

Tree Species Drive the Diversity of Epiphytic Bryophytes in the Alpine Forest Ecosystem: A Case Study in Tibet

Abstract
:1. Introduction
2. Materials and Methods
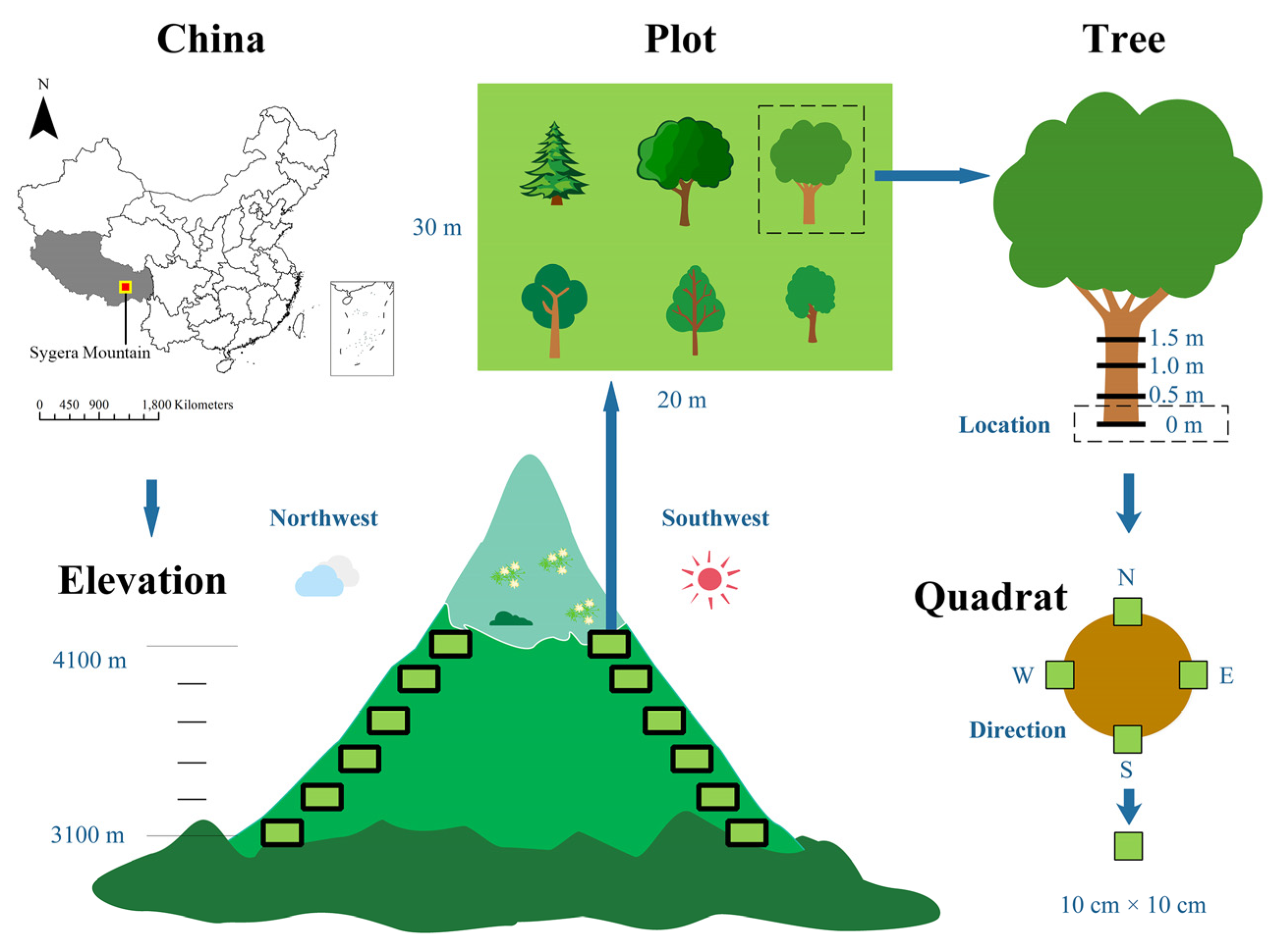
2.1. Study Area
2.2. Sampling and Identification
2.3. Environmental Variables
2.4. Data Analyses
3. Results
3.1. Diversity of Bryophytes
3.2. Relationship between Bryophyte Assemblages and Environmental Factors
4. Discussion
4.1. How Does Elevation Drive the Diversity and Assemblage Structure of Bryophytes?
4.2. How Do Tree Properties Drive the Diversity and Assemblage Structure of Bryophytes?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spence, A.R.; Tingley, M.W. The challenge of novel abiotic conditions for species undergoing climate-induced range shifts. Ecography 2020, 43, 1571–1590. [Google Scholar] [CrossRef]
- Mittelbach, G.G.; Schemske, D.W.; Cornell, H.V.; Allen, A.P.; Brown, J.M.; Bush, M.B.; Harrison, S.P.; Hurlbert, A.H.; Knowlton, N.; Lessios, H.A. Evolution and the latitudinal diversity gradient: Speciation, extinction and biogeography. Ecol. Lett. 2007, 10, 315–331. [Google Scholar] [CrossRef]
- Swenson, N.G. The role of evolutionary processes in producing biodiversity patterns, and the interrelationships between taxonomic, functional and phylogenetic biodiversity. Am. J. Bot. 2011, 98, 472–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Söderström, L.; Hagborg, A.; von Konrat, M.; Bartholomew-Began, S.; Bell, D.; Briscoe, L.; Brown, E.; Cargill, D.C.; Costa, D.P.; Crandall-Stotler, B.J. World checklist of hornworts and liverworts. PhytoKeys 2016, 59, 1–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- León Vargas, Y.; Engwald, S.; Proctor, M.C. Microclimate, light adaptation and desiccation tolerance of epiphytic bryophytes in two Venezuelan cloud forests. J. Biogeogr. 2006, 33, 901–913. [Google Scholar] [CrossRef]
- Medina, N.G.; Albertos, B.; Lara, F.; Mazimpaka, V.; Garilleti, R.; Draper, D.; Hortal, J. Species richness of epiphytic bryophytes: Drivers across scales on the edge of the Mediterranean. Ecography 2014, 37, 80–93. [Google Scholar] [CrossRef]
- de Oliveira, S.M.; ter Steege, H. Bryophyte communities in the Amazon forest are regulated by height on the host tree and site elevation. J. Ecol. 2015, 103, 441–450. [Google Scholar] [CrossRef]
- Mellado-Mansilla, D.; León, C.A.; Ortega-Solís, G.; Godoy-Güinao, J.; Moreno, R.; Díaz, I.A. Vertical patterns of epiphytic bryophyte diversity in a montane Nothofagus forest in the Chilean Andes. New Zealand J. Bot. 2017, 55, 514–529. [Google Scholar] [CrossRef]
- Ah-Peng, C.; Cardoso, A.W.; Flores, O.; West, A.; Wilding, N.; Strasberg, D.; Hedderson, T.A. The role of epiphytic bryophytes in interception, storage, and the regulated release of atmospheric moisture in a tropical montane cloud forest. J. Hydrol. 2017, 548, 665–673. [Google Scholar] [CrossRef]
- Pardow, A.; Gehrig-Downie, C.; Gradstein, R.; Lakatos, M. Functional diversity of epiphytes in two tropical lowland rainforests, French Guiana: Using bryophyte life-forms to detect areas of high biodiversity. Biodivers. Conserv. 2012, 21, 3637–3655. [Google Scholar] [CrossRef]
- Fritz, Ö.; Gustafsson, L.; Larsson, K. Does forest continuity matter in conservation?–A study of epiphytic lichens and bryophytes in beech forests of southern Sweden. Biol. Conserv. 2008, 141, 655–668. [Google Scholar] [CrossRef]
- Odland, A.; Reinhardt, S.; Pedersen, A. Differences in richness of vascular plants, mosses, and liverworts in southern Norwegian alpine vegetation. Plant Ecol. Divers. 2015, 8, 37–47. [Google Scholar] [CrossRef]
- Romanski, J.; Pharo, E.J.; Kirkpatrick, J.B. Epiphytic bryophytes and habitat variation in montane rainforest, Peru. Bryologist 2011, 114, 720–731. [Google Scholar] [CrossRef]
- Tusiime, F.M.; Byarujali, S.M.; Bates, J.W. Diversity and distribution of bryophytes in three forest types of Bwindi impenetrable National Park. Uganda. Afr. J. Ecol. 2007, 45, 79–87. [Google Scholar] [CrossRef]
- Ah-Peng, C.; Chuah-Petiot, M.; Descamps-Julien, B.; Bardat, J.; Stamenoff, P.; Strasberg, D. Bryophyte diversity and distribution along an altitudinal gradient on a lava flow in La Réunion. Divers. Distrib. 2007, 13, 654–662. [Google Scholar] [CrossRef]
- Song, L.; Ma, W.Z.; Yao, Y.L.; Liu, W.Y.; Li, S.; Chen, K.; Lu, H.Z.; Cao, M.; Sun, Z.H.; Tan, Z.H. Bole bryophyte diversity and distribution patterns along three altitudinal gradients in Yunnan, China. J. Veg. Sci. 2015, 26, 576–587. [Google Scholar] [CrossRef]
- Sun, S.Q.; Wu, Y.-H.; Wang, G.-X.; Zhou, J.; Yu, D.; Bing, H.-J.; Luo, J. Bryophyte species richness and composition along an altitudinal gradient in Gongga Mountain, China. PLoS ONE 2013, 8, e58131. [Google Scholar] [CrossRef]
- Medina, N.G.; Bowker, M.A.; Hortal, J.; Mazimpaka, V.; Lara, F. Shifts in the importance of the species pool and environmental controls of epiphytic bryophyte richness across multiple scales. Oecologia 2018, 186, 805–816. [Google Scholar] [CrossRef]
- Király, I.; Nascimbene, J.; Tinya, F.; Ódor, P. Factors influencing epiphytic bryophyte and lichen species richness at different spatial scales in managed temperate forests. Biodivers. Conserv. 2013, 22, 209–223. [Google Scholar] [CrossRef] [Green Version]
- Mežaka, A.; Brūmelis, G.; Piterāns, A. Tree and stand-scale factors affecting richness and composition of epiphytic bryophytes and lichens in deciduous woodland key habitats. Biodivers. Conserv. 2012, 21, 3221–3241. [Google Scholar] [CrossRef]
- Fritz, Ö.; Niklasson, M.; Churski, M. Tree age is a key factor for the conservation of epiphytic lichens and bryophytes in beech forests. Appl. Veg. Sci. 2009, 12, 93–106. [Google Scholar] [CrossRef]
- Patino, J.; Gómez Rodríguez, C.; Pupo Correia, A.; Sequeira, M.; Vanderpoorten, A. Trees as habitat islands: Temporal variation in alpha and beta diversity in epiphytic laurel forest bryophyte communities. J. Biogeogr. 2018, 45, 1727–1738. [Google Scholar] [CrossRef]
- Mitchell, R.J.; Hewison, R.L.; Beaton, J.; Douglass, J.R. Identifying substitute host tree species for epiphytes: The relative importance of tree size and species, bark and site characteristics. Appl. Veg. Sci. 2021, 24, e12569. [Google Scholar] [CrossRef]
- Bates, J.W.; Brown, D.H. Epiphyte differentiation between Quercus petraea and Fraxinus excelsior trees in a maritime area of South West England. Vegetatio. 1981, 48, 61–70. [Google Scholar] [CrossRef]
- Boudreault, C.; Coxson, D.S.; Vincent, E.; Bergeron, Y.; Marsh, J. Variation in epiphytic lichen and bryophyte composition and diversity along a gradient of productivity in Populus tremuloides stands of northeastern British Columbia, Canada. Ecoscience 2008, 15, 101–112. [Google Scholar] [CrossRef]
- Fritz, Ö.; Heilmann-Clausen, J. Rot holes create key microhabitats for epiphytic lichens and bryophytes on beech (Fagus sylvatica). Biol. Conserv. 2010, 143, 1008–1016. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, L.; Song, X.; Wang, Q.; Kou, J.; Jiang, Y.; Shao, X. Morphological traits of Bryum argenteum and its response to environmental variation in arid and semi-arid areas of Tibet. Ecol. Eng. 2019, 136, 101–107. [Google Scholar] [CrossRef]
- Shen, Z.; Lu, J.; Hua, M.; Tang, X.; Qu, X.; Xue, J.; Fang, J. Population structure and spatial pattern analysis of Quercus aquifolioides on Sejila Mountain, Tibet, China. J. For. Res. 2018, 29, 405–414. [Google Scholar] [CrossRef]
- Kou, J.; Wang, T. Yu, F.; Sun, Y.; Feng, C.; Shao, X. The moss genus Didymodon as an indicator of climate change on the Tibetan Plateau. Ecol. Indic. 2020, 113, 106204. [Google Scholar] [CrossRef]
- Liu, L.; Jiang, Y.; Song, X.; Tang, J.; Kou, J.; Fan, Y.; Shao, X. Temperature, not precipitation, drives the morphological traits of Didymodon rigidulus in Tibet. Ecol. Indic. 2021, 133, 108401. [Google Scholar] [CrossRef]
- Zhang, Y.; He, N.; Liu, Y. Temperature factors are a primary driver of the forest bryophyte diversity and distribution in the southeast Qinghai-Tibet Plateau. For. Ecol. Manag. 2023, 527, 120610. [Google Scholar] [CrossRef]
- Male, T.D.; Roberts, G.E. Host associations of the strangler fig Ficus watkinsiana in a subtropical Queensland rain forest. Austral Ecol. 2005, 30, 229–236. [Google Scholar] [CrossRef]
- Verma, A. Bio sorption of heavy metal Chromium (III) from synthetic waste water using Taxus baccata bark as bio adsorbent. Int. J. Chem. Sci. 2017, 5, 1758–1761. [Google Scholar]
- Kjeldahl, C. A new method for the determination of nitrogen in organic matter. Anal. Bioanal. Chem. 1883, 22, 366. [Google Scholar]
- Schielzeth, H.; Nakagawa, S. Nested by design: Model fitting and interpretation in a mixed model era. Methods Ecol. Evol. 2013, 4, 14–24. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2014, 6, 1–48. [Google Scholar]
- Dray, S.; Dufour, A. The ade4 package: Implementing the duality diagram for ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. R Package: Package ‘Lmertest’. Available online: https://github.com/runehaubo/lmerTestR (accessed on 23 October 2020).
- Lai, J.; Zou, Y.; Zhang, S.; Zhang, X.; Mao, L. Glmm.hp: An R package for computing individual effect of predictors in generalized linear mixed models. J. Plant. Ecol. 2022, 15, rtac096. [Google Scholar] [CrossRef]
- Kassambara, A. R Package: Package ‘Ggpubr’: “Ggplot2” Based Publication Ready Plots. Available online: https://github.com/tidyverse/ggplot2/issues (accessed on 4 November 2022).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; Hara, R.B.O.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. R package: Package ‘Vegan’ Community Ecology Package. Available online: https://github.com/vegandevs/vegan (accessed on 11 October 2022).
- Patiño, J.; González-Mancebo, J.M. Exploring the effect of host tree identity on epiphyte bryophyte communities in different Canarian subtropical cloud forests. Plant Ecol. 2011, 212, 433–449. [Google Scholar] [CrossRef]
- Gola, I.; Schumacker, R.; Zdeněk, S. Multivariate analysis—A tool for bryosociological studies. case study for the bryophyte epiphytic communities from the upper arieş river basin (Romania). Contrib. Bot. 2015, 50, 143–150. [Google Scholar]
- Sales, K.; Kerr, L.; Gardner, J. Factors influencing epiphytic moss and lichen distribution within Killarney National Park. Biosci. Horiz. 2016, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hugonnot, V. Remarkable bark-dwelling species in a Hautes-Alpes Juniperus thurifera L. forest with the new finding for France of Didymodon johansenii (RS Williams) HA Crum. Cryptogam. Bryol. 2014, 35, 217–222. [Google Scholar] [CrossRef]
- Song, L.; Lu, H.; Xu, X.; Li, S.; Shi, X.; Chen, X.; Wu, Y.; Huang, J.; Chen, Q.; Liu, S. Organic nitrogen uptake is a significant contributor to nitrogen economy of subtropical epiphytic bryophytes. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limpens, J.; Berendse, F. Growth reduction of Sphagnum magellanicum subjected to high nitrogen deposition: The role of amino acid nitrogen concentration. Oecologia 2003, 135, 339–345. [Google Scholar] [CrossRef]
- Pearce, I.; Woodin, S.J.; Van der Wal, R. Physiological and growth responses of the montane bryophyte Racomitrium lanuginosum to atmospheric nitrogen deposition. New Phytol. 2003, 160, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Baxter, R.; Emes, M.J.; Lee, J.A. Effects of an experimentally applied increase in ammonium on growth and amino-acid metabolism of Sphagnum cuspidatum Ehrh. ex. Hoffm. from differently polluted areas. New Phytol. 1992, 120, 265–274. [Google Scholar] [CrossRef]
- Nordin, A.; Strengbom, J.; Witzell, J.; Näsholm, T.; Ericson, L. Nitrogen deposition and the biodiversity of boreal forests: Implications for the nitrogen critical load. AMBIO A J. Hum. Environ. 2005, 34, 20–24. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Tree Type | Elevation (m) | Aspect |
|---|---|---|---|
| 1 | Salix sclerophylla | 3100 | West |
| 2 | Picea linzhiensis/Abies georgei var. smithii | 3300 | West |
| 3 | Prunus trichostoma/Abies georgei var. smithii | 3500 | West |
| 4 | Abies georgei var. smithii/Betula utilis/Salix sclerophylla | 3700 | West |
| 5 | Abies georgei var. smithii/Sorbus rehderiana | 3900 | West |
| 6 | Abies georgei var. smithii/Sorbus rehderiana | 4100 | West |
| 7 | Abies georgei var. smithii/Acer caudatum/Betula utilis | 3100 | East |
| 8 | Betula utilis/Quercus semecarpifolia | 3300 | East |
| 9 | Carpinus viminea/Quercus semecarpifolia | 3500 | East |
| 10 | Abies georgei var. smithii/Acer caudatum/Picea linzhiensis | 3700 | East |
| 11 | Abies georgei var. smithii | 3900 | East |
| 12 | Abies georgei var. smithii/ Rhododendron strigillosum var. monosematum | 4100 | East |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, J.; Song, X.; Liao, Y.; Ye, Y.; Wang, R.; Ma, H.; Shao, X. Tree Species Drive the Diversity of Epiphytic Bryophytes in the Alpine Forest Ecosystem: A Case Study in Tibet. Forests 2022, 13, 2154. https://doi.org/10.3390/f13122154
Gu J, Song X, Liao Y, Ye Y, Wang R, Ma H, Shao X. Tree Species Drive the Diversity of Epiphytic Bryophytes in the Alpine Forest Ecosystem: A Case Study in Tibet. Forests. 2022; 13(12):2154. https://doi.org/10.3390/f13122154
Chicago/Turabian StyleGu, Jiqi, Xiaotong Song, Yujia Liao, Yanhui Ye, Ruihong Wang, Heping Ma, and Xiaoming Shao. 2022. "Tree Species Drive the Diversity of Epiphytic Bryophytes in the Alpine Forest Ecosystem: A Case Study in Tibet" Forests 13, no. 12: 2154. https://doi.org/10.3390/f13122154
APA StyleGu, J., Song, X., Liao, Y., Ye, Y., Wang, R., Ma, H., & Shao, X. (2022). Tree Species Drive the Diversity of Epiphytic Bryophytes in the Alpine Forest Ecosystem: A Case Study in Tibet. Forests, 13(12), 2154. https://doi.org/10.3390/f13122154