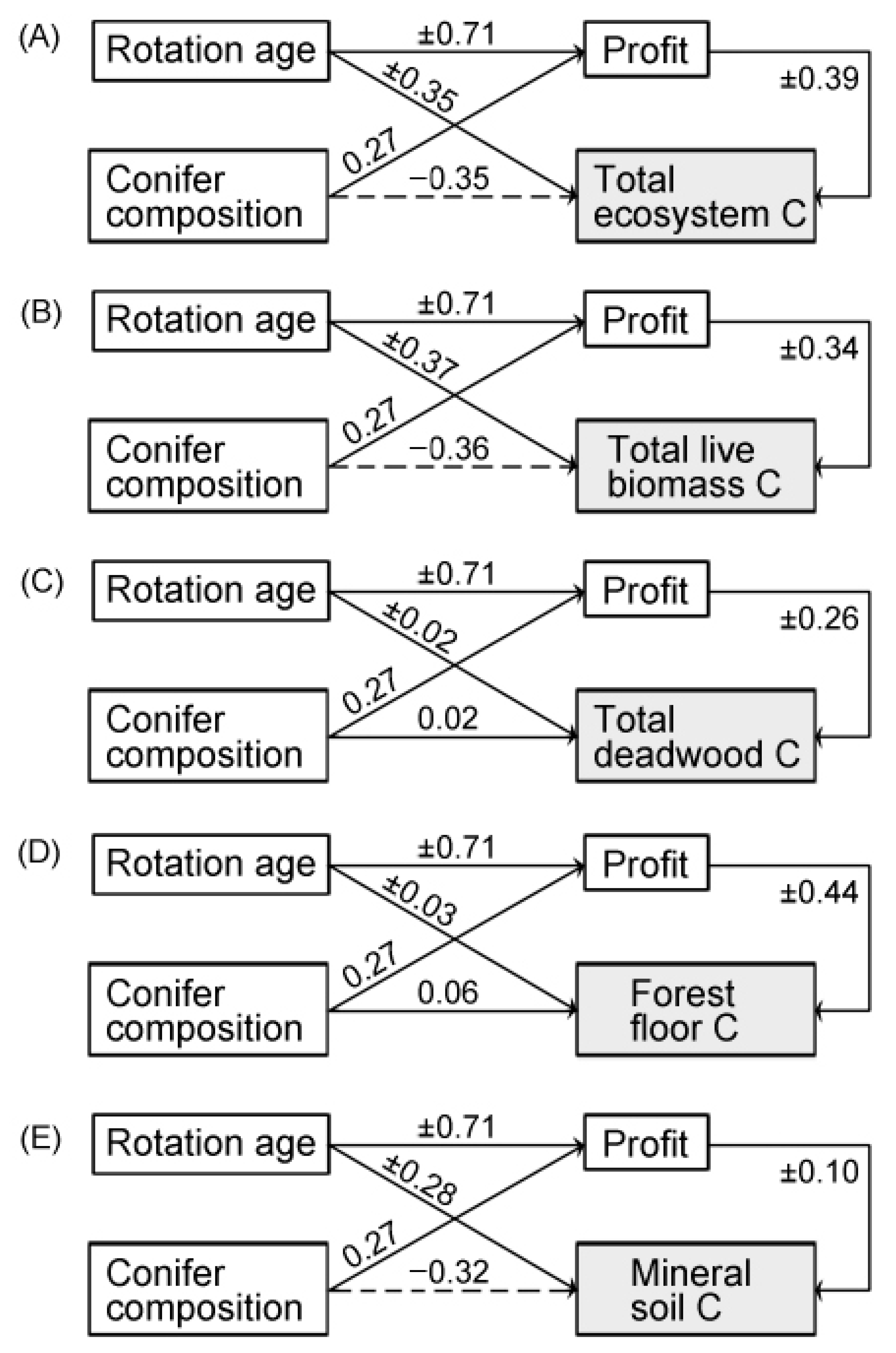
3.2. Effects of Forest Management Alternatives on Economic Gains and Carbon Stocks
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anonymous; Millennium Ecosystem Assessment. Ecosystems and Human Well-Being: Biodiversity Synthesis. Ecol. Manag. Restor. 2005, 6, 226–227. [Google Scholar]
- Duncker, P.S.; Raulund-Rasmussen, K.; Gundersen, P.; Katzensteiner, K.; De Jong, J.; Ravn, H.P.; Smith, M.; Eckmuellner, O.; Spiecker, H. How Forest Management Affects Ecosystem Services, Including Timber Production and Economic Return: Synergies and Trade-Offs. Ecol. Soc. 2012, 17, 50. [Google Scholar] [CrossRef] [Green Version]
- Houghton, R.A. Balancing the Global Carbon Budget. Annu. Rev. Earth Planet. Sci. 2007, 35, 313–347. [Google Scholar] [CrossRef] [Green Version]
- Trivino, M.; Juutinen, A.; Mazziotta, A.; Miettinen, K.; Podkopaev, D.; Reunanen, P.; Monkkonen, M. Managing a Boreal Forest Landscape for Providing Timber, Storing and Sequestering Carbon. Ecosyst. Serv. 2015, 14, 179–189. [Google Scholar] [CrossRef]
- Strengbom, J.; Axelsson, E.P.; Lundmark, T.; Nordin, A. Trade-Offs in the Multi-Use Potential of Managed Boreal Forests. J. Appl. Ecol. 2018, 55, 958–966. [Google Scholar] [CrossRef]
- Schwenk, W.S.; Donovan, T.M.; Keeton, W.S.; Nunery, J.S. Carbon Storage, Timber Production, and Biodiversity: Comparing Ecosystem Services with Multi-Criteria Decision Analysis. Ecol. Appl. 2012, 22, 1612–1627. [Google Scholar] [CrossRef]
- Kline, J.D.; Harmon, M.E.; Spies, T.A.; Morzillo, A.T.; Pabst, R.J.; McComb, B.C.; Schnekenburger, F.; Olsen, K.A.; Csuti, B.; Vogeler, J.C. Evaluating Carbon Storage, Timber Harvest, and Habitat Possibilities for a Western Cascades (USA) Forest Landscape. Ecol. Appl. 2016, 26, 2044–2059. [Google Scholar] [CrossRef]
- Kang, H.; Seely, B.; Wang, G.; Innes, J.; Zheng, D.; Chen, P.; Wang, T.; Li, Q. Evaluating Management Tradeoffs between Economic Fiber Production and Other Ecosystem Services in a Chinese-Fir Dominated Forest Plantation in Fujian Province. Sci. Total Environ. 2016, 557, 80–90. [Google Scholar] [CrossRef]
- Seidl, R.; Rammer, W.; Jaeger, D.; Currie, W.S.; Lexer, M.J. Assessing Trade-Offs between Carbon Sequestration and Timber Production within a Framework of Multi-Purpose Forestry in Austria. For. Ecol. Manag. 2007, 248, 64–79. [Google Scholar] [CrossRef]
- Rana, E.; Thwaites, R.; Luck, G. Trade-Offs and Synergies between Carbon, Forest Diversity and Forest Products in Nepal Community Forests. Environ. Conserv. 2017, 44, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Trivino, M.; Pohjanmies, T.; Mazziotta, A.; Juutinen, A.; Podkopaev, D.; Le Tortorec, E.; Monkkonen, M. Optimizing Management to Enhance Multifunctionality in a Boreal Forest Landscape. J. Appl. Ecol. 2017, 54, 61–70. [Google Scholar] [CrossRef]
- Lagergren, F.; Jonsson, A.M. Ecosystem Model Analysis of Multi-Use Forestry in a Changing Climate. Ecosyst. Serv. 2017, 26, 209–224. [Google Scholar] [CrossRef]
- Dixon, R.K.; Brown, S.; Houghton, R.A.; Solomon, A.M.; Trexler, M.C.; Wisniewski, J. Carbon Pools and Flux of Global Forest Ecosystems. Science 1994, 263, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Shahi, C.; Chen, H.Y.H.; McLaren, B. Economic Analysis of Forest Management Alternatives: Compositional Objectives, Rotation Ages, and Harvest Methods in Boreal Forests. For. Policy Econ. 2017, 85, 124–134. [Google Scholar] [CrossRef]
- Gao, B.; Taylor, A.R.; Searle, E.B.; Kumar, P.; Ma, Z.; Hume, A.M.; Chen, H.Y.H. Carbon Storage Declines in Old Boreal Forests Irrespective of Succession Pathway. Ecosystems 2018, 21, 1168–1182. [Google Scholar] [CrossRef]
- Liski, J.; Pussinen, A.; Pingoud, K.; MskipSs, R.; Karjalainen, T. Which Rotation Length Is Favourable to Carbon Sequestration? Can. J. For. Res. 2001, 31, 2004–2013. [Google Scholar] [CrossRef]
- Nakajima, T.; Shiraishi, N.; Kanomata, H.; Matsumoto, M. A Method to Maximise Forest Profitability through Optimal Rotation Period Selection under Various Economic, Site and Silvicultural Conditions. N. Z. J. For. Sci. 2017, 47, 4. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Chen, H.Y.H. Competition, Species Interaction and Ageing Control Tree Mortality in Boreal Forests. J. Ecol. 2011, 99, 1470–1480. [Google Scholar] [CrossRef]
- Seedre, M.; Chen, H.Y.H. Carbon Dynamics of Aboveground Live Vegetation of Boreal Mixedwoods after Wildfire and Clear-Cutting. Can. J. For. Res. 2010, 40, 1862–1869. [Google Scholar] [CrossRef]
- Laganiere, J.; Cavard, X.; Brassard, B.W.; Pare, D.; Bergeron, Y.; Chen, H.Y.H. The Influence of Boreal Tree Species Mixtures on Ecosystem Carbon Storage and Fluxes. For. Ecol. Manag. 2015, 354, 119–129. [Google Scholar] [CrossRef]
- Hart, S.A.; Chen, H.Y.H. Fire, Logging, and Overstory Affect Understory Abundance, Diversity, and Composition in Boreal Forest. Ecol. Monogr. 2008, 78, 123–140. [Google Scholar] [CrossRef]
- Ma, Z.; Chen, H.Y.H. Effects of Species Diversity on Fine Root Productivity Increase with Stand Development and Associated Mechanisms in a Boreal Forest. J. Ecol. 2017, 105, 237–245. [Google Scholar] [CrossRef]
- Ma, Z.; Chen, H.Y.H.; Kumar, P.; Gao, B. Species Mixture Increases Production Partitioning to Belowground in a Natural Boreal Forest. For. Ecol. Manag. 2019, 432, 667–674. [Google Scholar] [CrossRef]
- Environment Canada. Canadian Climate Normals 1981–2010. 2016. Available online: http://climate.weatheroffice.gc.ca/climate_normals/index_e.html (accessed on 1 February 2018).
- Senici, D.; Chen, H.Y.H.; Bergeron, Y.; Cyr, D. Spatiotemporal Variations of Fire Frequency in Central Boreal Forest. Ecosystems 2010, 13, 1227–1238. [Google Scholar] [CrossRef]
- Shrestha, B.M.; Chen, H.Y.H. Effects of Stand Age, Wildfire and Clearcut Harvesting on Forest Floor in Boreal Mixedwood Forests. Plant Soil 2010, 336, 267–277. [Google Scholar] [CrossRef]
- Chen, H.Y.H.; Popadiouk, R.V. Dynamics of North American Boreal Mixedwoods. Environ. Rev. 2002, 10, 137–166. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier Science, B.V.: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Hume, A.; Chen, H.Y.H.; Taylor, A.R.; Kayahara, G.J.; Man, R. Soil C:N:P Dynamics During Secondary Succession Following Fire in the Boreal Forest of Central Canada. For. Ecol. Manag. 2016, 369, 1–9. [Google Scholar] [CrossRef]
- Gao, B.; Taylor, A.R.; Chen, H.Y.H.; Wang, J. Variation in Total and Volatile Carbon Concentration among the Major Tree Species of the Boreal Forest. For. Ecol. Manag. 2016, 375, 191–199. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2013.
- Garrett, L.G.; Kimberley, M.O.; Oliver, G.R.; Pearce, S.H.; Beets, P.N. Decomposition of Coarse Woody Roots and Branches in Managed Pinus Radiata Plantations in New Zealand—A Time Series Approach. For. Ecol. Manag. 2012, 269, 116–123. [Google Scholar] [CrossRef]
- Wairiu, M.; Lal, R. Soil Organic Carbon in Relation to Cultivation and Topsoil Removal on Sloping Lands of Kolombangara, Solomon Islands. Soil Tillage Res. 2003, 70, 19–27. [Google Scholar] [CrossRef]
- Willcocks, A.J.; Bell, F.W.; Williams, J.; Duinker, P.N. A Crop-Planning Process for Northern Ontario Forests: Planning Process for Northern Ontario Forests; Ministry of Natural Resources, Northwestern Ontario Forest Technology Development Unit: Thunder Bay, ON, Canada, 1997. [Google Scholar]
- Woods, M.E.; Robinson, D.C.E. Development of Fvsontario: A Forest Vegetation Simulator Variant and Application Software for Ontario. In Proceedings of the 3rd Forest Vegetation Simulator Conference, Fort Collins, CO, USA, 13–15 February 2007. [Google Scholar]
- Honer, T.G. Metric Timber Tables for the Commercial Tree Species of Central and Eastern Canada; Environment Canada: Fredericton, NB, Canada; Canadian Forestry Service: Fredericton, NB, Canada; Maritimes Forest Research Centre: Fredericton, NB, Canada, 1983.
- Ride, K.R. An Investigation of Wood Fibre Recovery and Related Economics of Four Harvesting Systems Common to Northwestern Ontario; National Library of Canada: Ottawa, ON, Canada, 2001. [Google Scholar]
- Guo, J.H.; Cameron, J.H.; Tol, R.S.J.; Anthoff, D. Discounting and the Social Cost of Carbon: A Closer Look at Uncertainty. Environ. Sci. Policy 2006, 9, 205–216. [Google Scholar] [CrossRef]
- Kumar, P.; Brondiízio, E.; Elmqvist, T.; Gatzweiler, F.; Gowdy, J.; de Groot, D.; Muradian, R.; Pascual, U.; Reyers, B.; Smith, R. Key Messages and Linkages with National and Local Policies: The Economics of Ecosystems and Biodiversity. Ecologic and Economic Foundations; Earthscan: London UK; Washington, DC, USA, 2010. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multi-Model Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Adams, D.C.; Gurevitch, J.; Rosenberg, M.S. Resampling Tests for Meta-Analysis of Ecological Data. Ecology 1997, 78, 1277–1283. [Google Scholar] [CrossRef]
- Rosseel, Y. Lavaan: An R Package for Structural Equation Modeling. J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Pyorala, P.; Peltola, H.; Strandman, H.; Antti, K.; Antti, A.; Jylha, K.; Kellomaki, S. Effects of Management on Economic Profitability of Forest Biomass Production and Carbon Neutrality of Bioenergy Use in Norway Spruce Stands under the Changing Climate. Bioenergy Res. 2014, 7, 279–294. [Google Scholar] [CrossRef]
- Taylor, A.R.; Seedre, M.; Brassard, B.W.; Chen, H.Y.H. Decline in Net Ecosystem Productivity Following Canopy Transition to Late-Succession Forests. Ecosystems 2014, 17, 778–791. [Google Scholar] [CrossRef]
- Pare, D.; Bergeron, Y. Above-Ground Biomass Accumulation Along a 230-Year Chronosequence in the Southern Portion of the Canadian Boreal Forest. J. Ecol. 1995, 83, 1001–1007. [Google Scholar] [CrossRef]
- Augusto, L.; De Schrijver, A.; Vesterdal, L.; Smolander, A.; Prescott, C.; Ranger, J. Influences of Evergreen Gymnosperm and Deciduous Angiosperm Tree Species on the Functioning of Temperate and Boreal Forests. Biol. Rev. 2015, 90, 444–466. [Google Scholar] [CrossRef] [PubMed]
- Hart, S.A.; Chen, H.Y.H. Understory Vegetation Dynamics of North American Boreal Forests. Crit. Rev. Plant Sci. 2006, 25, 381–397. [Google Scholar] [CrossRef]
- Seedre, M.; Shrestha, B.M.; Chen, H.Y.H.; Colombo, S.; Jogiste, K. Carbon Dynamics of North American Boreal Forest after Stand Replacing Wildfire and Clearcut Logging. J. For. Res. 2011, 16, 168–183. [Google Scholar] [CrossRef]
- Laganiere, J.; Pare, D.; Bergeron, Y.; Chen, H.Y.H. The Effect of Boreal Forest Composition on Soil Respiration Is Mediated through Variations in Soil Temperature and C Quality. Soil Biol. Biochem. 2012, 53, 18–27. [Google Scholar] [CrossRef]
- Laganiere, J.; Pare, D.; Bergeron, Y.; Chen, H.Y.H.; Brassard, B.W.; Cavard, X. Stability of Soil Carbon Stocks Varies with Forest Composition in the Canadian Boreal Biome. Ecosystems 2013, 16, 852–865. [Google Scholar] [CrossRef]
- Lang, S.I.; Cornelissen, J.H.C.; Klahn, T.; van Logtestijn, R.S.P.; Broekman, R.; Schweikert, W.; Aerts, R. An Experimental Comparison of Chemical Traits and Litter Decomposition Rates in a Diverse Range of Subarctic Bryophyte, Lichen and Vascular Plant Species. J. Ecol. 2009, 97, 886–900. [Google Scholar] [CrossRef]
- Brassard, B.W.; Chen, H.Y.H. Effects of Forest Type and Disturbance on Diversity of Coarse Woody Debris in Boreal Forest. Ecosystems 2008, 11, 1078–1090. [Google Scholar] [CrossRef]
- Shorohova, E.; Kapitsa, E.; Kazartsev, I.; Romashkin, I.; Polevoi, A.; Kushnevskaya, H. Tree Species Traits Are the Predominant Control on the Decomposition Rate of Tree Log Bark in a Mesic Old-Growth Boreal Forest. For. Ecol. Manag. 2016, 377, 36–45. [Google Scholar] [CrossRef]
- Silver, W.L.; Miya, R.K. Global Patterns in Root Decomposition: Comparisons of Climate and Litter Quality Effects. Oecologia 2001, 129, 407–419. [Google Scholar] [CrossRef]
- Ma, Z.; Chen, H.Y.H. Positive Species Mixture Effects on Fine Root Turnover and Mortality in Natural Boreal Forests. Soil Biol. Biochem. 2018, 121, 130–137. [Google Scholar] [CrossRef]
- Bennett, E.M.; Peterson, G.D.; Gordon, L.J. Understanding Relationships among Multiple Ecosystem Services. Ecol. Lett. 2009, 12, 1394–1404. [Google Scholar] [CrossRef] [PubMed]
- van der Plas, F.; Ratcliffe, S.; Ruiz-Benito, P.; Scherer-Lorenzen, M.; Verheyen, K.; Wirth, C.; Zavala, M.A.; Ampoorter, E.; Baeten, L.; Barbaro, L.; et al. Continental Mapping of Forest Ecosystem Functions Reveals a High but Unrealised Potential for Forest Multifunctionality. Ecol. Lett. 2018, 21, 31–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, S.; Bell, F.W.; Chen, H.Y.H. Long-Term Effects of Intensive Silvicultural Practices on Productivity, Composition, and Structure of Northern Temperate and Boreal Plantations in Ontario, Canada. For. Ecol. Manag. 2007, 241, 115–126. [Google Scholar] [CrossRef]
- Bell, F.W.; Parton, J.; Stocker, N.; Joyce, D.; Reid, D.; Wester, M.; Stinson, A.; Kayahara, G.; Towill, B. Developing a Silvicultural Framework and Definitions for Use in Forest Management Planning and Practice. For. Chron. 2008, 84, 678–693. [Google Scholar] [CrossRef]
- Chen, S.; Shahi, C.; Chen, H.Y.H.; Kumar, P.; Ma, Z.; McLaren, B. Trade-Offs and Synergies between Economic Gains and Plant Diversity across a Range of Management Alternatives in Boreal Forests. Ecol. Econ. 2018, 151, 162–172. [Google Scholar] [CrossRef]
- Chen, S.; Shahi, C.; Chen, H.Y.H. Economic and Ecological Trade-Off Analysis of Forest Ecosystems: Options for Boreal Forests. Environ. Rev. 2016, 24, 348–361. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Z.; Chen, S.; Shahi, C.; Chen, H.Y.H.; Chen, H. Trade-Offs between Economic Gains and Carbon Stocks across a Range of Management Alternatives in Boreal Forests. Forests 2022, 13, 1777. https://doi.org/10.3390/f13111777
Ma Z, Chen S, Shahi C, Chen HYH, Chen H. Trade-Offs between Economic Gains and Carbon Stocks across a Range of Management Alternatives in Boreal Forests. Forests. 2022; 13(11):1777. https://doi.org/10.3390/f13111777
Chicago/Turabian StyleMa, Zilong, Si Chen, Chander Shahi, Han Y. H. Chen, and Hao Chen. 2022. "Trade-Offs between Economic Gains and Carbon Stocks across a Range of Management Alternatives in Boreal Forests" Forests 13, no. 11: 1777. https://doi.org/10.3390/f13111777
APA StyleMa, Z., Chen, S., Shahi, C., Chen, H. Y. H., & Chen, H. (2022). Trade-Offs between Economic Gains and Carbon Stocks across a Range of Management Alternatives in Boreal Forests. Forests, 13(11), 1777. https://doi.org/10.3390/f13111777