
Genome-Wide Identification and Expression Analysis of the MADS-Box Family in Ginkgo biloba

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Identification of MADS-box Family Genes in G. biloba
2.3. Chromosomal Location and Collinearity Analyses for GbMADS Genes
2.4. Conserved Motifs, and Gene Structure Analysis
2.5. Phylogenetic Analysis and Classification of the GbMADS Genes
2.6. miRNA Target Prediction
2.7. Promoter Analysis of the GbMADS Genes
2.8. Gene Expression Analysis
3. Results
3.1. Identification and Physicochemical Properties of the MADS-Box Gene Family in G. biloba
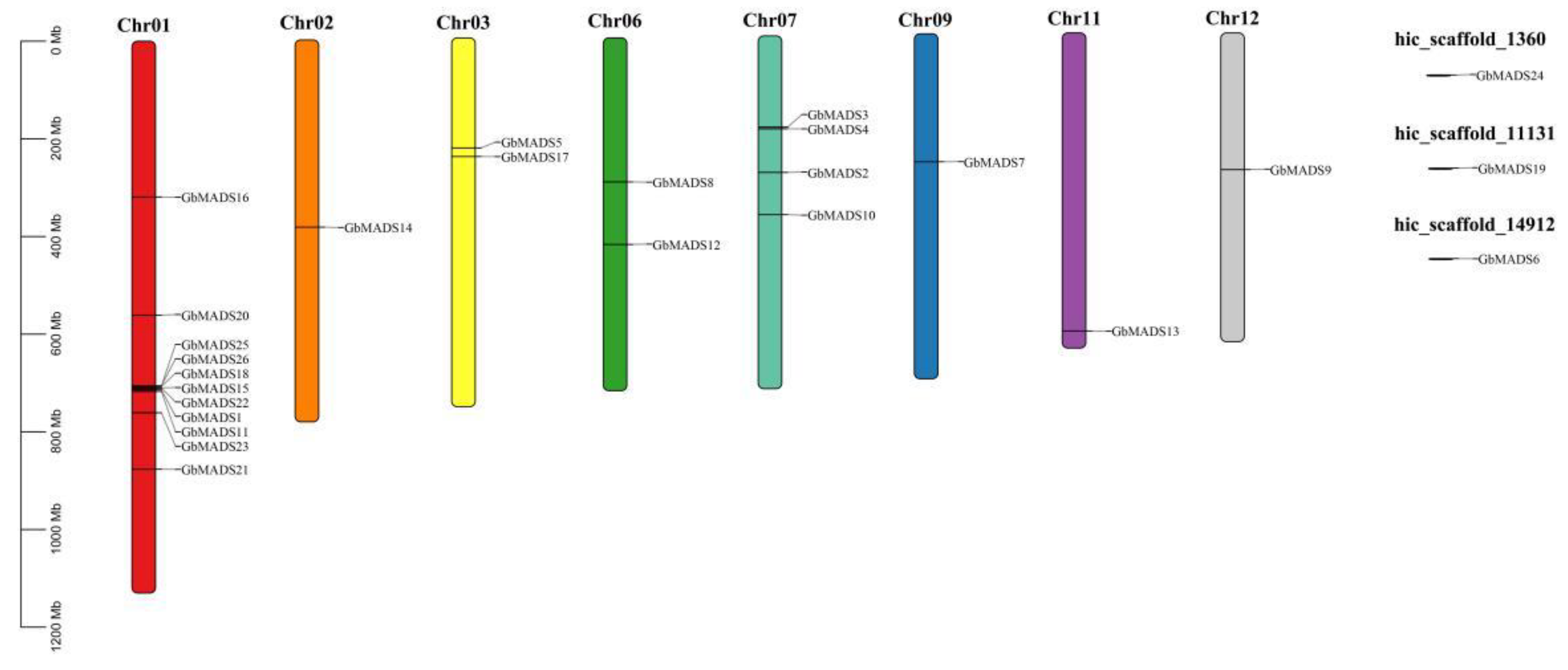
3.2. Chromosome Mapping and Collinearity Analysis of GbMADS
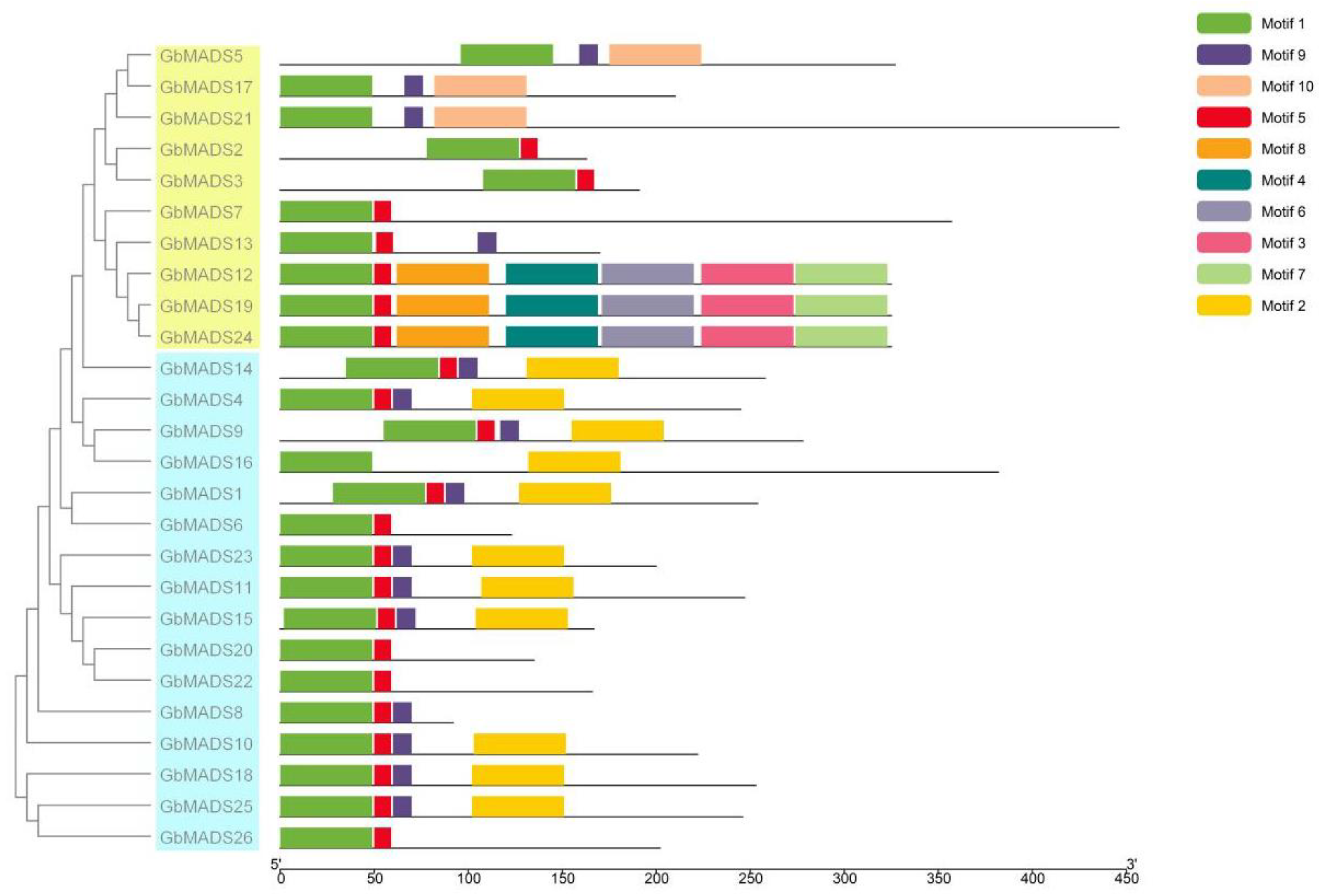
3.3. Classification, Conserved Motifs, and Gene Structure Analysis of the GbMADS Family
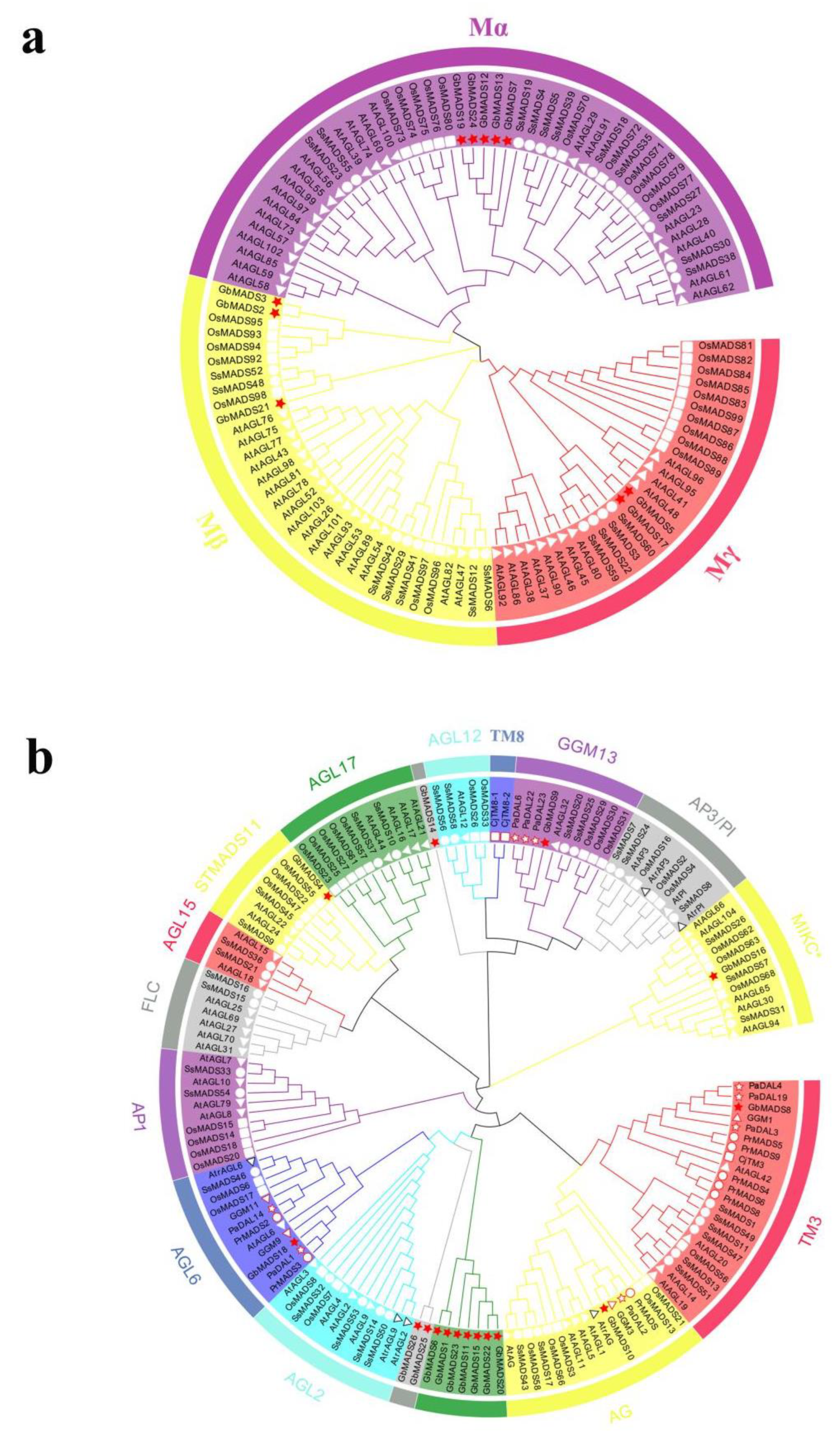
3.4. Phylogenetic Analysis of the GbMADS Gene Family
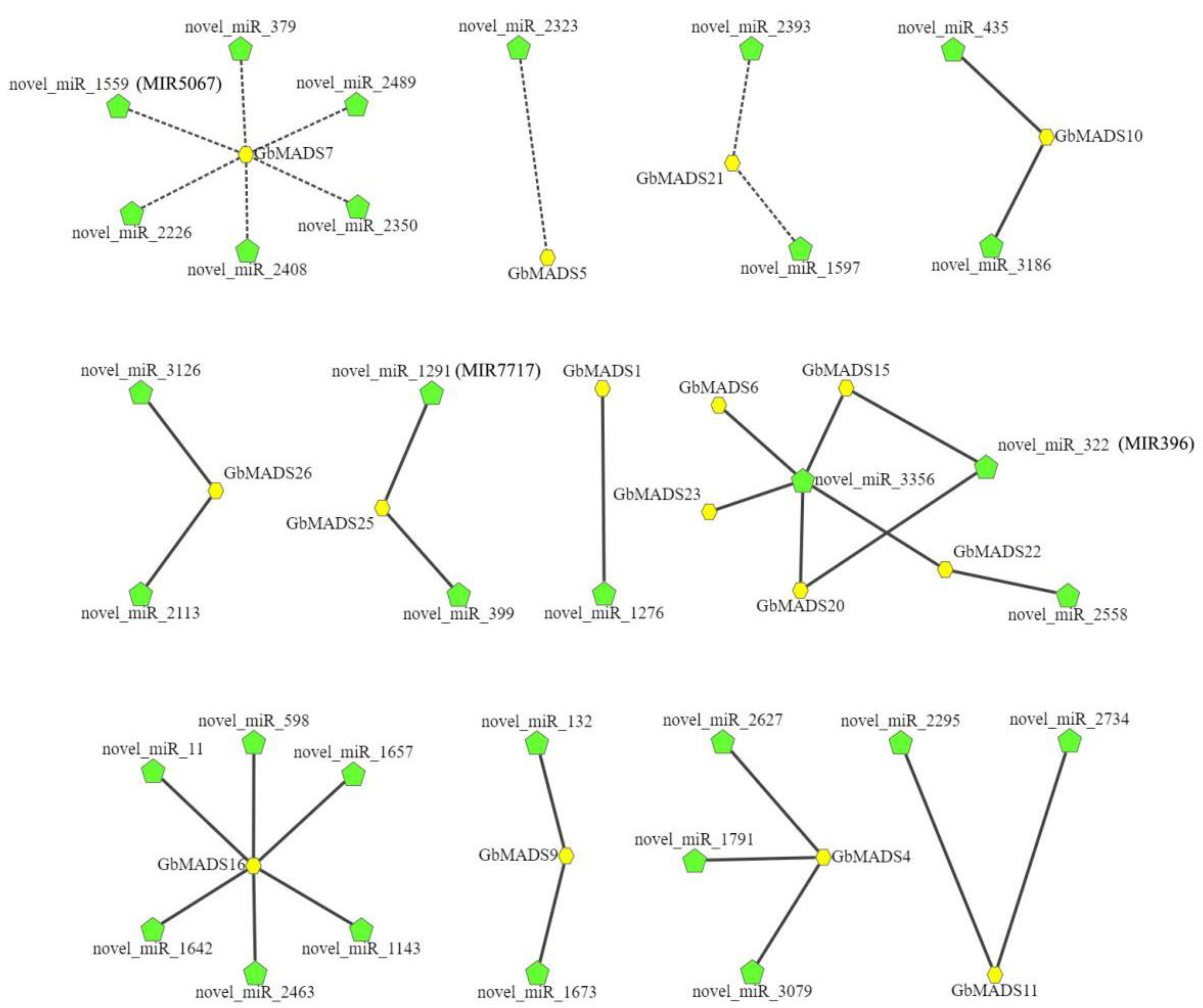
3.5. Prediction of the miRNA Target Sites of the GbMADS Gene Family
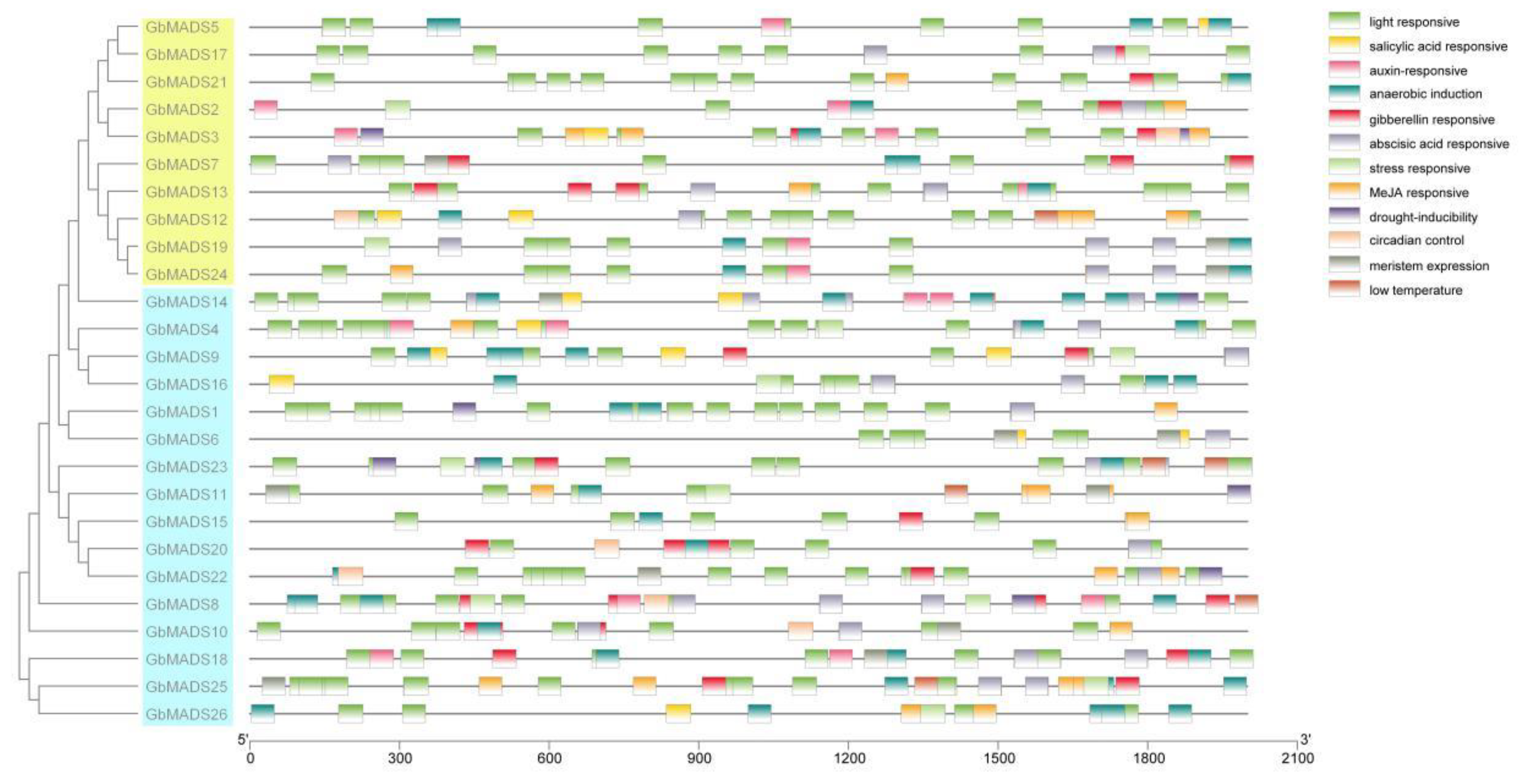
3.6. Promoter Sequence Analysis of the GbMADS Gene Family
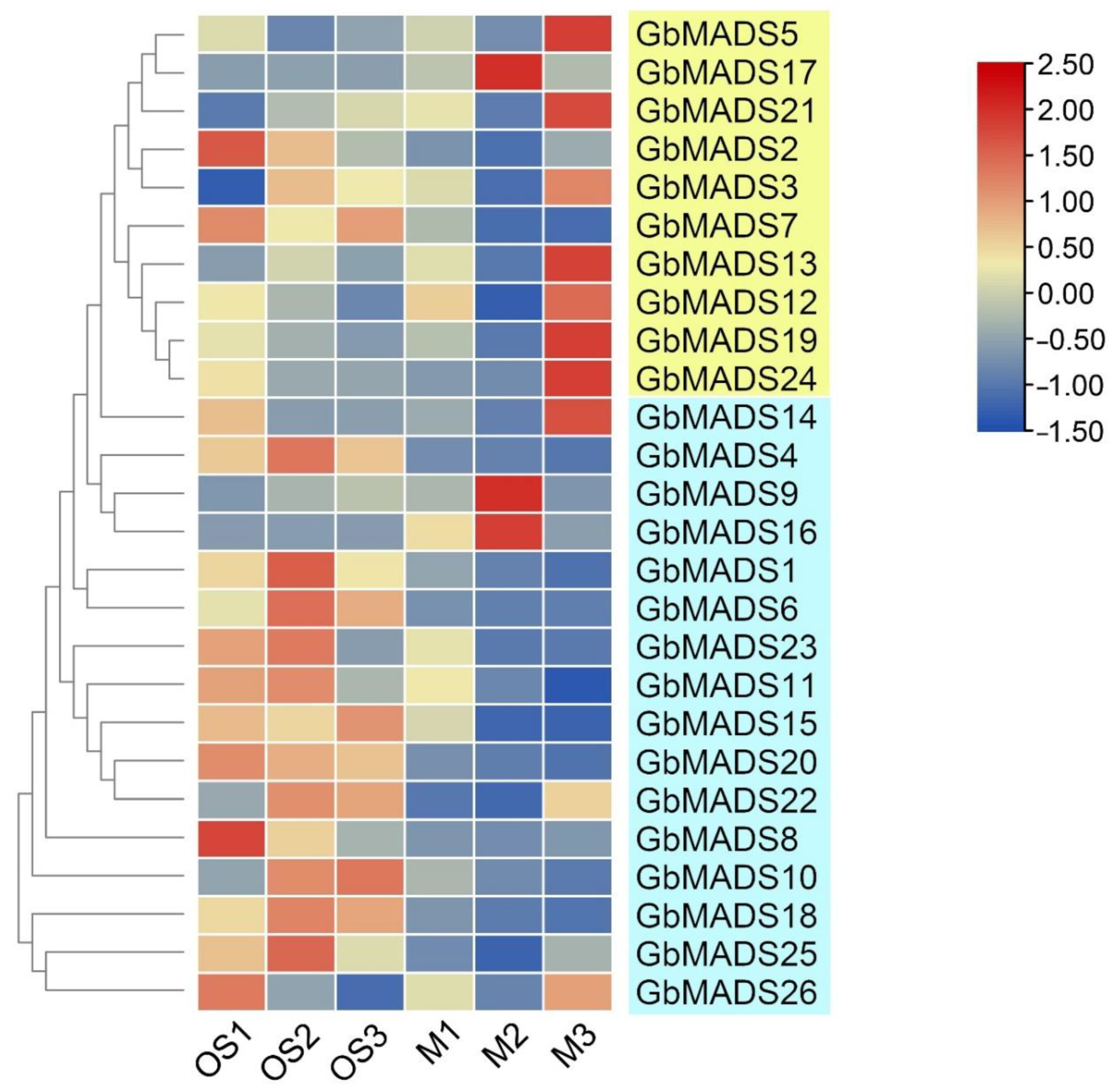
3.7. Expression Profile Analysis of the GbMADS Genes in the Male and Female Flowers of G. biloba
4. Discussion
4.1. Genealogical Evolution of the GbMADS Genes in G. biloba
4.2. Bioinformatics Prediction of GbMADS Gene’s Function
4.3. Expression Pattern and Function of the GbMADS Genes in the Male and Female Flowers of G. biloba
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liu, H.; Cao, F.; Yin, T.; Chen, Y. A highly dense genetic map for Ginkgo biloba constructed using sequence-based markers. Front. Plant Sci. 2017, 8, 1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.B.; Cheng, S.Y.; Zhou, X.; Chen, Z.X.; Kim, S.U.; Tan, J.P.; Zheng, J.R.; Xu, F.; Zhang, W.W.; Liao, Y.L.; et al. A global survey of full-length transcriptome of Ginkgo biloba reveals transcript variants involved in flavonoid biosynthesis. Ind. Crops Prod. 2019, 139, 111547. [Google Scholar] [CrossRef]
- Ye, J.B.; Zhang, X.; Tan, J.P.; Xu, F.; Cheng, S.Y.; Chen, Z.X.; Zhang, W.W.; Liao, Y.L. Global identification of Ginkgo biloba microRNAs and insight into their role in metabolism regulatory network of terpene trilactones by high-throughput sequencing and degradome analysis. Ind. Crops Prod. 2020, 148, 112289. [Google Scholar] [CrossRef]
- Yang, Y.; Li, Y.; Wang, J.; Sun, K.; Tao, W.; Wang, Z.; Xiao, W.; Pan, Y.; Zhang, S.; Wang, Y. Systematic investigation of Ginkgo biloba leaves for treating cardio-cerebrovascular diseases in an animal model. ACS Chem. Biol. 2017, 12, 1363–1372. [Google Scholar] [CrossRef]
- Li, M.; Li, B.; Xia, Z.M.; Tian, Y.; Zhang, D.; Rui, W.J.; Dong, J.X.; Xiao, F.J. Anticancer effects of five biflavonoids from Ginkgo biloba L. male flowers in vitro. Molecules 2019, 24, 1496. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.; Wang, Z.; Wang, P.; Wang, M. Extraction, structure and bioactivities of the polysaccharides from Ginkgo biloba: A review. Int. J. Biol. Macromol. 2020, 162, 1897–1905. [Google Scholar] [CrossRef]
- Mei, N.; Guo, X.; Ren, Z.; Kobayashi, D.; Wada, K.; Guo, L. Review of Ginkgo biloba-induced toxicity, from experimental studies to human case reports. J. Environ. Sci. Health. Part C Environ. Carcinog. Ecotoxicol. Rev. 2017, 35, 1–28. [Google Scholar] [CrossRef]
- Chassagne, F.; Huang, X.; Lyles, J.T.; Quave, C.L. Validation of a 16th century traditional chinese medicine use of Ginkgo biloba as a topical antimicrobial. Front. Microbiol. 2019, 10, 775. [Google Scholar] [CrossRef]
- Becker, A.; Theissen, G. The major clades of MADS-box genes and their role in the development and evolution of flowering plants. Mol. Phylogenet. Evol. 2003, 29, 464–489. [Google Scholar] [CrossRef]
- Henschel, K.; Kofuji, R.; Hasebe, M.; Saedler, H.; Münster, T.; Theissen, G. Two ancient classes of MIKC-type MADS-box genes are present in the moss Physcomitrella patens. Mol. Biol. Evol. 2002, 19, 801–814. [Google Scholar] [CrossRef]
- Masiero, S.; Colombo, L.; Grini, P.E.; Schnittger, A.; Kater, M.M. The emerging importance of type I MADS box transcription factors for plant reproduction. Plant Cell 2011, 23, 865–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gramzow, L.; Theissen, G. A hitchhiker’s guide to the MADS world of plants. Genome Biol. 2010, 11, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jack, T.; Brockman, L.L.; Meyerowitz, E.M. The homeotic gene APETALA3 of Arabidopsis thaliana encodes a MADS box and is expressed in petals and stamens. Cell 1992, 68, 683–697. [Google Scholar] [CrossRef]
- Rümpler, F.; Gramzow, L.; Theißen, G.; Melzer, R. Did convergent protein evolution enable phytoplasmas to generate ‘Zombie Plants’? Trends Plant Sci. 2015, 20, 798–806. [Google Scholar] [CrossRef]
- Kaufmann, K.; Melzer, R.; Theissen, G. MIKC-type MADS-domain proteins: Structural modularity, protein interactions and network evolution in land plants. Gene 2005, 347, 183–198. [Google Scholar] [CrossRef]
- Bowman, J.L.; Smyth, D.R.; Meyerowitz, E.M. Genetic interactions among floral homeotic genes of Arabidopsis. Development 1991, 112, 1–20. [Google Scholar] [CrossRef]
- Colombo, L.; Franken, J.; Koetje, E.; van Went, J.; Dons, H.J.; Angenent, G.C.; van Tunen, A.J. The petunia MADS box gene FBP11 determines ovule identity. Plant Cell 1995, 7, 1859–1868. [Google Scholar] [CrossRef] [Green Version]
- Causier, B.; Schwarz-Sommer, Z.; Davies, B. Floral organ identity: 20 years of ABCs. Semin. Cell Dev. Biol. 2010, 21, 73–79. [Google Scholar] [CrossRef]
- Kwantes, M.; Liebsch, D.; Verelst, W. How MIKC * MADS-box genes originated and evidence for their conserved function throughout the evolution of vascular plant gametophytes. Mol. Biol. Evol. 2012, 29, 293–302. [Google Scholar] [CrossRef]
- Melzer, R.; Wang, Y.Q.; Theissen, G. The naked and the dead: The ABCs of gymnosperm reproduction and the origin of the angiosperm flower. Semin. Cell Dev. Biol. 2010, 21, 118–128. [Google Scholar] [CrossRef]
- Nystedt, B.; Street, N.R.; Wetterbom, A.; Zuccolo, A.; Lin, Y.C.; Scofield, D.G.; Vezzi, F.; Delhomme, N.; Giacomello, S.; Alexeyenko, A.; et al. The Norway spruce genome sequence and conifer genome evolution. Nature 2013, 497, 579–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gramzow, L.; Weilandt, L.; Theißen, G. MADS goes genomic in conifers: Towards determining the ancestral set of MADS-box genes in seed plants. Ann. Bot. 2014, 114, 1407–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, S.; Yuan, H.; Sun, X.; Porth, I.; Li, Y.; El-Kassaby, Y.A.; Li, W. A transcriptomics investigation into pine reproductive organ development. New Phytol. 2016, 209, 1278–1289. [Google Scholar] [CrossRef] [PubMed]
- Deng, N.; Hou, C.; He, B.; Ma, F.; Song, Q.; Shi, S.; Liu, C.; Tian, Y. A full-length transcriptome and gene expression analysis reveal genes and molecular elements expressed during seed development in Gnetum luofuense. BMC Plant Biol. 2020, 20, 531. [Google Scholar] [CrossRef]
- Yang, F.; Xu, F.; Wang, X.H.; Liao, Y.L.; Chen, Q.W.; Meng, X.X. Characterization and functional analysis of a MADS-box transcription factor gene (GbMADS9) from Ginkgo biloba. Sci. Hortic. 2016, 212, 104–114. [Google Scholar] [CrossRef]
- Parenicová, L.; de Folter, S.; Kieffer, M.; Horner, D.S.; Favalli, C.; Busscher, J.; Cook, H.E.; Ingram, R.M.; Kater, M.M.; Davies, B.; et al. Molecular and phylogenetic analyses of the complete MADS-box transcription factor family in Arabidopsis: New openings to the MADS world. Plant Cell 2003, 15, 1538–1551. [Google Scholar] [CrossRef] [Green Version]
- Masiero, S.; Li, M.A.; Will, I.; Hartmann, U.; Saedler, H.; Huijser, P.; Schwarz-Sommer, Z.; Sommer, H. INCOMPOSITA: A MADS-box gene controlling prophyll development and floral meristem identity in Antirrhinum. Development 2004, 131, 5981–5990. [Google Scholar] [CrossRef] [Green Version]
- Leseberg, C.H.; Li, A.; Kang, H.; Duvall, M.; Mao, L. Genome-wide analysis of the MADS-box gene family in Populus trichocarpa. Gene 2006, 378, 84–94. [Google Scholar] [CrossRef]
- Duan, W.; Song, X.; Liu, T.; Huang, Z.; Ren, J.; Hou, X.; Li, Y. Genome-wide analysis of the MADS-box gene family in Brassica rapa (Chinese cabbage). Mol. Genet. Genom. 2015, 290, 239–255. [Google Scholar] [CrossRef]
- Hu, J.; Chang, X.; Zhang, Y.; Yu, X.; Qin, Y.; Sun, Y.; Zhang, L. The pineapple MADS-box gene family and the evolution of early monocot flower. Sci. Rep. 2021, 11, 849. [Google Scholar] [CrossRef]
- Tian, Y.; Dong, Q.; Ji, Z.; Chi, F.; Cong, P.; Zhou, Z. Genome-wide identification and analysis of the MADS-box gene family in apple. Gene 2015, 555, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Paul, P.; Dhatt, B.K.; Miller, M.; Folsom, J.J.; Wang, Z.; Krassovskaya, I.; Liu, K.; Sandhu, J.; Yu, H.; Zhang, C.; et al. MADS78 and MADS79 are essential regulators of early seed development in rice. Plant Physiol. 2020, 182, 933–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatima, M.; Zhang, X.; Lin, J.; Zhou, P.; Zhou, D.; Ming, R. Expression profiling of MADS-box gene family revealed its role in vegetative development and stem ripening in S. spontaneum. Sci. Rep. 2020, 10, 20536. [Google Scholar] [CrossRef]
- Ortuño-Miquel, S.; Rodríguez-Cazorla, E.; Zavala-Gonzalez, E.A.; Martínez-Laborda, A.; Vera, A. Arabidopsis HUA ENHANCER 4 delays flowering by upregulating the MADS-box repressor genes FLC and MAF4. Sci. Rep. 2019, 9, 1478. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Yang, L.; Wu, D.; Meng, Q.; Deng, X.; Huang, G.; Zhang, J.; Chen, X.; Ferrándiz, C.; Liang, W.; et al. Rice SEPALLATA genes OsMADS5 and OsMADS34 cooperate to limit inflorescence branching by repressing the TERMINAL FLOWER1-like gene RCN4. New Phytol. 2021, 233, 1682–1700. [Google Scholar] [CrossRef] [PubMed]
- Schilling, S.; Pan, S.; Kennedy, A.; Melzer, R. MADS-box genes and crop domestication: The jack of all traits. J. Exp. Bot. 2018, 69, 1447–1469. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, Y.; Naeem, M.; Zhu, M.; Li, J.; Yu, X.; Hu, Z.; Chen, G. An AGAMOUS MADS-box protein, SlMBP3, regulates the speed of placenta liquefaction and controls seed formation in tomato. J. Exp. Bot. 2019, 70, 909–924. [Google Scholar] [CrossRef]
- Jibran, R.; Tahir, J.; Cooney, J.; Hunter, D.A.; Dijkwel, P.P. Arabidopsis AGAMOUS regulates sepal senescence by driving jasmonate production. Front. Plant Sci. 2017, 8, 2101. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Qu, Y.; Bi, C.; He, B.; Ye, N.; Yin, T.; Xu, L. Genome-wide identification and characterization of the MADS-box gene family in Salix suchowensis. PeerJ 2019, 7, e8019. [Google Scholar] [CrossRef]
- Arora, R.; Agarwal, P.; Ray, S.; Singh, A.K.; Singh, V.P.; Tyagi, A.K.; Kapoor, S. MADS-box gene family in rice: Genome-wide identification, organization and expression profiling during reproductive development and stress. BMC Genom. 2007, 8, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlsbecker, A.; Sundström, J.F.; Englund, M.; Uddenberg, D.; Izquierdo, L.; Kvarnheden, A.; Vergara-Silva, F.; Engström, P. Molecular control of normal and acrocona mutant seed cone development in Norway spruce (Picea abies) and the evolution of conifer ovule-bearing organs. New Phytol. 2013, 200, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Winter, K.U.; Becker, A.; Münster, T.; Theissen, G. MADS-box genes reveal that gnetophytes are more closely related to conifers than to flowering plants. Proc. Natl. Acad. Sci. USA 1999, 96, 7342–7347. [Google Scholar] [CrossRef] [Green Version]
- Futamura, N.; Totoki, Y.; Toyoda, A.; Igasaki, T.; Nanjo, T.; Seki, M.; Sakaki, Y.; Mari, A.; Shinozaki, K.; Shinohara, K. Characterization of expressed sequence tags from a full-length enriched cDNA library of Cryptomeria japonica male strobili. BMC Genom. 2008, 9, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.M.; Cheng, S.Y.; Ye, J.B.; Chen, Z.X.; Liao, Y.L.; Zhang, W.W.; Kim, S.U.; Xu, F. Screening and identification of miRNAs related to sexual differentiation of strobili in Ginkgo biloba by integration analysis of small RNA, RNA, and degradome sequencing. BMC Plant Biol. 2020, 20, 387. [Google Scholar] [CrossRef]
- Su, X.Y.; Shi, Y.B.; Yang, X.M.; Wang, G.B.; Cao, F.L. Selection and verification of internal reference genes in real-time fluorescence quantitative PCR analysis of Ginkgo biloba. Plant Physiol. J. 2019, 55, 875–882. (in Chinese). [Google Scholar] [CrossRef]
- Yang, K.; Han, H.; Li, Y.; Ye, J.; Xu, F. Significance of miRNA in enhancement of flavonoid biosynthesis. Plant Biol. 2021, 24, 217–226. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Hu, Z.; Guo, X.; Tian, S.; Chen, G. Genome-wide analysis of the MADS-box transcription factor family in Solanum lycopersicum. Int. J. Mol. Sci. 2019, 20, 2961. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Hao, Z.; Long, X.; Wang, Z.; Zheng, X.; Ye, D.; Peng, Y.; Wu, W.; Hu, X.; Wang, G.; et al. The Transcriptome of Cunninghamia lanceolata male/female cone reveal the association between MIKC MADS-box genes and reproductive organs development. BMC Plant Biol. 2020, 20, 508. [Google Scholar] [CrossRef]
- Jin, J.; Tian, F.; Yang, D.C.; Meng, Y.Q.; Kong, L.; Luo, J.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res. 2017, 45, D1040–D1045. [Google Scholar] [CrossRef]
- Tian, F.; Yang, D.C.; Meng, Y.Q.; Jin, J.; Gao, G. PlantRegMap: Charting functional regulatory maps in plants. Nucleic Acids Res. 2020, 48, D1104–D1113. [Google Scholar] [CrossRef] [PubMed]
- Guan, R.; Zhao, Y.; Zhang, H.; Fan, G.; Liu, X.; Zhou, W.; Shi, C.; Wang, J.; Liu, W.; Liang, X.; et al. Draft genome of the living fossil Ginkgo biloba. Gigascience 2016, 5, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrándiz, C.; Gu, Q.; Martienssen, R.; Yanofsky, M.F. Redundant regulation of meristem identity and plant architecture by FRUITFULL, APETALA1 and CAULIFLOWER. Development 2000, 127, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Pelaz, S.; Ditta, G.S.; Baumann, E.; Wisman, E.; Yanofsky, M.F. B and C floral organ identity functions require SEPALLATA MADS-box genes. Nature 2000, 405, 200–203. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, C.C.; Rouse, D.T.; Finnegan, E.J.; Peacock, W.J.; Dennis, E.S. The molecular basis of vernalization: The central role of FLOWERING LOCUS C (FLC). Proc. Natl. Acad. Sci. USA 2000, 97, 3753–3758. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, X.; Wang, G.; Cui, P.; Wu, S.; Ai, C.; Hu, N.; Li, A.; He, B.; Shao, X.; et al. The nearly complete genome of Ginkgo biloba illuminates gymnosperm evolution. Nat. Plants 2021, 7, 748–756. [Google Scholar] [CrossRef]
- Chen, F.; Zhang, X.; Liu, X.; Zhang, L. Evolutionary analysis of MIKCc-type MADS-box genes in gymnosperms and angiosperms. Front. Plant Sci. 2017, 8, 895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapia-López, R.; García-Ponce, B.; Dubrovsky, J.G.; Garay-Arroyo, A.; Pérez-Ruíz, R.V.; Kim, S.H.; Acevedo, F.; Pelaz, S.; Alvarez-Buylla, E.R. An AGAMOUS-related MADS-box gene, XAL1 (AGL12), regulates root meristem cell proliferation and flowering transition in Arabidopsis. Plant Physiol. 2008, 146, 1182–1192. [Google Scholar] [CrossRef] [Green Version]
- Niu, Q.; Li, J.; Cai, D.; Qian, M.; Jia, H.; Bai, S.; Hussain, S.; Liu, G.; Teng, Y.; Zheng, X. Dormancy-associated MADS-box genes and microRNAs jointly control dormancy transition in pear (Pyrus pyrifolia white pear group) flower bud. J. Exp. Bot. 2016, 67, 239–257. [Google Scholar] [CrossRef] [Green Version]
- Aceto, S.; Sica, M.; De Paolo, S.; D’Argenio, V.; Cantiello, P.; Salvatore, F.; Gaudio, L. The analysis of the inflorescence miRNome of the orchid Orchis italica reveals a DEF-like MADS-box gene as a new miRNA target. PLoS ONE 2014, 9, e97839. [Google Scholar] [CrossRef]
- Yang, C.Y.; Huang, Y.H.; Lin, C.P.; Lin, Y.Y.; Hsu, H.C.; Wang, C.N.; Daisy Liu, L.Y.; Shen, B.N.; Lin, S.S. MicroRNA396-targeted SHORT VEGETATIVE PHASE is required to repress flowering and is related to the development of abnormal flower symptoms by the phyllody symptoms1 effector. Plant Physiol. 2015, 168, 1702–1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, S.; Li, Z.; Yuan, N.; Hu, Q.; Zhou, M.; Zhao, J.; Li, D.; Luo, H. MiR396 is involved in plant response to vernalization and flower development in Agrostis stolonifera. Hortic. Res. 2020, 7, 173. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Chromosome Location | CDS (bp) | Protein Length (aa) | MW (KDa) | GRAVY | pI | II | AI | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|---|---|
| GbMADS1 | Gb_01884 | Chr1: 713,430,121 … 713889578 | 762 | 253 | 28.57 | −0.521 | 9.8 | 45.66 | 87.11 | Nucleus |
| GbMADS2 | Gb_03068 | Chr7: 279,659,904 … 279661933 | 489 | 162 | 18.42 | −0.608 | 9.96 | 58.83 | 78.27 | Nucleus |
| GbMADS3 | Gb_03807 | Chr7: 186,875,289 … 186876549 | 573 | 190 | 20.98 | −0.104 | 10.05 | 32.83 | 88.26 | Extracell |
| GbMADS4 | Gb_05128 | Chr7: 190,259,000 … 190423917 | 735 | 244 | 28.05 | −0.668 | 4.96 | 44.36 | 86.27 | Nucleus |
| GbMADS5 | Gb_05359 | Chr3: 225,287,517 … 225289874 | 1320 | 327 | 37.65 | −0.738 | 5.31 | 64.01 | 71.87 | Nucleus |
| GbMADS6 | Gb_12581 | Scaffold 14912: 35,255 … 35785 | 369 | 122 | 13.74 | −0.397 | 10.02 | 40.68 | 83.11 | Nucleus |
| GbMADS7 | Gb_12586 | Chr9: 262,143,415 … 262144485 | 1071 | 356 | 40.51 | −0.65 | 6.32 | 60.9 | 71.8 | Nucleus |
| GbMADS8 | Gb_12778 | Chr6: 294,934,960 … 295095100 | 276 | 91 | 10.58 | −0.693 | 9.76 | 35.09 | 70.77 | Nucleus |
| GbMADS9 | Gb_15398 | Chr12: 279,825,002 … 279826961 | 834 | 277 | 31.89 | −0.405 | 8.86 | 40.93 | 81.66 | Nucleus |
| GbMADS10 | Gb_16301 | Chr7: 366,097,174 … 366139501 | 666 | 221 | 25.38 | −0.605 | 8.96 | 60.62 | 85.25 | Nucleus |
| GbMADS11 | Gb_19178 | Chr1: 717,036,617 … 717203413 | 741 | 246 | 28.20 | −0.594 | 6.26 | 59.86 | 84.31 | Chloroplast |
| GbMADS12 | Gb_19258 | Chr6: 423,032,522 … 423033532 | 1011 | 336 | 37.20 | −0.704 | 8.33 | 62.85 | 67.71 | Nucleus |
| GbMADS13 | Gb_21526 | Chr11: 610,402,766 … 610403275 | 510 | 169 | 19.41 | −0.499 | 10.17 | 53.10 | 79.59 | Chloroplast |
| GbMADS14 | Gb_28587 | Chr2: 383,363,358 … 383365563 | 774 | 257 | 29.39 | −0.416 | 9.90 | 51.03 | 98.68 | Nucleus |
| GbMADS15 | Gb_30604 | Chr1: 709,778,670 … 709874204 | 501 | 166 | 19.37 | −0.710 | 10.20 | 55.36 | 86.39 | Nucleus |
| GbMADS16 | Gb_31417 | Chr1: 318,984,240 … 319003647 | 1146 | 381 | 43.30 | −0.672 | 5.96 | 47.31 | 73.02 | Nucleus |
| GbMADS17 | Gb_33168 | Chr3: 242,930,058 … 242930687 | 630 | 209 | 24.35 | −0.708 | 5.53 | 63.85 | 80.19 | Nucleus |
| GbMADS18 | Gb_36364 | Chr1: 707,044,181 … 707202195 | 759 | 252 | 29.17 | −0.917 | 8.88 | 54.23 | 77.30 | Nucleus |
| GbMADS19 | Gb_37613 | Scaffold 11131: 15,283 … 16293 | 1011 | 336 | 37.28 | −0.680 | 8.33 | 58.81 | 67.11 | Nucleus |
| GbMADS20 | Gb_38365 | Chr1: 560,863,035 … 560864326 | 405 | 134 | 15.27 | −0.480 | 9.34 | 50.94 | 72.09 | Nucleus |
| GbMADS21 | Gb_38883 | Chr1: 876,632,006 … 876634445 | 1338 | 445 | 50.95 | −0.667 | 5.93 | 47.36 | 69.69 | Nucleus |
| GbMADS22 | Gb_38922 | Chr1: 710,260,574 … 710261629 | 498 | 165 | 18.37 | 0.131 | 9.37 | 38.36 | 96.42 | Nucleus |
| GbMADS23 | Gb_39109 | Chr1: 760,814,262 … 760970131 | 600 | 199 | 22.58 | −0.513 | 6.22 | 66.05 | 90.15 | Nucleus |
| GbMADS24 | Gb_40092 | Scaffold 1360: 32,384 … 33394 | 1011 | 336 | 37.26 | −0.687 | 8.33 | 59.06 | 66.55 | Nucleus |
| GbMADS25 | Gb_41549 | Chr1: 705,678,899 … 705806511 | 738 | 245 | 28.35 | −0.754 | 9.61 | 46.67 | 82.37 | Nucleus |
| GbMADS26 | Gb_41550 | Chr1: 705,910,701 … 705911828 | 606 | 201 | 22.54 | −0.051 | 9.28 | 59.08 | 100.40 | Nucleus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, K.; Liu, Z.; Chen, X.; Zhou, X.; Ye, J.; Xu, F.; Zhang, W.; Liao, Y.; Yang, X.; Wang, Q. Genome-Wide Identification and Expression Analysis of the MADS-Box Family in Ginkgo biloba. Forests 2022, 13, 1953. https://doi.org/10.3390/f13111953
Yang K, Liu Z, Chen X, Zhou X, Ye J, Xu F, Zhang W, Liao Y, Yang X, Wang Q. Genome-Wide Identification and Expression Analysis of the MADS-Box Family in Ginkgo biloba. Forests. 2022; 13(11):1953. https://doi.org/10.3390/f13111953
Chicago/Turabian StyleYang, Ke, Zhongbing Liu, Xueyin Chen, Xian Zhou, Jiabao Ye, Feng Xu, Weiwei Zhang, Yongling Liao, Xiaoyan Yang, and Qijian Wang. 2022. "Genome-Wide Identification and Expression Analysis of the MADS-Box Family in Ginkgo biloba" Forests 13, no. 11: 1953. https://doi.org/10.3390/f13111953
APA StyleYang, K., Liu, Z., Chen, X., Zhou, X., Ye, J., Xu, F., Zhang, W., Liao, Y., Yang, X., & Wang, Q. (2022). Genome-Wide Identification and Expression Analysis of the MADS-Box Family in Ginkgo biloba. Forests, 13(11), 1953. https://doi.org/10.3390/f13111953