

Plastome Characterization, Phylogenetic Relationships, and Regional Conservation Status of Ficus populifolia Vahl. (Moraceae), a Peripherally Isolated Plant Population in the Arabian Peninsula

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plastome Comparative Analysis
2.1.1. Plant Sampling
2.1.2. DNA Extraction, Library Construction, and Genome Sequencing
2.1.3. Plastome Assembly and Annotation
2.1.4. Codon Usage and RNA Editing Sites
2.1.5. Repeat Analysis in the Plastome
2.1.6. Sequence Divergence and Boundary
2.1.7. Characterization of the Substitution Rate
2.1.8. Phylogenomic Analysis
2.2. Conservation Assessment
3. Results and Discussion
3.1. Plastome Characterization
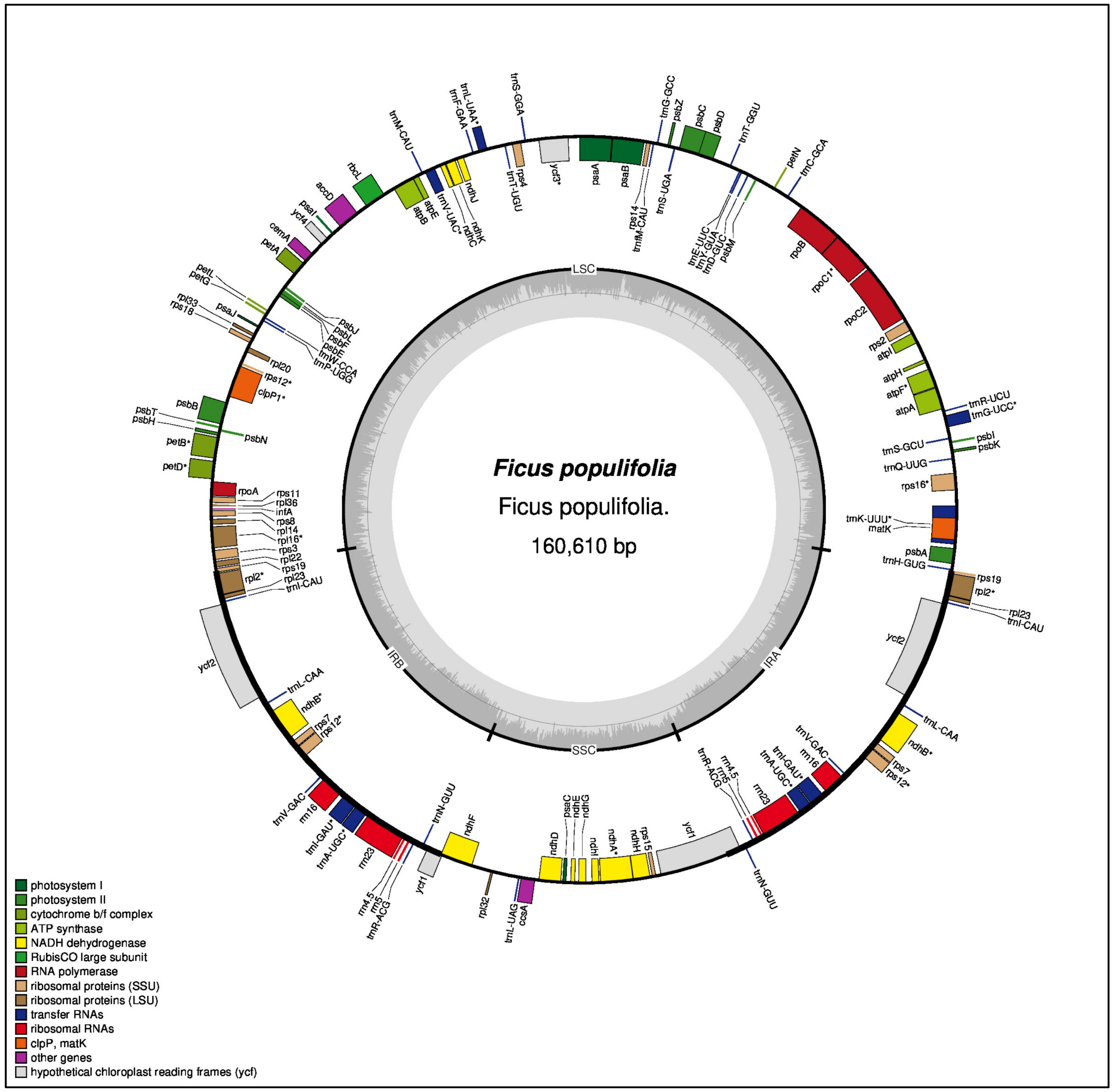
3.1.1. General Characteristics of the Ficus populifolia Plastome
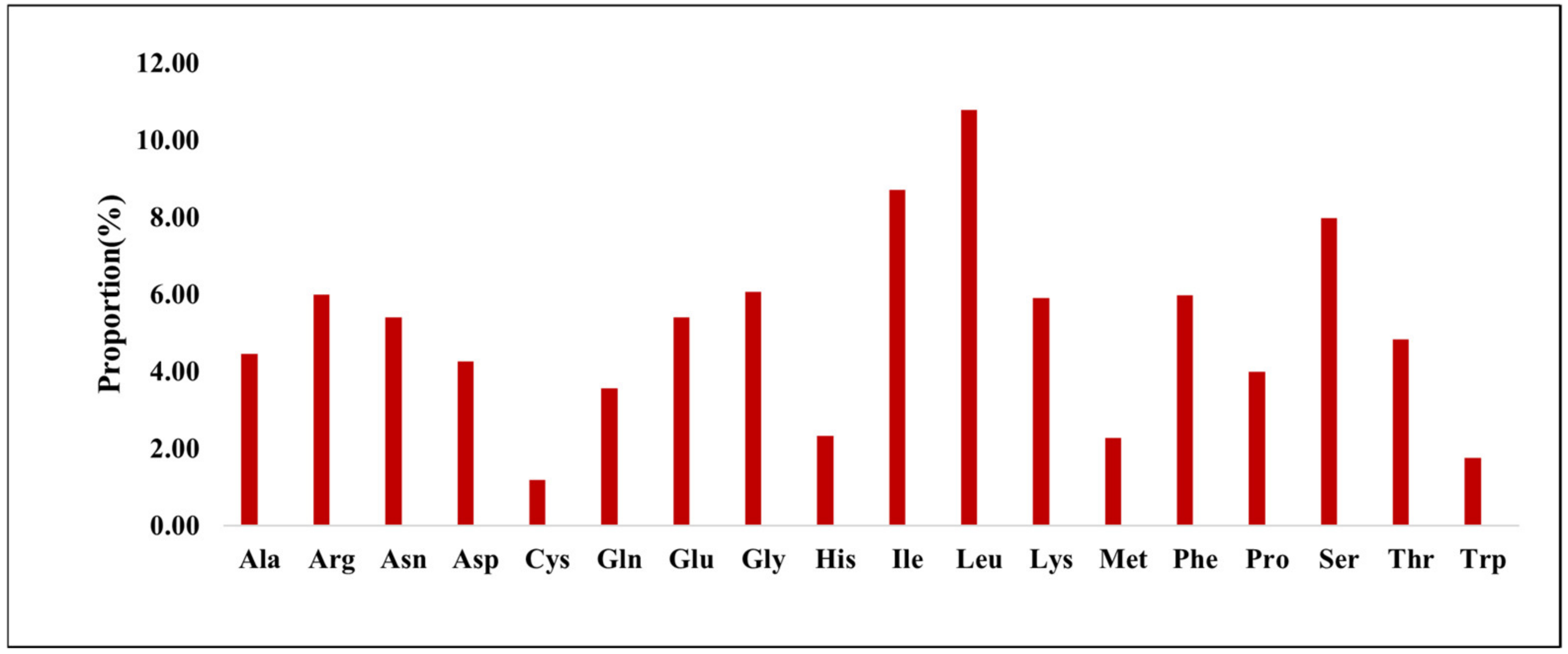
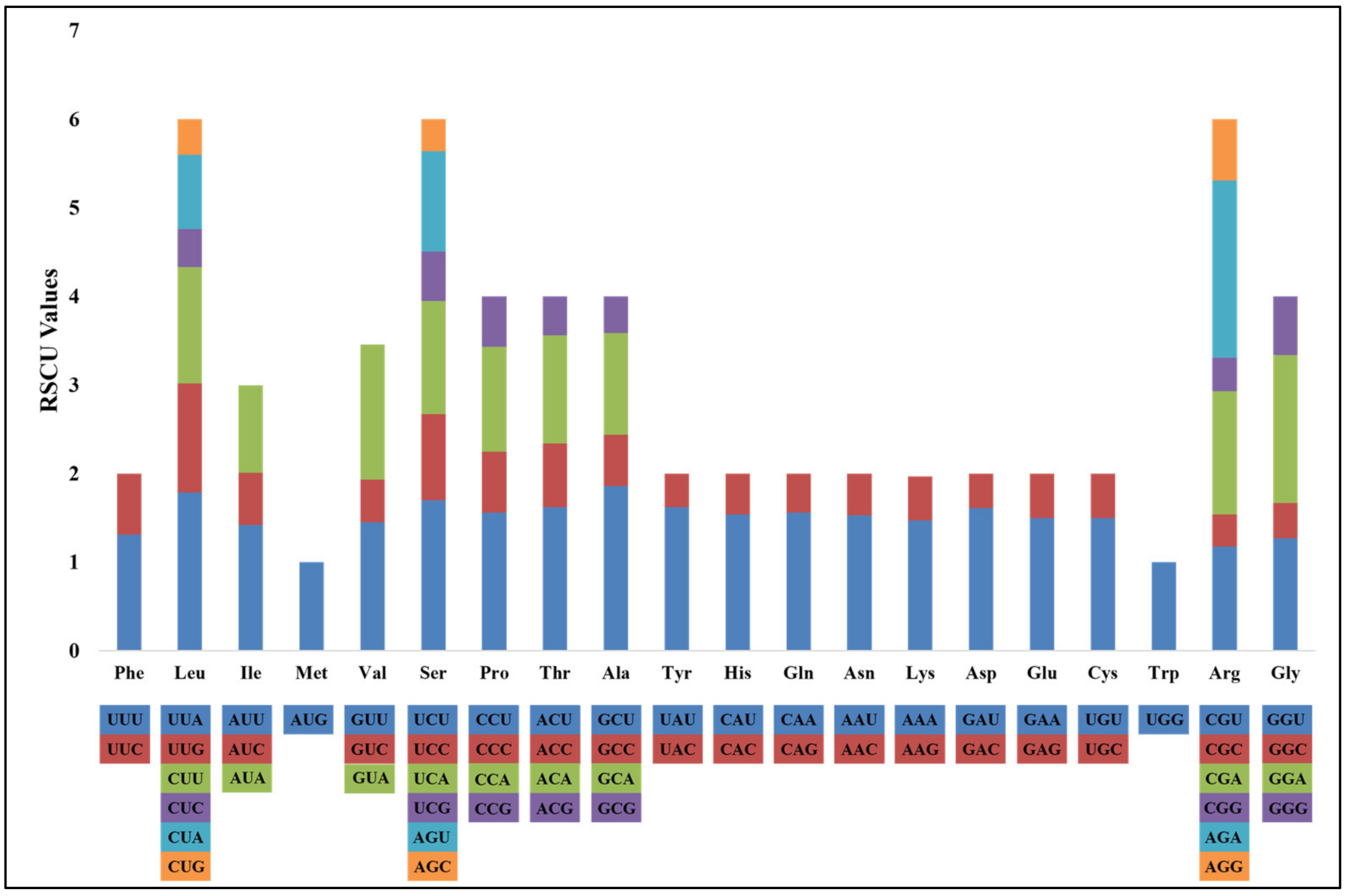
3.1.2. Codon Usage Bias
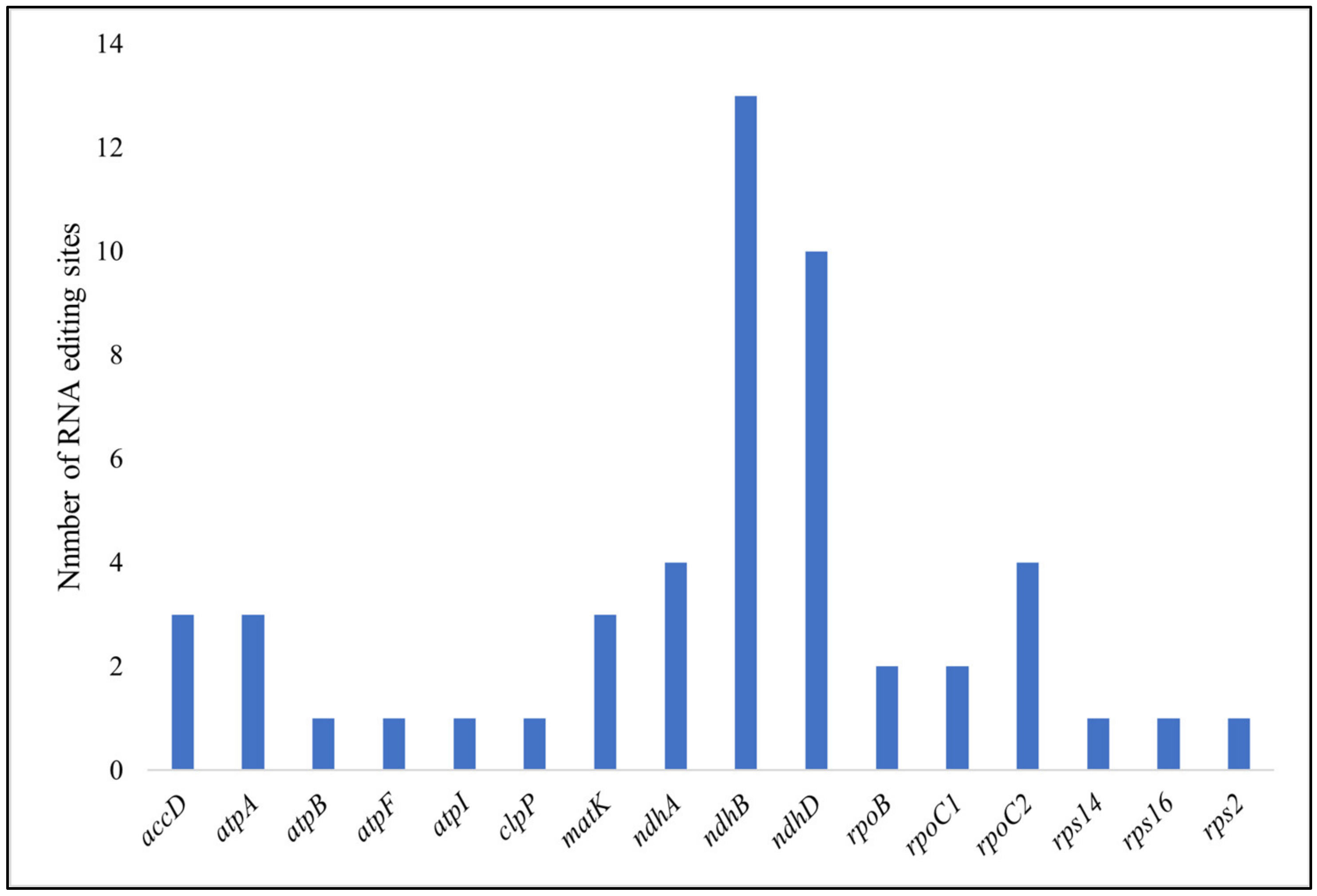
3.1.3. RNA Editing Sites
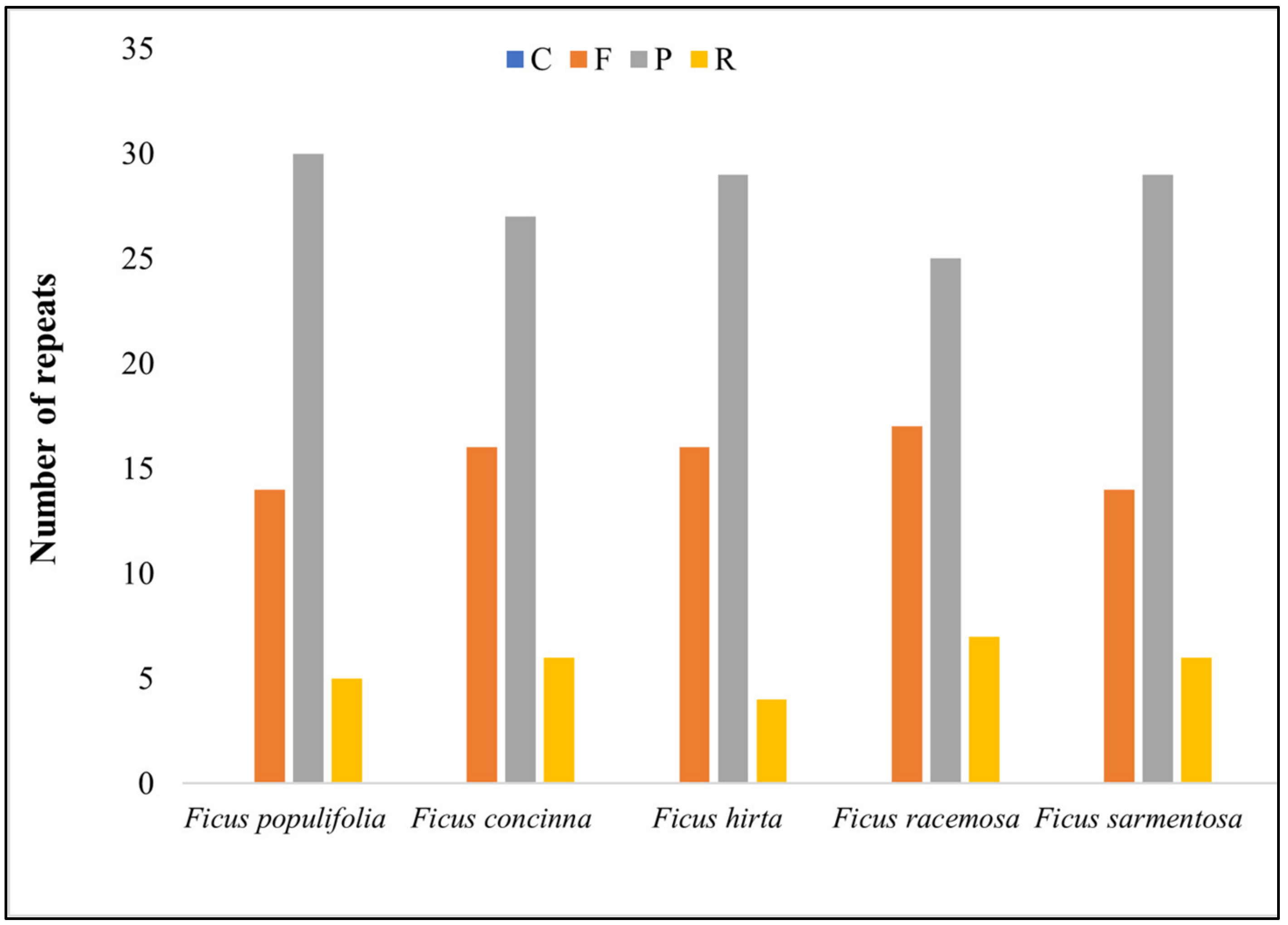
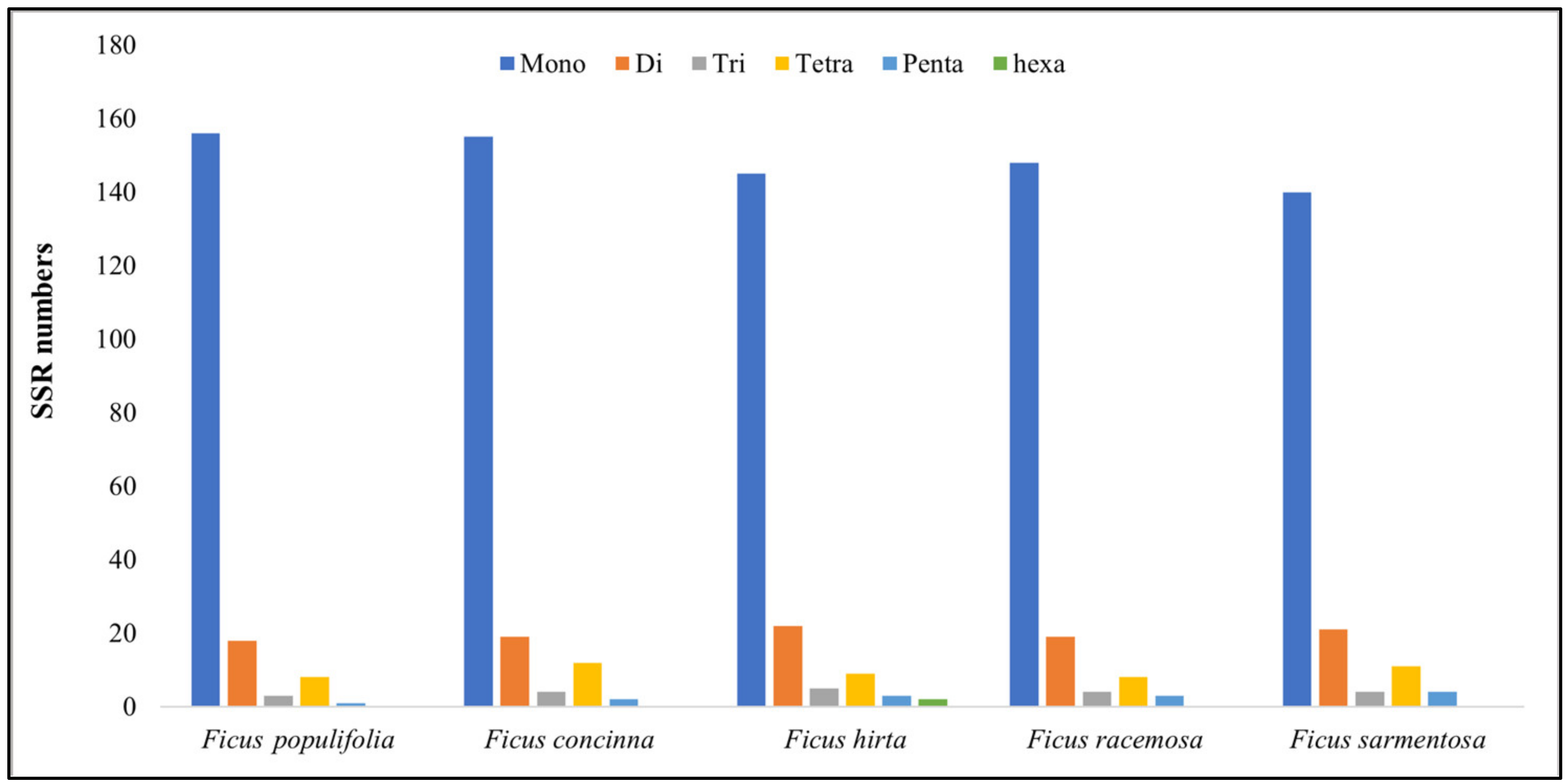
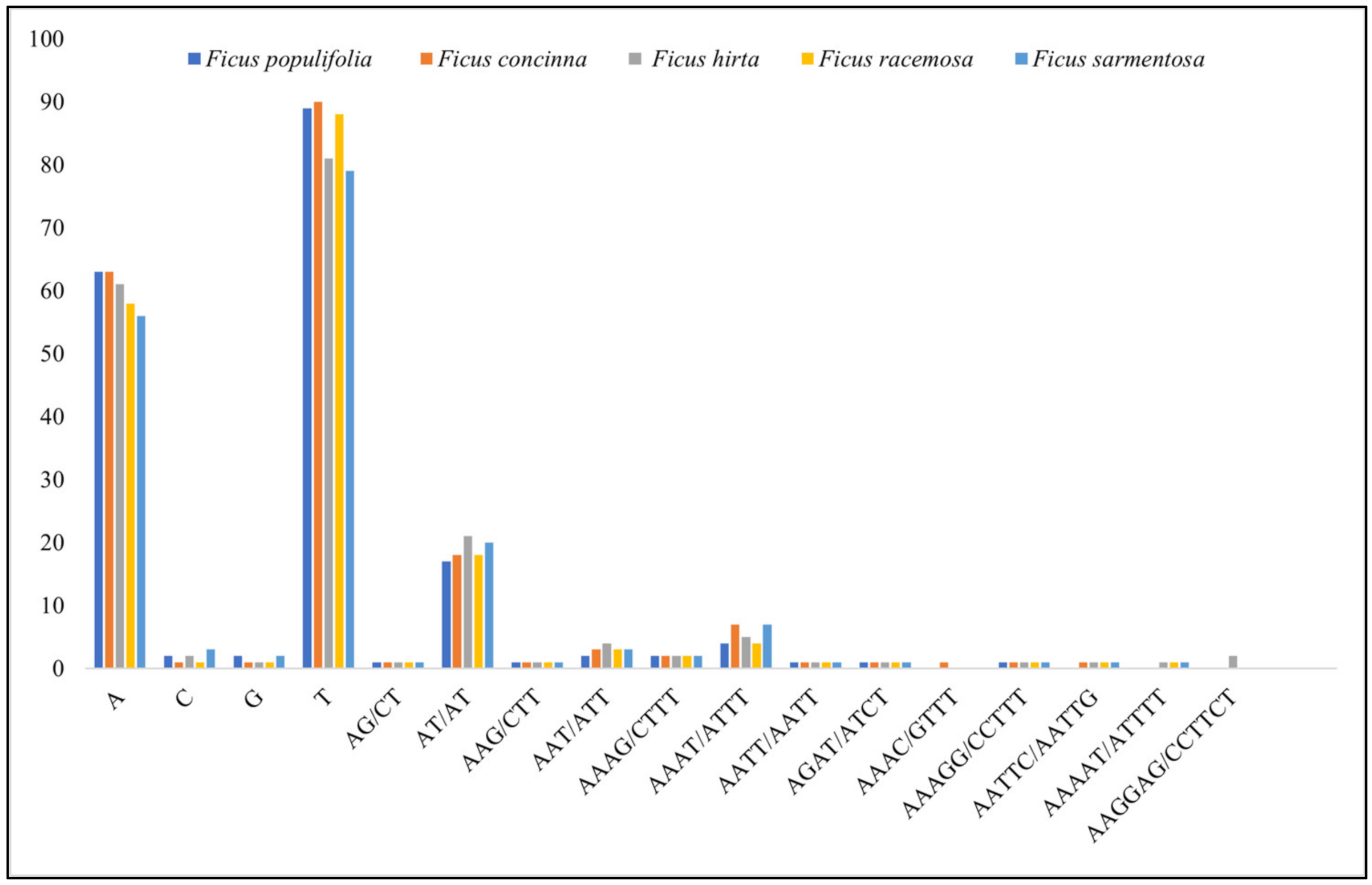
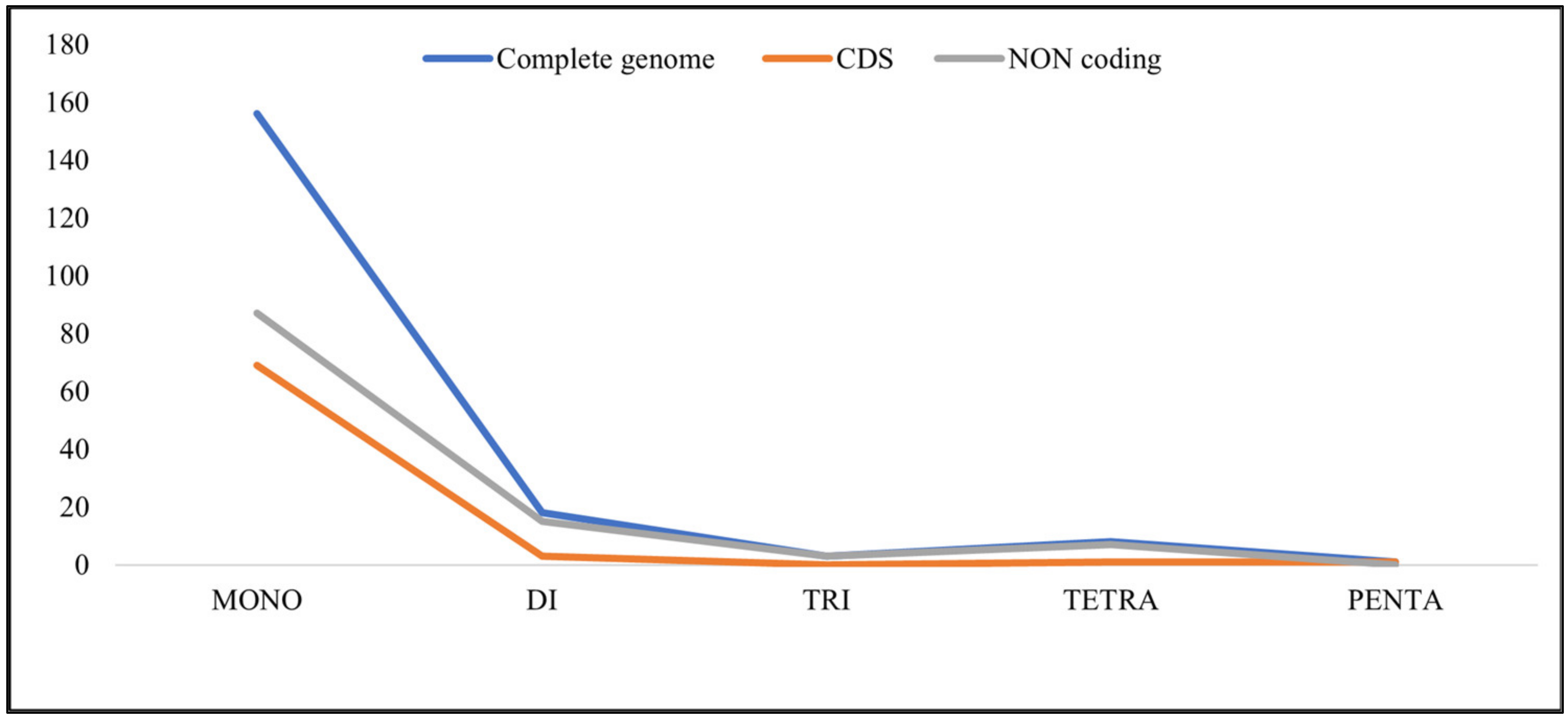
3.1.4. Repeat Analysis
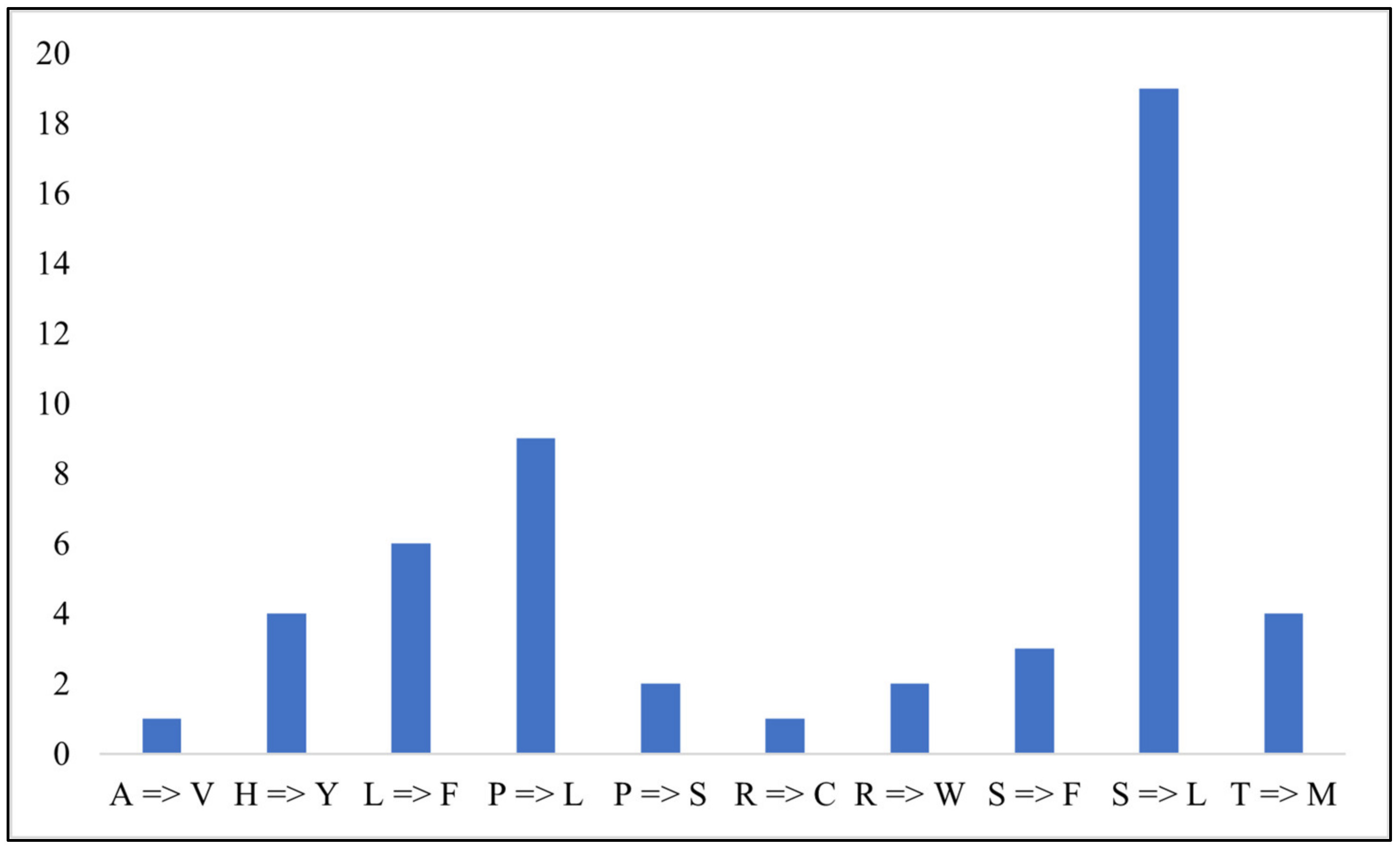
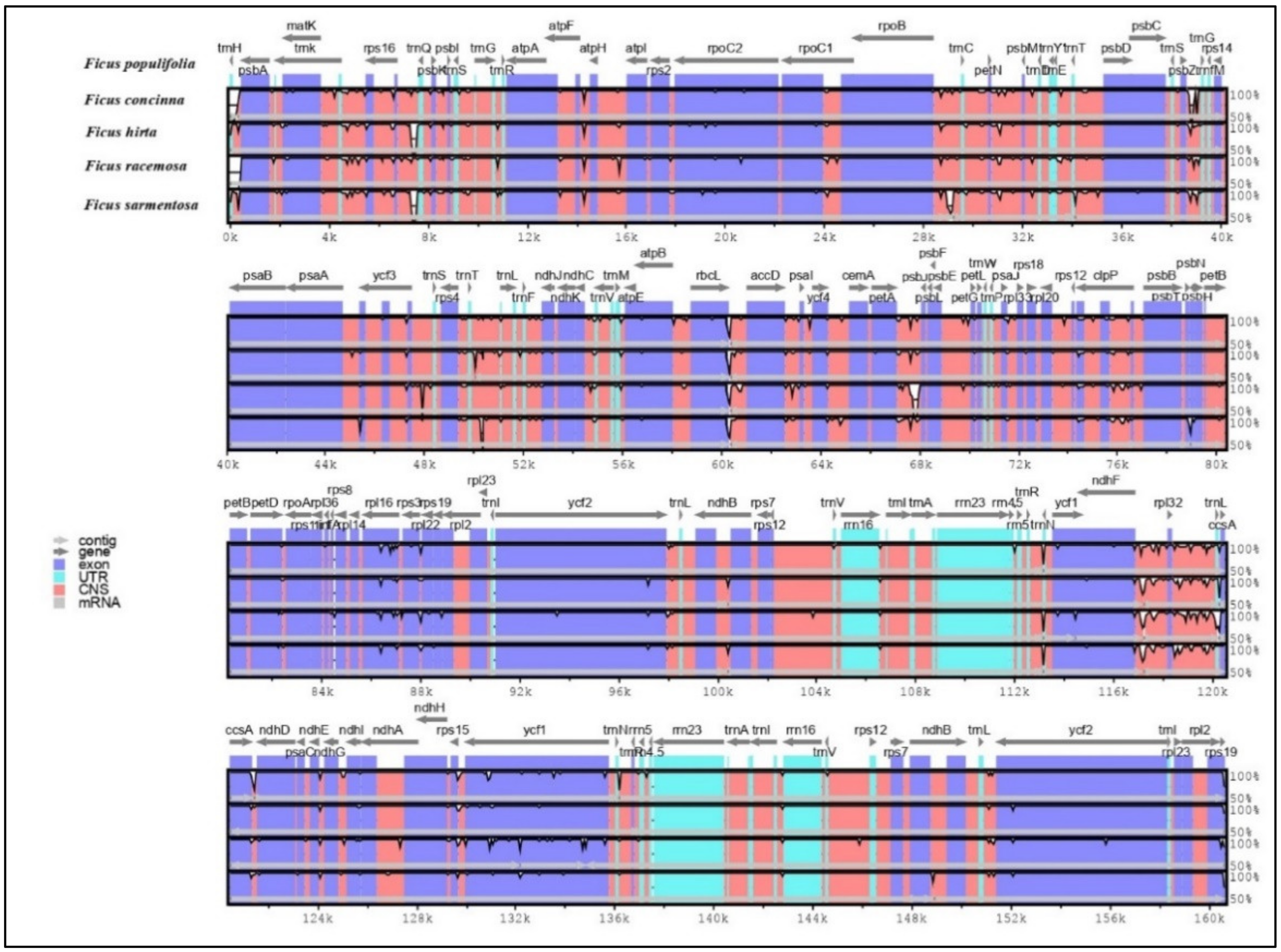
3.1.5. Sequence Divergence
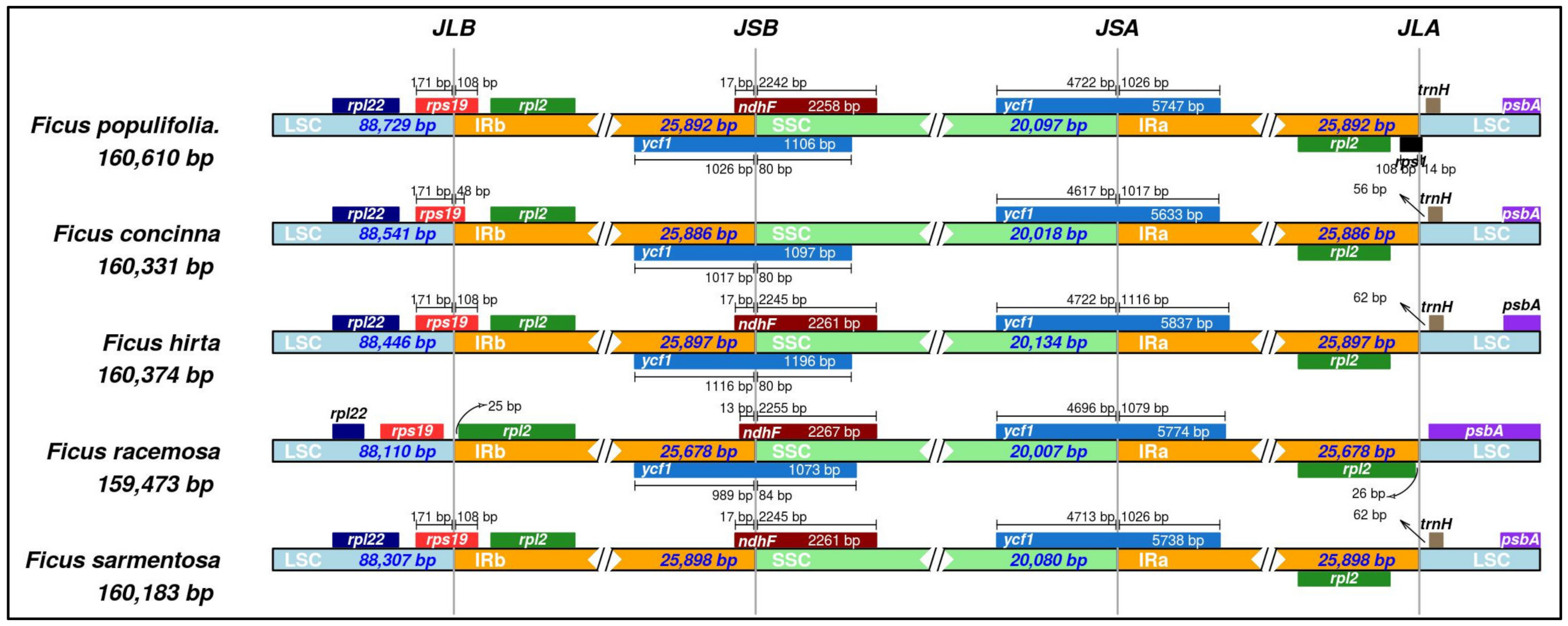
3.1.6. Contraction and Expansion of the Inverted Repeat Region
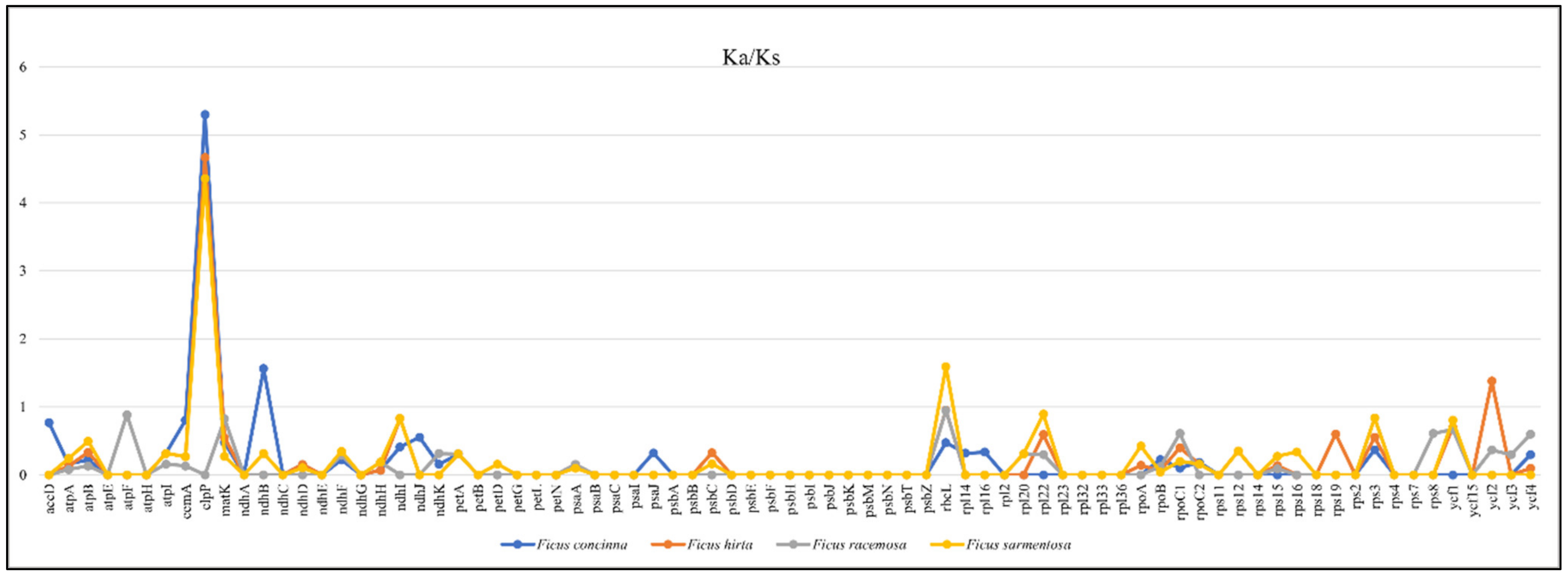
3.1.7. Characterization of Substitution Rate
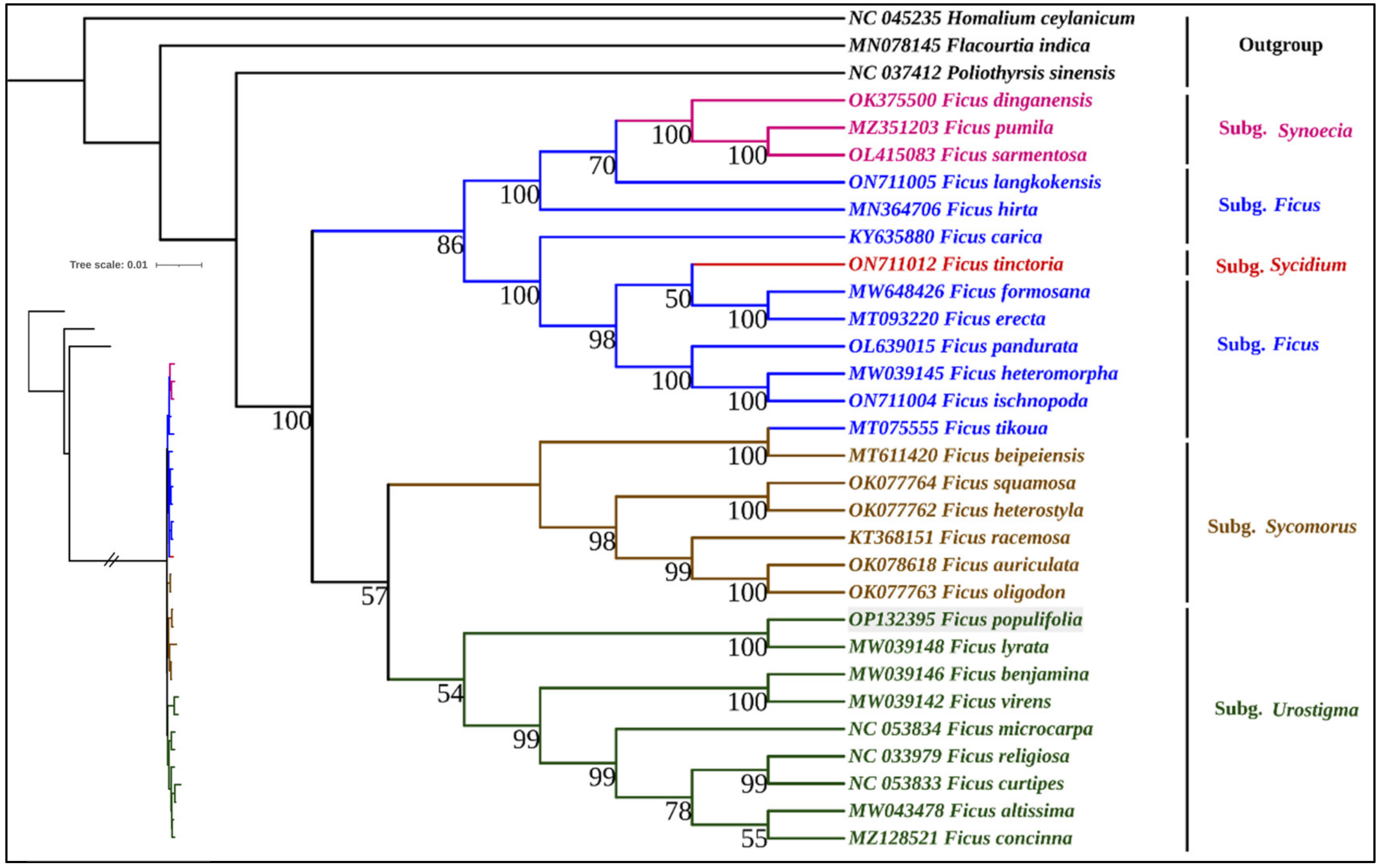
3.1.8. Phylogenomic Analysis
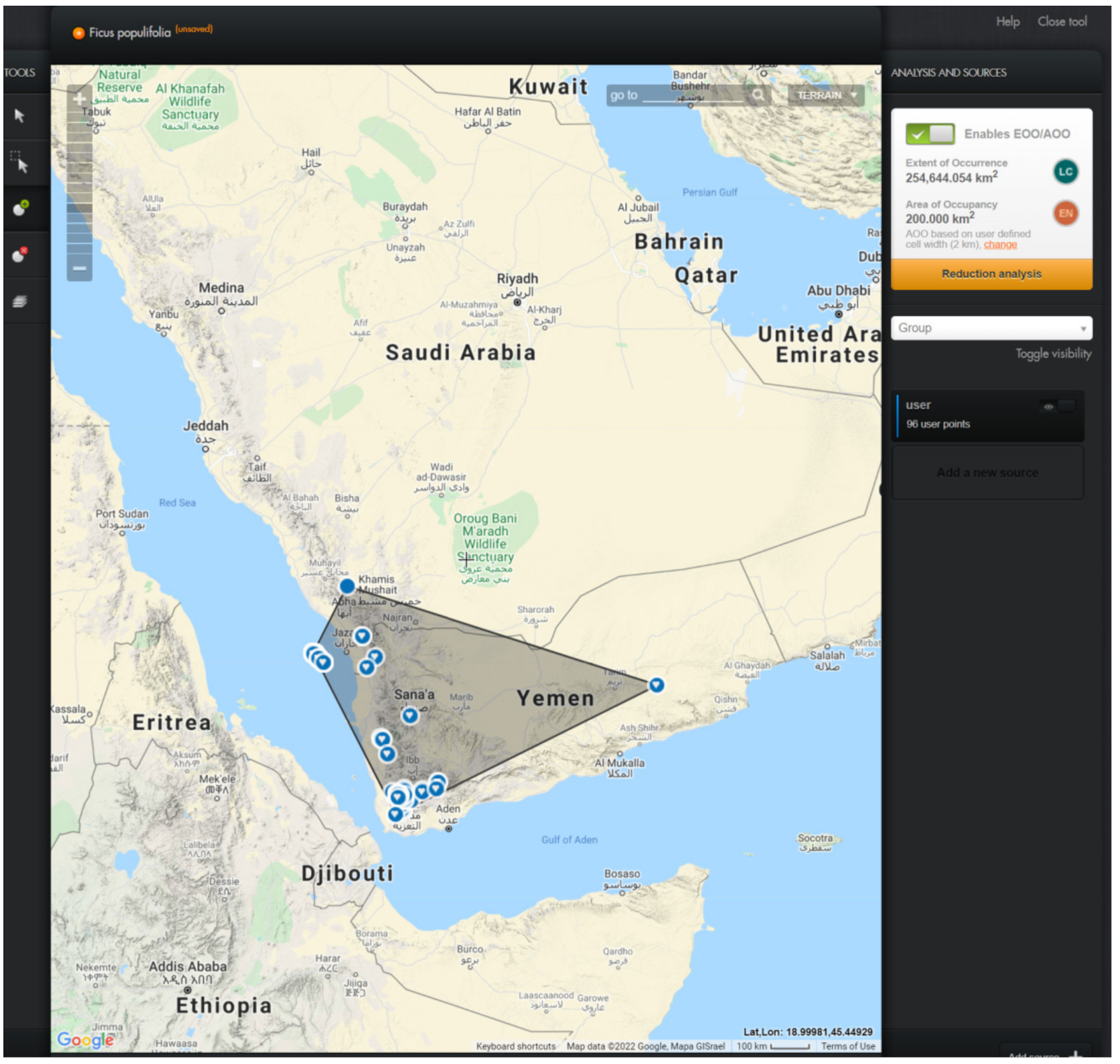
3.2. Conservation Assessment
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Berg, C.C.; Corner, E.J.H. Flora Malesiana. Series I, Seed Plants. Volume 17, Part 2: Moraceae (Ficus); Nationaal Herbarium Nederland: Leiden, The Netherlands, 2005; Volume 17. [Google Scholar]
- Cruaud, A.; Rønsted, N.; Chantarasuwan, B.; Chou, L.S.; Clement, W.L.; Couloux, A.; Cousins, B.; Genson, G.; Harrison, R.D.; Hanson, P.E. An extreme case of plant–insect codiversification: Figs and fig-pollinating wasps. Syst. Biol. 2012, 61, 1029–1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, R.D. Figs and the Diversity of Tropical Rainforests. BioScience 2005, 55, 1053–1064. [Google Scholar] [CrossRef]
- Ronsted, N.; Salvo, G.; Savolainen, V. Biogeographical and phylogenetic origins of African fig species (Ficus section Galoglychia). Mol. Phylogenet. Evol. 2007, 43, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.G.; Cope, T.A. Flora of the Arabian Peninsula and Socotra Volume 1; Edinburgh University Press in Association with Royal Botanic Garden: Edinburgh, UK, 1996; Volume 1. [Google Scholar]
- Beentje, H.J. Fig Trees (Ficus, Moraceae) of Kenya; East Africa Natural History Soc.: Nairobi, Kenya, 1988; Volume 76. [Google Scholar]
- Aweke, G. Revision of the Genus Ficus L. (Moraceae) in Ethiopia; (Primitiae Africanae XI); Meded. Landbouwhogeschool: Wageningen, The Netherlands, 1979; pp. 79–83. [Google Scholar]
- Quattrocchi, U. CRC World Dictionary of Medicinal and Poisonous Plants: Common Names, Scientific Names, Eponyms, Synonyms, and Etymology; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Abeli, T.; Gentili, R.; Rossi, G.; Bedini, G.; Foggi, B. Can the IUCN criteria be effectively applied to peripheral isolated plant populations? Biodivers. Conserv. 2009, 18, 3877–3890. [Google Scholar] [CrossRef]
- Zhou, R.; Ling, S.; Zhao, W.; Osada, N.; Chen, S.; Zhang, M.; He, Z.; Bao, H.; Zhong, C.; Zhang, B.; et al. Population genetics in nonmodel organisms: II. Natural selection in marginal habitats revealed by deep sequencing on dual platforms. Mol. Biol. Evol. 2011, 28, 2833–2842. [Google Scholar]
- Arnaud-Haond, S.; Teixeira, S.; Massa, S.I.; Billot, C.; Saenger, P.; Coupland, G.; Duarte, C.M.; Serrão, E.A. Genetic structure at range edge: Low diversity and high inbreeding in Southeast Asian mangrove (Avicennia marina) populations. Mol. Ecol. 2006, 15, 3515–3525. [Google Scholar] [CrossRef]
- Fady, B.; Aravanopoulos, F.A.; Alizoti, P.; Mátyás, C.; von Wühlisch, G.; Westergren, M.; Belletti, P.; Cvjetkovic, B.; Ducci, F.; Huber, G. Evolution-based approach needed for the conservation and silviculture of peripheral forest tree populations. For. Ecol. Manag. 2016, 100, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Alleaume-Benharira, M.; Pen, I.R.; Ronce, O. Geographical patterns of adaptation within a species’ range: Interactions between drift and gene flow. J. Evol. Biol. 2006, 19, 203–215. [Google Scholar] [CrossRef]
- UNDP. National Biodiversity Strategy and Action Plan II “Achieving a Resilient, Productive and Sustainable Socio-Ecosystem by 2050”; Ministry of Water and Environment and Environment Protection Authority (EPA): Yemen, 2017; Available online: https://andp.unescwa.org/sites/default/files/2020-10/National%20Biodiversity%20Strategy%20and%20Action%20Plan%202-achieving%20a%20resilient%2C%20productive%20and%20sustainable%20socio-ecosystem%20by%202050.pdf (accessed on 14 February 2022).
- Planta Europa. A Sustainable Future for Europe; The European Strategy for Plant Conservation 2008–2014; Plantlife International: Salisbury, UK; The Council of Europe: Strasbourg, France, 2008. [Google Scholar]
- Botanic Gardens Conservation International (BGCI); IUCN SSC Global Tree Specialist Group. Ficus Populifolia; The IUCN Red List of Threatened Species: Gland, Switzerland, 2019; e.T143276998A143294738. [Google Scholar] [CrossRef]
- IUCN Standards and Petitions Committee. Guidelines for Using the IUCN Red List Categories and Criteria. Version 15.1; Prepared by the Standards and Petitions Committee: Cambridge, UK, 2022. [Google Scholar]
- IUCN. Guidelines for Application of IUCN Red List Criteria at Regional and National Levels: Version 4.0; IUCN: Gland, Switzerland; Cambridge, UK, 2012. [Google Scholar]
- Li, X.; Yang, Y.; Henry, R.J.; Rossetto, M.; Wang, Y.; Chen, S. Plant DNA barcoding: From gene to genome. Biol. Rev. 2015, 90, 157–166. [Google Scholar] [CrossRef]
- Jansen, R.K.; Ruhlman, T.A. Plastid genomes of seed plants. In Genomics of Chloroplasts and Mitochondria; Springer: Berlin/Heidelberg, Germany, 2012; pp. 103–126. [Google Scholar]
- Van Noort, S.; Rasplus, J. Figweb: Figs and Fig Wasps of the World. Available online: www.figweb.org (accessed on 20 February 2022).
- Huang, Y.; Li, J.; Yang, Z.; An, W.; Xie, C.; Liu, S.; Zheng, X. Comprehensive analysis of complete chloroplast genome and phylogenetic aspects of ten Ficus species. BMC Plant Biol. 2022, 22, 253. [Google Scholar] [CrossRef]
- Xia, X.; Peng, J.; Yang, L.; Zhao, X.; Duan, A.; Wang, D. Comparative Analysis of the Complete Chloroplast Genomes of Eight Ficus Species and Insights into the Phylogenetic Relationships of Ficus. Life 2022, 12, 848. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.; Doyle, J. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phyotochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar] [CrossRef] [Green Version]
- Lohse, M.; Drechsel, O.; Bock, R. OrganellarGenomeDRAW (OGDRAW): A tool for the easy generation of high-quality custom graphical maps of plastid and mitochondrial genomes. Curr. Genet. 2007, 52, 267–274. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Mower, J.P. The PREP suite: Predictive RNA editors for plant mitochondrial genes, chloroplast genes and user-defined alignments. Nucleic Acids Res. 2009, 37, 253–259. [Google Scholar] [CrossRef]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef] [Green Version]
- Thiel, T.; Michalek, W.; Varshney, R.; Graner, A. Exploiting EST databases for the development and characterization of gene-derived SSR-markers in barley (Hordeum vulgare L.). Theor. Appl. Genet. 2003, 106, 411–422. [Google Scholar] [CrossRef]
- Mayor, C.; Brudno, M.; Schwartz, J.R.; Poliakov, A.; Rubin, E.M.; Frazer, K.A.; Pachter, L.S.; Dubchak, I. VISTA: Visualizing global DNA sequence alignments of arbitrary length. Bioinformatics 2000, 16, 1046. [Google Scholar] [CrossRef] [Green Version]
- Amiryousefi, A.; Hyvönen, J.; Poczai, P. IRscope: An online program to visualize the junction sites of chloroplast genomes. Bioinformatics 2018, 34, 3030–3031. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, Y.; Zhang, Z.; Zhu, J.; Yu, J. KaKs_Calculator 2.0: A Toolkit Incorporating Gamma-Series Methods and Sliding Window Strategies. Genom. Proteom. Bioinform. 2010, 8, 77–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Darriba, D.; Taboada, G.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akaike, H. A new look at the statistical model identification. In Selected Papers of Hirotugu Akaike; Springer: Berlin/Heidelberg, Germany, 1974; pp. 215–222. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree OF Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- El-Shabasy, A.; Kasem, W. Systematic composition, species diversity and plant chorology at Wadi Tashar, Jazan, Saudi Arabia. J. Med. Plants Stud. 2018, 6, 83–88. [Google Scholar]
- Gabali, S.A.; Al-Gifri, A.N. Flora of South Yemen—Angiospermae A provisional checklist. Feddes Repert. 1990, 101, 373–383. [Google Scholar] [CrossRef]
- Hamed, M.A.; Kasem, W.T.; Shalabi, L.F. Floristic diversity and vegetation-soil correlations in Wadi Qusai, Jazan, Saudi Arabia. Int. J. Plant Soil Sci. 2018, 1–18. [Google Scholar] [CrossRef]
- Kasem, W.T.; Marei, A. Floristic Compositions and its affinities to phytogeographical regions in Wadi Khulab of Jazan, Saudi Arabia. Int. J. Plant Soil Sci. 2017, 16, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bachman, S.; Moat, J.; Hill, A.W.; de Torre, J.; Scott, B. Supporting Red List threat assessments with GeoCAT: Geospatial conservation assessment tool. ZooKeys 2011, 150, 117–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IUCN. IUCN Red List Categories and Criteria: Version 3.1, 2nd ed.; IUCN: Gland, Switzerland; Cambridge, UK, 2012. [Google Scholar]
- Wicke, S.; Schneeweiss, G.M.; Depamphilis, C.W.; Müller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.H.; Hou, Z. The complete chloroplast genome sequence of Ficus concinna (Moraceae) from Sichuan province. Mitochondrial DNA B Resour 2022, 7, 390–392. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, W.; Li, F.; Li, C.; Xie, X.; Chao, Z.; Tian, E. The complete chloroplast genome sequence of Ficus hirta (Moraceae). Mitochondrial DNA Part B 2019, 4, 4041–4042. [Google Scholar] [CrossRef] [Green Version]
- Dobrogojski, J.; Adamiec, M.; Luciński, R. The chloroplast genome: A review. Acta Physiol. Plant. 2020, 42, 98. [Google Scholar] [CrossRef]
- Asaf, S.; Ahmad, W.; Al-Harrasi, A.; Khan, A.L. Uncovering the first complete plastome genomics, comparative analyses, and phylogenetic dispositions of endemic medicinal plant Ziziphus hajarensis (Rhamnaceae). BMC Genom. 2022, 23, 83. [Google Scholar] [CrossRef]
- Al-Juhani, W.S.; Alharbi, S.A.; Al Aboud, N.M.; Aljohani, A.Y. Complete chloroplast genome of the desert date (Balanites aegyptiaca (L.) Del. comparative analysis, and phylogenetic relationships among the members of Zygophyllaceae. BMC Genom. 2022, 23, 626. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Z.; Guo, Y.; Chen, S.; Xu, X.; Wang, R. Complete chloroplast genomes of Leptodermis scabrida complex: Comparative genomic analyses and phylogenetic relationships. Gene 2021, 791, 145715. [Google Scholar] [CrossRef]
- Li, C.-J.; Wang, R.-N.; Li, D.-Z. Comparative analysis of plastid genomes within the Campanulaceae and phylogenetic implications. PLoS ONE 2020, 15, e0233167. [Google Scholar] [CrossRef]
- Shaul, O. How introns enhance gene expression. Int. J. Biochem. Cell Biol. 2017, 91, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, D.; Albokhari, E.; Abba, A.; Yaradua, S. The first complete chloroplast genome sequences in Resedaceae: Genome structure and comparative analysis. Sci. Prog. 2021, 104, 003685042110599. [Google Scholar] [CrossRef] [PubMed]
- Quax, T.E.S.; Claassens, N.J.; Söll, D.; van der Oost, J. Codon Bias as a Means to Fine-Tune Gene Expression. Mol. Cell 2015, 59, 149–161. [Google Scholar] [CrossRef] [Green Version]
- Mazumdar, P.; Binti Othman, R.; Mebus, K.; Ramakrishnan, N.; Ann Harikrishna, J. Codon usage and codon pair patterns in non-grass monocot genomes. Ann. Bot. 2017, 120, 893–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, W.; Luo, C. Molecular and Functional Diversity of RNA Editing in Plant Mitochondria. Mol. Biotechnol. 2018, 60, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Shikanai, T. RNA editing in plant organelles: Machinery, physiological function and evolution. Cell. Mol. Life Sci. CMLS 2006, 63, 698–708. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.V.; Miller, J.T.; Small, I.; Nevill, P.G.; Boykin, L.M. Integration of complete chloroplast genome sequences with small amplicon datasets improves phylogenetic resolution in Acacia. Mol. Phylogenet. Evol. 2016, 96, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Jansen, R.K.; Raubeson, L.A.; Boore, J.L.; Depamphilis, C.W.; Chumley, T.W.; Haberle, R.C.; Wyman, S.K.; Alverson, A.J.; Peery, R.; Herman, S.J. Methods for obtaining and analyzing whole chloroplast genome sequences. Methods Enzymol. 2005, 395, 348–384. [Google Scholar]
- Li, Y.-C.; Korol, A.B.; Fahima, T.; Beiles, A.; Nevo, E. Microsatellites: Genomic distribution, putative functions and mutational mechanisms: A review. Mol. Ecol. 2002, 11, 2453–2465. [Google Scholar] [CrossRef] [PubMed]
- Kalia, R.K.; Rai, M.K.; Kalia, S.; Singh, R.; Dhawan, A.K. Microsatellite markers: An overview of the recent progress in plants. Euphytica 2011, 177, 309–334. [Google Scholar] [CrossRef]
- Ebert, D.; Peakall, R. Chloroplast simple sequence repeats (cpSSRs): Technical resources and recommendations for expanding cpSSR discovery and applications to a wide array of plant species. Mol. Ecol. Resour. 2009, 9, 673–690. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.H.; Zhang, J.J.; Yao, X.H.; Huang, H.W. Chloroplast microsatellite markers in Liriodendron tulipifera (Magnoliaceae) and cross-species amplification in L. chinense. Am. J. Bot. 2011, 98, e123–e126. [Google Scholar] [CrossRef]
- Liu, F.; Movahedi, A.; Yang, W.; Xu, L.; Xie, J.; Zhang, Y. The complete chloroplast genome and characteristics analysis of Callistemon rigidus R. Br. Mol. Biol. Rep. 2020, 47, 5013–5024. [Google Scholar] [CrossRef]
- Xie, D.-F.; Yu, Y.; Deng, Y.-Q.; Li, J.; Liu, H.-Y.; Zhou, S.-D.; He, X.-J. Comparative Analysis of the Chloroplast Genomes of the Chinese Endemic Genus Urophysa and Their Contribution to Chloroplast Phylogeny and Adaptive Evolution. Int. J. Mol. Sci. 2018, 19, 1847. [Google Scholar] [CrossRef] [Green Version]
- Song, W.; Chen, Z.; Shi, W.; Han, W.; Feng, Q.; Shi, C.; Engel, M.S.; Wang, S. Comparative Analysis of Complete Chloroplast Genomes of Nine Species of Litsea (Lauraceae): Hypervariable Regions, Positive Selection, and Phylogenetic Relationships. Genes 2022, 13, 1550. [Google Scholar] [CrossRef]
- Perry, A.S.; Wolfe, K.H. Nucleotide substitution rates in legume chloroplast DNA depend on the presence of the inverted repeat. J. Mol. Evol. 2002, 55, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Clegg, M.T.; Gaut, B.S.; Learn, G.H.; Morton, B.R. Rates and patterns of chloroplast DNA evolution. Proc. Natl. Acad. Sci. USA 1994, 91, 6795–6801. [Google Scholar] [CrossRef] [Green Version]
- Schutze, P.; Freitag, H.; Weising, K. An integrated molecular and morphological study of the subfamily Suaedoideae Ulbr. (Chenopodiaceae). Plant Syst. Evol. 2003, 239, 257–286. [Google Scholar] [CrossRef]
- Dong, W.; Xu, C.; Li, C.; Sun, J.; Zuo, Y.; Shi, S.; Cheng, T.; Guo, J.; Zhou, S. ycf1, the most promising plastid DNA barcode of land plants. Sci. Rep. 2015, 5, 8348. [Google Scholar] [CrossRef] [Green Version]
- Awad, M.; Fahmy, R.M.; Mosa, K.A.; Helmy, M.; El-Feky, F.A. Identification of effective DNA barcodes for Triticum plants through chloroplast genome-wide analysis. Comput. Biol. Chem. 2017, 71, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Zhu, A.; Guo, W.; Gupta, S.; Fan, W.; Mower, J.P. Evolutionary dynamics of the plastid inverted repeat: The effects of expansion, contraction, and loss on substitution rates. New Phytol. 2016, 209, 1747–1756. [Google Scholar] [CrossRef]
- Wang, R.-J.; Cheng, C.-L.; Chang, C.-C.; Wu, C.-L.; Su, T.-M.; Chaw, S.-M. Dynamics and evolution of the inverted repeat-large single copy junctions in the chloroplast genomes of monocots. BMC Evol. Biol. 2008, 8, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffares, D.C.; Tomiczek, B.; Sojo, V.; Dos Reis, M. A Beginners Guide to Estimating the Non-Synonymous to Synonymous Rate Ratio of all Protein-Coding Genes in a Genome; Springer: New York, NY, USA, 2015; pp. 65–90. [Google Scholar] [CrossRef]
- Kosiol, C.; Vinař, T.; Da Fonseca, R.R.; Hubisz, M.J.; Bustamante, C.D.; Nielsen, R.; Siepel, A. Patterns of Positive Selection in Six Mammalian Genomes. PLoS Genet. 2008, 4, e1000144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, M. The neutral theory of molecular evolution and the world view of the neutralists. Genome 1989, 31, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Bock, D.G.; Andrew, R.L.; Rieseberg, L.H. On the adaptive value of cytoplasmic genomes in plants. Mol. Ecol. 2014, 23, 4899–4911. [Google Scholar] [CrossRef]
- Drescher, A.; Ruf, S.; Calsa, T.; Carrer, H.; Bock, R. The two largest chloroplast genome-encoded open reading frames of higher plants are essential genes. Plant J. 2000, 22, 97–104. [Google Scholar] [CrossRef]
- Kuroda, H.; Maliga, P. The plastid clpP1 protease gene is essential for plant development. Nature 2003, 425, 86–89. [Google Scholar] [CrossRef]
- Sloan, D.B.; Triant, D.A.; Forrester, N.J.; Bergner, L.M.; Wu, M.; Taylor, D.R. A recurring syndrome of accelerated plastid genome evolution in the angiosperm tribe Sileneae (Caryophyllaceae). Mol. Phylogenet. Evol. 2014, 72, 82–89. [Google Scholar] [CrossRef]
- Tavares, J.C.C.; Achakkagari, S.R.; Archambault, A.; Strömvik, M.V. The plastome of Arctic Oxytropis arctobia (Fabaceae) is significantly different from that of O. splendens and other related species. Genome 2022, 65, 301–313. [Google Scholar] [CrossRef]
- Ronsted, N.; Weiblen, G.D.; Clement, W.; Zerega, N.J.; Savolainen, V. Reconstructing the phylogeny of figs (Ficus, Moraceae) to reveal the history of the fig pollination mutualism. Symbiosis 2008, 45, 45. [Google Scholar]
- Ronsted, N.; Weiblen, G.D.; Cook, J.M.; Salamin, N.; Machado, C.A.; Savolainen, V. 60 million years of co-divergence in the fig-wasp symbiosis. Proc. Biol. Sci. 2005, 272, 2593–2599. [Google Scholar] [CrossRef]
- Bruun-Lund, S.; Clement, W.L.; Kjellberg, F.; Ronsted, N. First plastid phylogenomic study reveals potential cyto-nuclear discordance in the evolutionary history of Ficus L. (Moraceae). Mol. Phylogenet. Evol. 2017, 109, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Berg, C.C.; Wiebes, J.T. African Fig Trees and Fig Wasps; Koninklijke Nederlandse Akademie van Wetenschappen, Verhandelingen Afdeling Natuurkunde, Tweede Reeks: Amsterdam, The Netherlands, 1992; Volume 89. [Google Scholar]
- Berg, C.C. Subdivision of Ficus subg. Urostigma sect. Galoglychia (Moraceae). Misc. Publ. Univ. Utrecht Herb. 1986, 1, 121–127. [Google Scholar]
- Cruaud, A.; Jabbour-Zahab, R.; Genson, G.; Ungricht, S.; Rasplus, J.-Y. Testing the Emergence of New Caledonia: Fig Wasp Mutualism as a Case Study and a Review of Evidence. PLoS ONE 2012, 7, e30941. [Google Scholar] [CrossRef] [PubMed]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | F. populifolia | Feature | F. populifolia |
|---|---|---|---|
| Plastome size (bp) | 160,610 | T (U) % | 32.44 |
| Inverted repeat (IR) region (bp) | 25,892 | C % | 18.22 |
| Large single-copy (LSC) region (bp) | 88,729 | A % | 31.70 |
| Small single-copy (SSC) region (bp) | 20,097 | G % | 17.62 |
| Total number of genes | 130 (113) | Overall GC content % | 35.84 |
| rRNA | 8 (4) | GC content in LSC % | 33.47 |
| tRNA | 37 (30) | GC content in SSC % | 28.91 |
| Protein-coding genes | 85 (79) | GC content in IR % | 42.58 |
| Gene | Location | Exon I (bp) | Entron I (bp) | Exon II (bp) | Entron II (bp) | Exon III (bp) |
|---|---|---|---|---|---|---|
| rps16 | LSC | 229 | 910 | 39 | ||
| atpF | LSC | 409 | 770 | 144 | ||
| rpoC1 | LSC | 1616 | 806 | 431 | ||
| ycf3 | LSC | 123 | 785 | 229 | 752 | 152 |
| ndhK | LSC | 60 | 0 | 740 | ||
| clpP | LSC | 70 | 903 | 291 | 701 | 263 |
| petB | LSC | 5 | 803 | 641 | ||
| petD | LSC | 7 | 736 | 474 | ||
| rpl16 | LSC | 8 | 1071 | 398 | ||
| rpI2 | IR | 390 | 676 | 433 | ||
| ndhB | IR | 755 | 676 | 594 | ||
| rps12 | IR | 25 | 537 | 231 | ||
| ndhA | SSC | 540 | 1173 | 550 | ||
| trnK-UUU | LSC | 34 | 2588 | 36 | ||
| trnG-UCC | LSC | 22 | 722 | 47 | ||
| trnL-UAA | LSC | 36 | 501 | 49 | ||
| trnV-UAC | LSC | 36 | 619 | 36 | ||
| trnI-GAU | IR | 34 | 946 | 117 | ||
| trnA-UGC | IR | 37 | 803 | 45 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alharbi, S.A.; AL-Juhani, W.S.; Albokhari, E.J. Plastome Characterization, Phylogenetic Relationships, and Regional Conservation Status of Ficus populifolia Vahl. (Moraceae), a Peripherally Isolated Plant Population in the Arabian Peninsula. Forests 2022, 13, 2063. https://doi.org/10.3390/f13122063
Alharbi SA, AL-Juhani WS, Albokhari EJ. Plastome Characterization, Phylogenetic Relationships, and Regional Conservation Status of Ficus populifolia Vahl. (Moraceae), a Peripherally Isolated Plant Population in the Arabian Peninsula. Forests. 2022; 13(12):2063. https://doi.org/10.3390/f13122063
Chicago/Turabian StyleAlharbi, Samah A., Widad S. AL-Juhani, and Enas J. Albokhari. 2022. "Plastome Characterization, Phylogenetic Relationships, and Regional Conservation Status of Ficus populifolia Vahl. (Moraceae), a Peripherally Isolated Plant Population in the Arabian Peninsula" Forests 13, no. 12: 2063. https://doi.org/10.3390/f13122063
APA StyleAlharbi, S. A., AL-Juhani, W. S., & Albokhari, E. J. (2022). Plastome Characterization, Phylogenetic Relationships, and Regional Conservation Status of Ficus populifolia Vahl. (Moraceae), a Peripherally Isolated Plant Population in the Arabian Peninsula. Forests, 13(12), 2063. https://doi.org/10.3390/f13122063