
The Relationship between Landscape Patterns and Populations of Asian Longhorned Beetles



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Regions
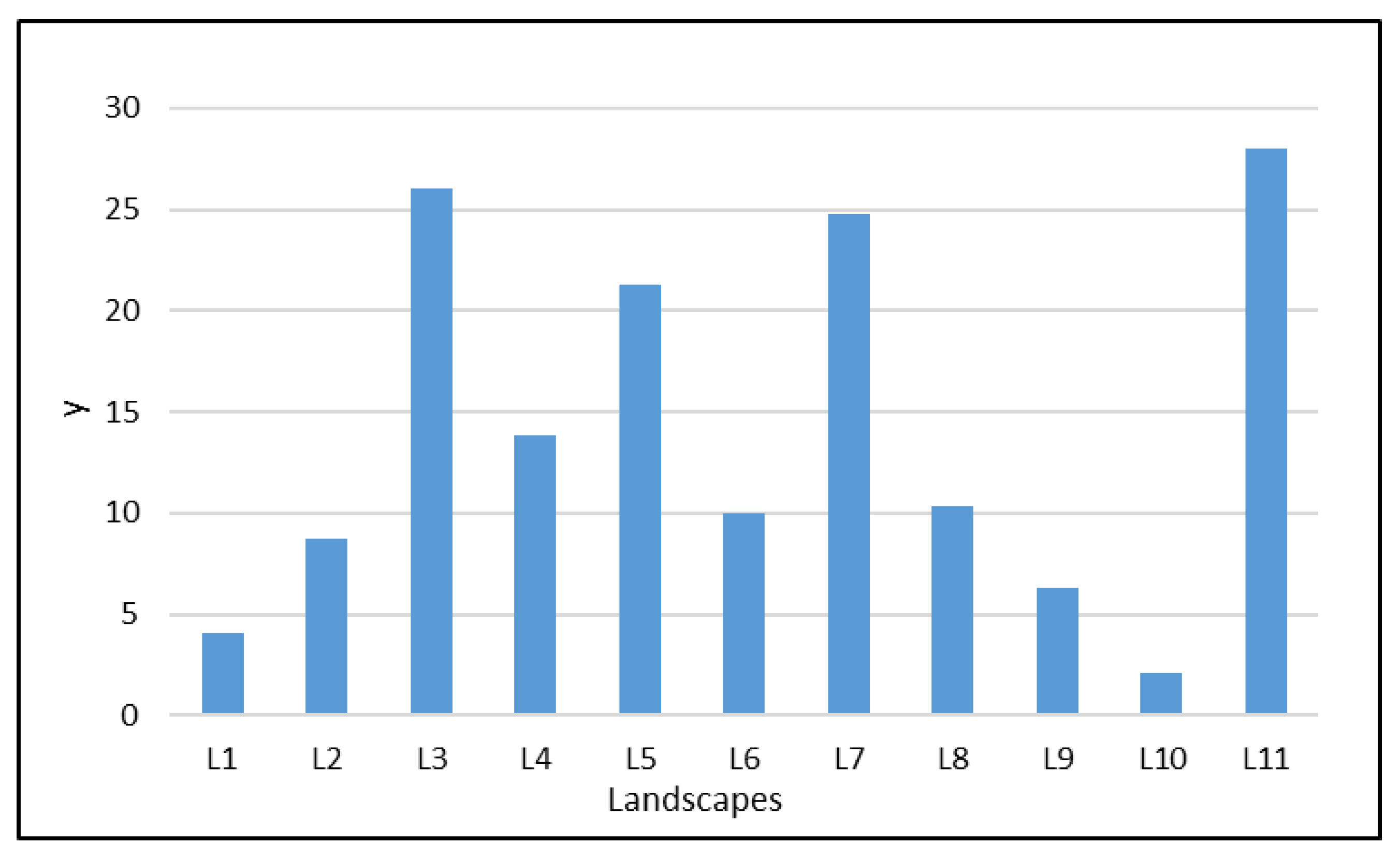
2.2. Surveys
2.3. Landscape Metrics and Mapping
2.4. Data Analysis
3. Results
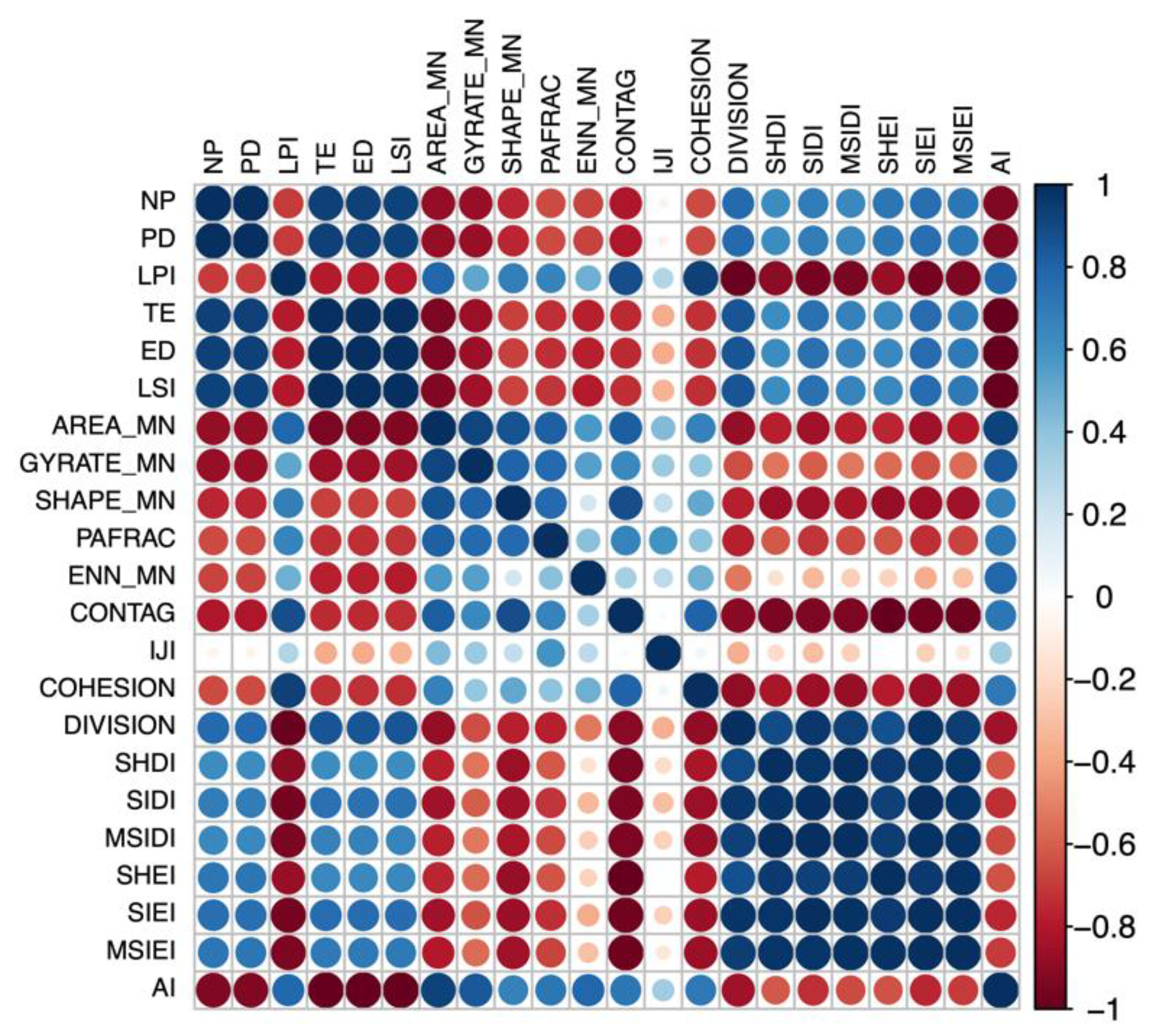
3.1. Correlation of Landscape-Level Metrics and ALBs
3.2. Correlation of Class-Level Metrics and ALBs
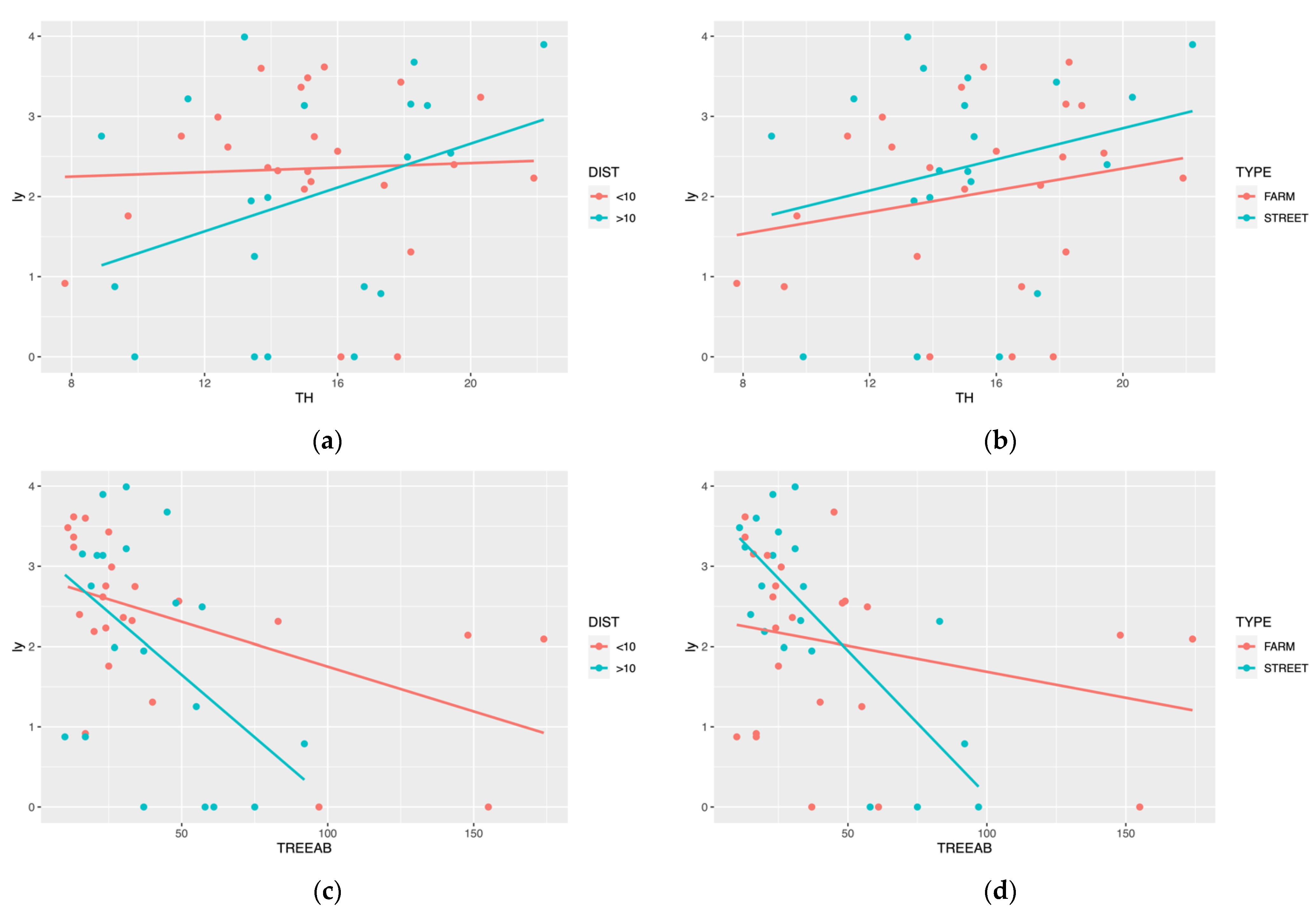
3.3. Correlation of Forests Stand Attributes and ALBs
4. Discussion
4.1. Landscape-Level Matrices and ALBs
4.2. Forest Patch Metrics and ALBs
4.3. Forest Stand Attributes and ALBs
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; Invasive Species Specialist Group: Auckland, New Zealand, 2000; Volume 12. [Google Scholar]
- Haack, R.A.; Herard, F.; Sun, J.; Turgeon, J.J. Managing invasive populations of Asian longhorned beetle and citrus longhorned beetle: A worldwide perspective. Annu. Rev. Entomol. 2010, 55, 521–546. [Google Scholar] [CrossRef] [Green Version]
- Meng, P.S.; Hoover, K.; Keena, M.A. Asian Longhorned Beetle (Coleoptera: Cerambycidae), an Introduced Pest of Maple and Other Hardwood Trees in North America and Europe. J. Integr. Pest Manag. 2015, 6, 1–13. [Google Scholar] [CrossRef]
- Lingafelter, S.W.; Hoebeke, E.R. Revision of the Genus Anoplophora (Coleoptera: Cerambycidae); Entomological Society of Washington: Washington, DC, USA, 2002; ISBN 0-9720714-1-5. [Google Scholar]
- Wang, Z. Study on the Occurrence Dynamics of Anoplophora glabripennis (Coleoptera: Cerambycidae) and Its Control Measures. PhD Thesis, Northeast Forestry University, Harbin, China, 2004. [Google Scholar]
- Luo, Y.; Liu, R.; Xu, Z. Theories and technologies of ecologically regulating poplar longhorned beetle disaster in shelter forest. J. Beijing For. Univ. 2002, 24, 160–164. [Google Scholar]
- Javal, M.; Roques, A.; Haran, J.; Hérard, F.; Keena, M.; Roux, G. Complex invasion history of the Asian long-horned beetle: Fifteen years after first detection in Europe. J. Pest Sci. 2017, 92, 173–187. [Google Scholar] [CrossRef]
- Hu, J.; Angeli, S.; Schuetz, S.; Luo, Y.; Hajek, A.E. Ecology and management of exotic and endemic Asian longhorned beetle Anoplophora glabripennis. Agric. For. Entomol. 2009, 11, 359–375. [Google Scholar] [CrossRef]
- Golec, J.R.; Li, F.; Cao, L.M.; Wang, X.Y.; Duan, J.J. Mortality factors of Anoplophora glabripennis (Coleoptera: Cerambycidae) infesting Salix and Populus in central, northwest, and northeast China. Biol. Control 2018, 126, 198–208. [Google Scholar] [CrossRef]
- Haack, R.A. Exotic bark-and wood-boring Coleoptera in the United States: Recent establishments and interceptions. Can. J. For. Res. 2006, 36, 269–288. [Google Scholar] [CrossRef]
- Bartell, S.M.; Nair, S.K. Establishment risks for invasive species. Risk Anal. 2004, 24, 833–845. [Google Scholar] [CrossRef] [PubMed]
- Nowak, D.J.; Pasek, J.E.; Sequeira, R.A.; Crane, D.E.; Mastro, V.C. Potential effect of Anoplophora glabripennis (Coleoptera: Cerambycidae) on urban trees in the United States. J. Econ. Entomol. 2001, 94, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Hérard, F.; Ciampitti, M.; Maspero, M.; Krehan, H.; Benker, U.; Boegel, C.; Schrage, R.; Bouhot-Delduc, L.; Bialooki, P. Anoplophora species in Europe: Infestations and management processes 1. EPPO Bull. 2006, 36, 470–474. [Google Scholar] [CrossRef]
- Hua, L.; Li, S.; Zhang, X. Coleoptera, Cerambycidae. In Iconography of Forest Insects Hunan China; Peng, J., Liu, W., Eds.; Hunan Scientific and Technical Publishing House: Changsha, China, 1992; pp. 467–524. [Google Scholar]
- Yang, Z.; Wang, X.; Yao, W.; Chu, X.; Li, P. Generation differentiation and effective accumulated temperature of Anoplophora glabripennis (Motsch.). For. Pest Dis. 2000, 19, 12–14. [Google Scholar]
- Keena, M.A. Effects of temperature on Anoplophora glabripennis (Coleoptera: Cerambycidae) adult survival, reproduction, and egg hatch. Environ. Entomol. 2006, 35, 912–921. [Google Scholar] [CrossRef]
- Keena, M.A.; Moore, P.M. Effects of temperature on Anoplophora glabripennis (Coleoptera: Cerambycidae) larvae and pupae. Env. Entomol 2010, 39, 1323–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, V.; Keena, M.A. Development of the teneral adult Anoplophora glabripennis (Coleoptera: Cerambycidae): Time to initiate and completely bore out of maple wood. Env. Entomol 2013, 42, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Du, W.; Li, M. Global warming and adaptive changes of forest pests. J. Northwest For. Univ. 2011, 26, 124–128. [Google Scholar]
- Smith, M.T.; Tobin, P.C.; Bancroft, J.; Li, G.H.; Gao, R.T. Dispersal and spatiotemporal dynamics of Asian longhorned beetle (Coleoptera: Cerambycidae) in China. Environ. Entomol. 2004, 33, 435–442. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.W.; Li, G.; Gao, R. Tracking movements of individual Anoplophora glabripennis (Coleoptera: Cerambycidae) adults: Application of harmonic radar. Environ. Entomol. 2004, 33, 644–649. [Google Scholar] [CrossRef]
- Colunga-Garcia, M.; Haack, R.A.; Magarey, R.A.; Margosian, M.L. Modeling spatial establishment patterns of exotic forest insects in urban areas in relation to tree cover and propagule pressure. J. Econ. Entomol. 2010, 103, 108–118. [Google Scholar] [CrossRef]
- Huang, J.; Lu, X.; Liu, H.; Zong, S. The Driving Forces of Anoplophora glabripennis Have Spatial Spillover Effects. Forests 2021, 12, 1678. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Jactel, H.; Parrotta, J.A.; Ferraz, S.F. Role of eucalypt and other planted forests in biodiversity conservation and the provision of biodiversity-related ecosystem services. For. Ecol. Manag. 2013, 301, 43–50. [Google Scholar] [CrossRef]
- Lu, Z.; Ouyang, F.; Zhang, Y.; Guan, X.; Men, X. Impacts of landscape patterns on populations of the wheat mites, Petrobia latens (Müller) and Penthaleus major (Duges), in the North China Plain. Acta Ecol. Sin. 2016, 36, 4447–4455. [Google Scholar] [CrossRef]
- Bianchi, F.J.J.A.; Booij, C.J.H.; Tscharntke, T. Sustainable pest regulation in agricultural landscapes: A review on landscape composition, biodiversity and natural pest control. Proc. R. Soc. B Biol. Sci. 2006, 273, 1715–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, F.; Ge, F. Effects of agricultural landscape patterns on insects. Chin. J. Appl. Entomol. 2011, 48, 1177–1183. [Google Scholar]
- Kowe, P.; Mutanga, O.; Dube, T. Advancements in the remote sensing of landscape pattern of urban green spaces and vegetation fragmentation. Int. J. Remote Sens. 2021, 42, 3797–3832. [Google Scholar] [CrossRef]
- Rodman, K.C.; Andrus, R.A.; Butkiewicz, C.L.; Chapman, T.B.; Gill, N.S.; Harvey, B.J.; Kulakowski, D.; Tutland, N.J.; Veblen, T.T.; Hart, S.J. Effects of Bark Beetle Outbreaks on Forest Landscape Pattern in the Southern Rocky Mountains, USA. Remote Sens. 2021, 13, 1089. [Google Scholar] [CrossRef]
- Pazúr, R.; Price, B.; Atkinson, P.M. Fine temporal resolution satellite sensors with global coverage: An opportunity for landscape ecologists. Landsc. Ecol. 2021, 36, 2199–2213. [Google Scholar] [CrossRef]
- Ferrenberg, S. Landscape Features and Processes Influencing Forest Pest Dynamics. Curr. Landsc. Ecol. Rep. 2016, 1, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Dominik, C.; Seppelt, R.; Horgan, F.G.; Settele, J.; Vaclavik, T. Landscape composition, configuration, and trophic interactions shape arthropod communities in rice agroecosystems. J. Appl. Ecol. 2018, 55, 2461–2472. [Google Scholar] [CrossRef]
- Seppelt, R.; Beckmann, M.; Ceausu, S.; Cord, A.F.; Gerstner, K.; Gurevitch, J.; Kambach, S.; Klotz, S.; Mendenhall, C.; Phillips, H.R.P.; et al. Harmonizing Biodiversity Conservation and Productivity in the Context of Increasing Demands on Landscapes. Bioscience 2016, 66, 890–896. [Google Scholar] [CrossRef]
- Fahrig, L.; Baudry, J.; Brotons, L.; Burel, F.G.; Crist, T.O.; Fuller, R.J.; Sirami, C.; Siriwardena, G.M.; Martin, J.L. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol Lett 2011, 14, 101–112. [Google Scholar] [CrossRef]
- McGarigal, K.; Cushman, S.A.; Neel, M.C.; Ene, E. FRAGSTATS: Spatial Pattern Analysis Program for Categorical and Continuous Maps. Available online: http://www.umass.edu/landeco/research/fragstats/fragstats.html (accessed on 10 May 2020).
- McGarigal, K. FRAGSTATS Help. Available online: http://www.umass.edu/landeco/research/fragstats/documents/fragstats.help.4.2.pdf (accessed on 21 April 2020).
- Syrbe, R.-U.; Walz, U. Spatial indicators for the assessment of ecosystem services: Providing, benefiting and connecting areas and landscape metrics. Ecol. Indic. 2012, 21, 80–88. [Google Scholar] [CrossRef]
- Badora, K.; Wróbel, R. Changes in the Spatial Structure of the Landscape of Isolated Forest Complexes in the 19th and 20th Centuries and Their Potential Effects on Supporting Ecosystem Services Related to the Protection of Biodiversity Using the Example of the Niemodlin Forests (SW Poland). Sustainability 2020, 12, 4237. [Google Scholar] [CrossRef]
- Perović, D.; Gámez-Virués, S.; Börschig, C.; Klein, A.-M.; Krauss, J.; Steckel, J.; Rothenwöhrer, C.; Erasmi, S.; Tscharntke, T.; Westphal, C.; et al. Configurational landscape heterogeneity shapes functional community composition of grassland butterflies. J. Appl. Ecol. 2015, 52, 505–513. [Google Scholar] [CrossRef]
- Montagnana, P.C.; Alves, R.S.C.; Garófalo, C.A.; Ribeiro, M.C. Landscape heterogeneity and forest cover shape cavity-nesting hymenopteran communities in a multi-scale perspective. Basic Appl. Ecol. 2021, 56, 239–249. [Google Scholar] [CrossRef]
- Galpern, P.; Best, L.R.; Devries, J.H.; Johnson, S.A. Wild bee responses to cropland landscape complexity are temporally-variable and taxon-specific: Evidence from a highly replicated pseudo-experiment. Agric. Ecosyst. Environ. 2021, 322, 107652. [Google Scholar] [CrossRef]
- Coutinho, J.G.E.; Hipólito, J.; Santos, R.L.S.; Moreira, E.F.; Boscolo, D.; Viana, B.F. Landscape Structure Is a Major Driver of Bee Functional Diversity in Crops. Front. Ecol. Evol. 2021, 9, 624835. [Google Scholar] [CrossRef]
- Schirmel, J.; Albrecht, M.; Bauer, P.M.; Sutter, L.; Pfister, S.C.; Entling, M.H. Landscape complexity promotes hoverflies across different types of semi-natural habitats in farmland. J. Appl. Ecol. 2018, 55, 1747–1758. [Google Scholar] [CrossRef]
- Holdenrieder, O.; Pautasso, M.; Weisberg, P.J.; Lonsdale, D. Tree diseases and landscape processes: The challenge of landscape pathology. Trends Ecol. Evol. 2004, 19, 446–452. [Google Scholar] [CrossRef]
- Meentemeyer, R.K.; Haas, S.E.; Václavík, T. Landscape epidemiology of emerging infectious diseases in natural and human-altered ecosystems. Annu. Rev. Phytopathol. 2012, 50, 379–402. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Tian, C.; Sun, J. Effects of landscape complexity and stand factors on arthropod communities in poplar forests. Ecol. Evol. 2019, 9, 7143–7156. [Google Scholar] [CrossRef] [Green Version]
- Lantschner, M.V.; Corley, J.C. Spatial Pattern of Attacks of the Invasive Woodwasp Sirex noctilio, at Landscape and Stand Scales. PLoS ONE 2015, 10, e0127099. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S; Springer: New York, NY, USA, 2002; ISBN 0-387-95457-0. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; ISBN 978-1-5443-3647-3. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Pinheiro, J.; Bates, D. Mixed-Effects Models in S and S-PLUS; Springer: New York, NY, USA, 2000; ISBN 0-387-98957-9. [Google Scholar]
- McGarigal, K. Landscape Metrics for Categorical Map Patterns. Available online: http://www.umass.edu/landeco/teaching/landscape_ecology/schedule/chapter9_metrics.pdf (accessed on 3 July 2020).
- Wen, J.; Li, Y.; Xia, N.; Luo, Y. Study on dispersal pattern of Anoplophora glabripennis adults in poplars. Acta Ecol. Sin. 1998, 18, 269–277. [Google Scholar]
- Ronce, O. How does it feel to be like a rolling stone? Ten questions about dispersal evolution. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 231–253. [Google Scholar] [CrossRef]
- Mazzi, D.; Dorn, S. Movement of insect pests in agricultural landscapes. Ann. Appl. Biol. 2012, 160, 97–113. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef] [Green Version]
- Ryser, R.; Hirt, M.R.; Haussler, J.; Gravel, D.; Brose, U. Landscape heterogeneity buffers biodiversity of simulated meta-food-webs under global change through rescue and drainage effects. Nat. Commun. 2021, 12, 4716. [Google Scholar] [CrossRef]
- Taylor, P.D.; Fahrig, L.; Henein, K.; Merriam, G. Connectivity is a vital element of landscape structure. Oikos 1993, 68(3), 571–573. [Google Scholar] [CrossRef] [Green Version]
- Ziv, Y.; Davidowitz, G. When Landscape Ecology Meets Physiology: Effects of Habitat Fragmentation on Resource Allocation Trade-Offs. Front. Ecol. Evol. 2019, 7, 137. [Google Scholar] [CrossRef] [Green Version]
- Slattery, Z.; Fenner, R. Spatial Analysis of the Drivers, Characteristics, and Effects of Forest Fragmentation. Sustainability 2021, 13, 3246. [Google Scholar] [CrossRef]
- Stamps, J.A.; Buechner, M.; Krishnan, V.V. The Effects of Edge Permeability and Habitat Geometry on Emigration from Patches of Habitat. Am. Nat. 1987, 129, 533–552. [Google Scholar] [CrossRef]
- Collinge, S.K.; Palmer, T.M. The influences of patch shape and boundary contrast on insect response to fragmentation in California grasslands. Landsc. Ecol. 2002, 17, 647–656. [Google Scholar] [CrossRef]
- Su, Z.; Li, X.; Zhou, W.; Ouyang, Z. Effect of landscape pattern on insect species density within urban green spaces in Beijing, China. PLoS ONE 2015, 10, e0119276. [Google Scholar] [CrossRef]
- Larsen, A.E.; Noack, F. Impact of local and landscape complexity on the stability of field-level pest control. Nat. Sustain. 2020, 4, 120–128. [Google Scholar] [CrossRef]
- Wang, X.; Ouyang, S.; Sun, O.J.; Fang, J. Forest biomass patterns across northeast China are strongly shaped by forest height. For. Ecol. Manag. 2013, 293, 149–160. [Google Scholar] [CrossRef]
- Hui, D.; Wang, J.; Le, X.; Shen, W.; Ren, H. Influences of biotic and abiotic factors on the relationship between tree productivity and biomass in China. For. Ecol. Manag. 2012, 264, 72–80. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Landscape Level | |||||
|---|---|---|---|---|---|
| Variable | Estimate | p-Value | R2 (Model) | Adj-R2 (Model) | p-Value (Model) |
| LPI | −1.6686 | 0.0087 ** | 0.7873 | 0.6455 | 0.0324 * |
| GYRATE_MN | 1.2231 | 0.0087 * | |||
| SHAPE_MN | −1.9686 | 0.0153 * | |||
| SHDI | −2.1956 | 0.0187 * | |||
| Class Level: Forest | |||||
| LPI | −0.5809 | 0.0206 * | 0.5979 | 0.4973 | 0.0262 * |
| AI | 0.6520 | 0.0121 * |
| Model | AIC | |||||||
|---|---|---|---|---|---|---|---|---|
| Model 1 | TH | DBH | TCW | TREEAB | TSD | (1|TYPE) | (1|DIST) | 165.4574 |
| p-value | 0.0701 | 0.0095 ** | ||||||
| Estimate | 0.1225 | −0.0145 | ||||||
| Model 2 | TH | DBH | TCW | TREEAB | TSD | (1|TYPE) | 163.5127 | |
| p-value | 0.0465 * | 0.0083 ** | ||||||
| Estimate | 0.1312 | −0.0148 | ||||||
| Model 3 | TH | DBH | TCW | TREEAB | TSD | (1| DIST) | 163.4574 | |
| p-value | 0.0701 | 0.0095 ** | ||||||
| Estimate | 0.1225 | −0.0145 | ||||||
| Model 4 | TH | TREEAB | (1|TYPE) | (1|DIST) | 153.5881 | |||
| p-value | 0.0490 * | 0.0011 ** | ||||||
| Estimate | 0.0966 | −0.0147 | ||||||
| Model 5 | TH | TREEAB | (1|TYPE) | |||||
| p-value | 0.0498 * | 0.0014 ** | 151.8876 | |||||
| Estimate | 0.0976 | −0.0145 | ||||||
| Model 6 | TH | TREEAB | (1|DIST) | 151.5881 | ||||
| p-value | 0.0490* | 0.0011 ** | ||||||
| Estimate | 0.0966 | −0.0147 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.; Zhan, Z.; Zong, S.; Ren, L. The Relationship between Landscape Patterns and Populations of Asian Longhorned Beetles. Forests 2022, 13, 1981. https://doi.org/10.3390/f13121981
Yang C, Zhan Z, Zong S, Ren L. The Relationship between Landscape Patterns and Populations of Asian Longhorned Beetles. Forests. 2022; 13(12):1981. https://doi.org/10.3390/f13121981
Chicago/Turabian StyleYang, Chao, Zhongyi Zhan, Shixiang Zong, and Lili Ren. 2022. "The Relationship between Landscape Patterns and Populations of Asian Longhorned Beetles" Forests 13, no. 12: 1981. https://doi.org/10.3390/f13121981
APA StyleYang, C., Zhan, Z., Zong, S., & Ren, L. (2022). The Relationship between Landscape Patterns and Populations of Asian Longhorned Beetles. Forests, 13(12), 1981. https://doi.org/10.3390/f13121981