
Genetic Diversity Revealed by Microsatellites in Genus Carya


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
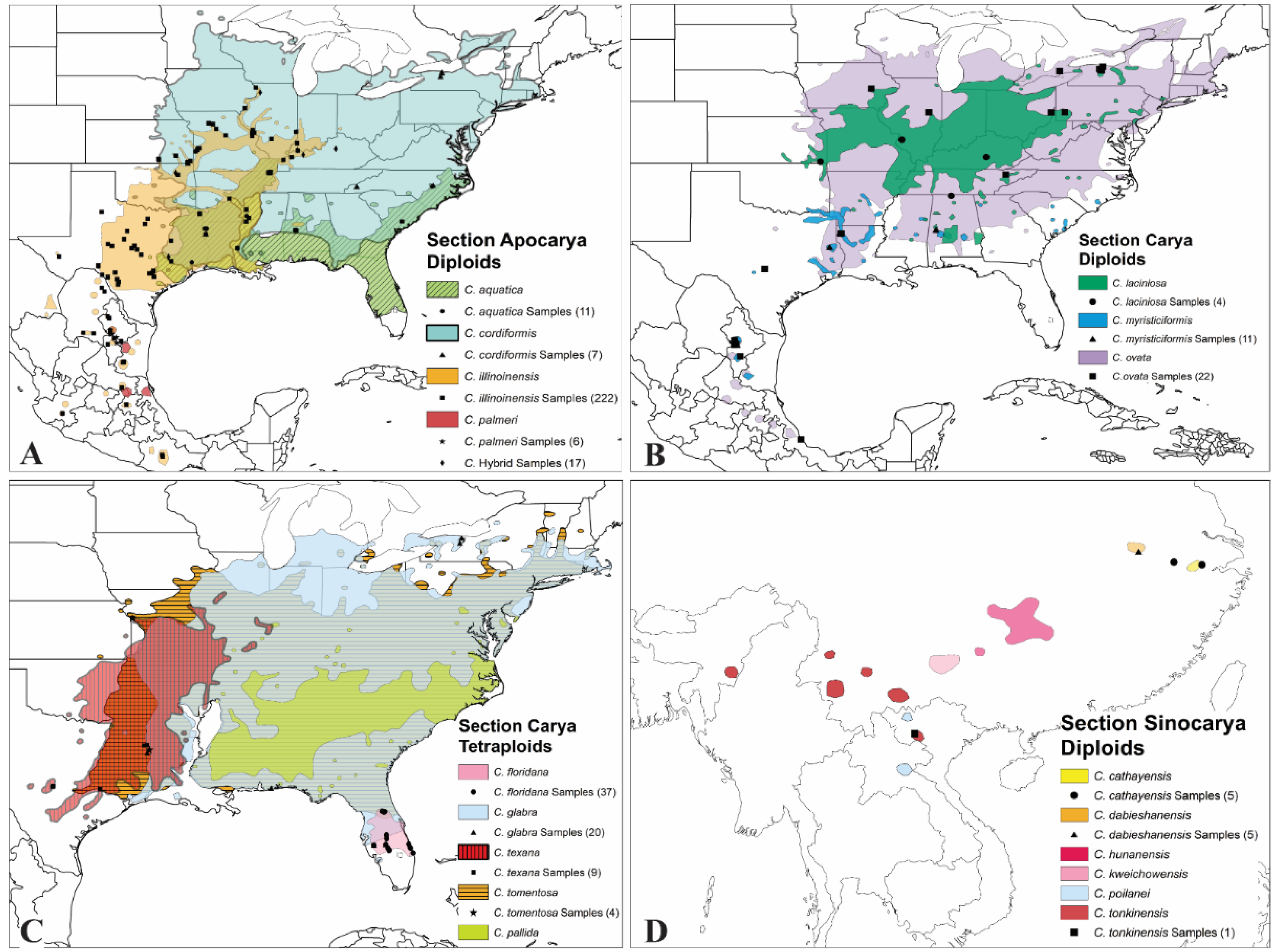
2.2. Georeferencing
2.3. DNA Extraction
2.4. PCR Amplification
2.5. Fragment Analysis
2.6. Population Structure Analysis
3. Results
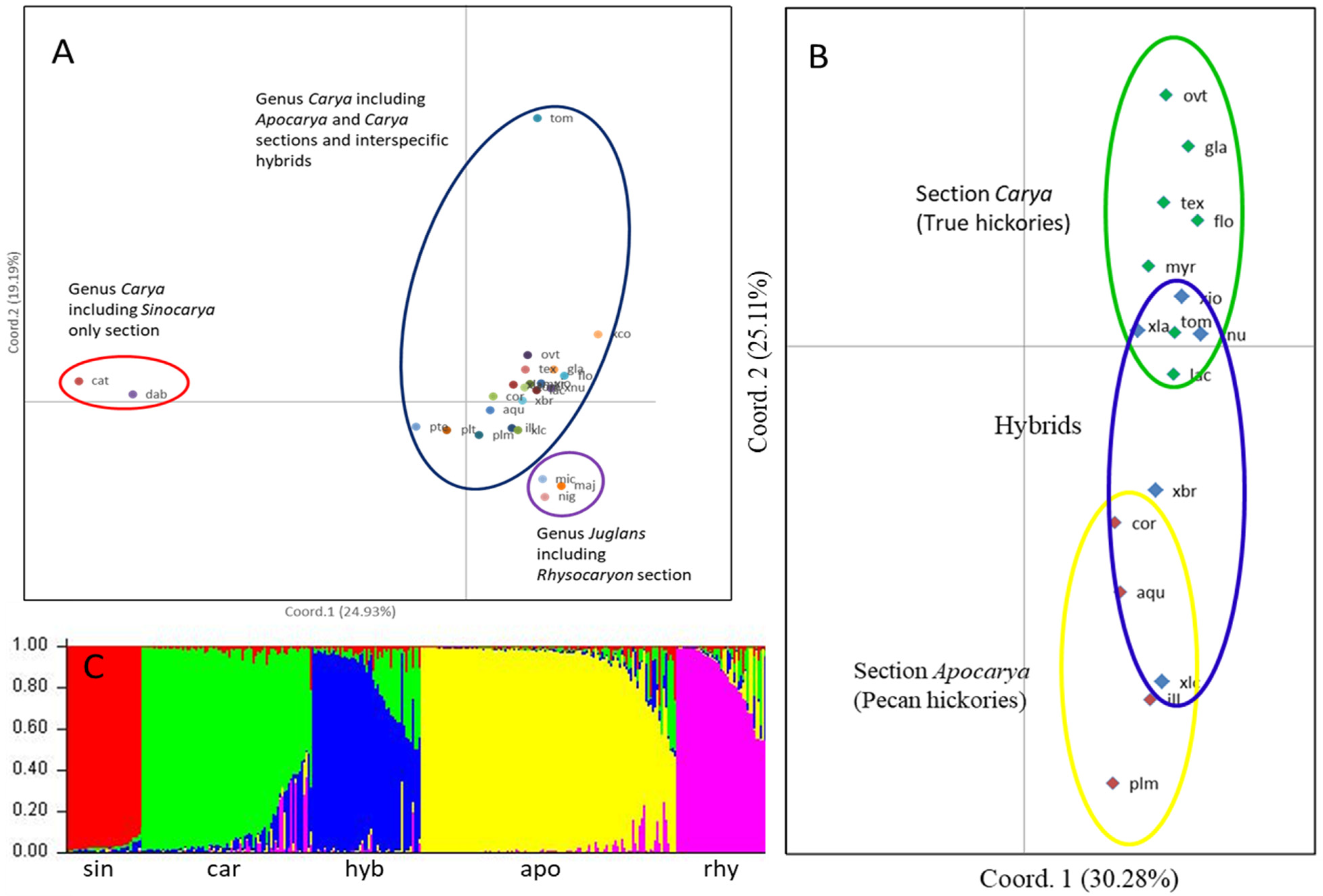
3.1. Genetic Differentiation in the Three Sections of Genus Carya
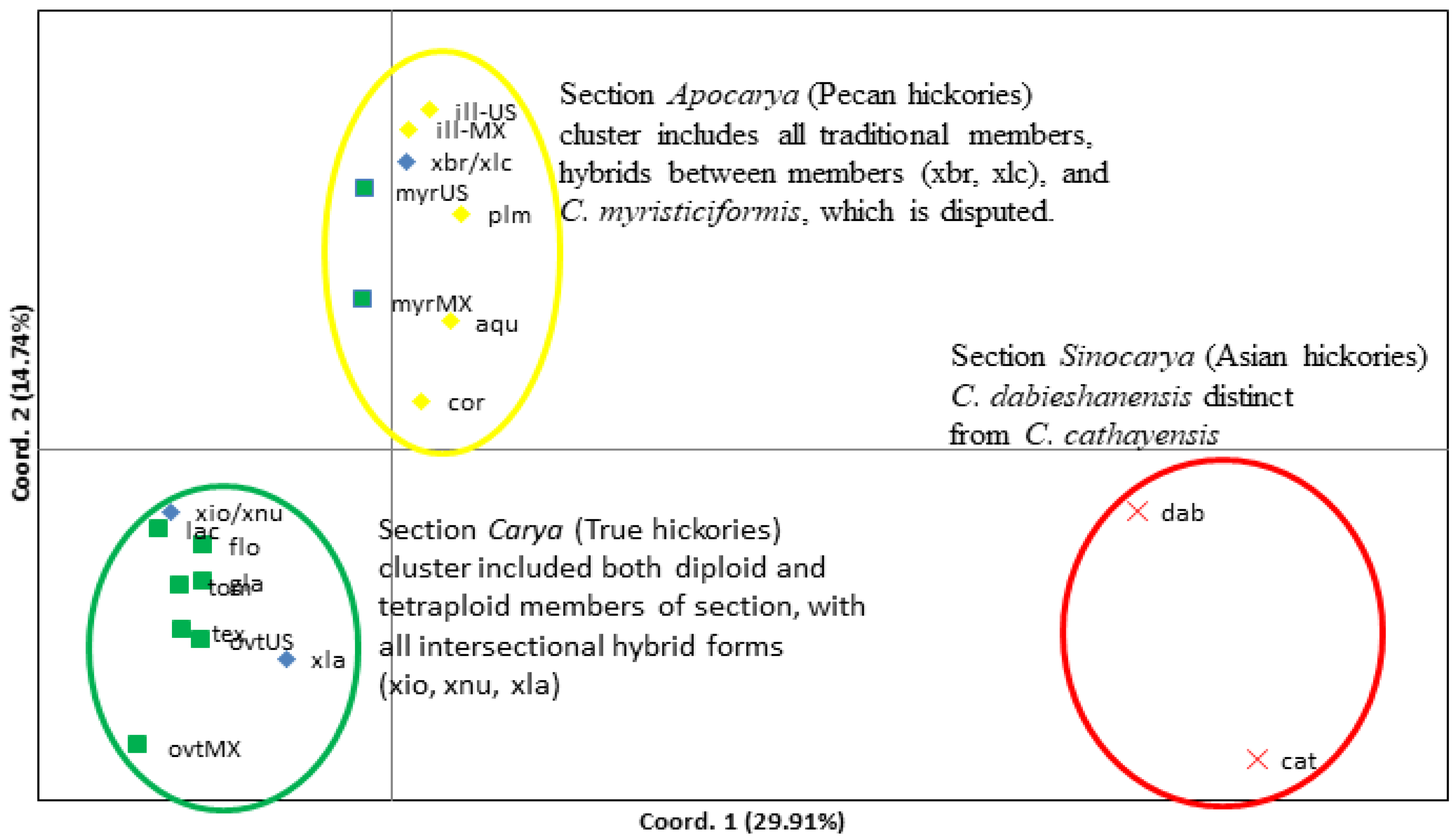
3.2. Relationships among Species within Carya Sections
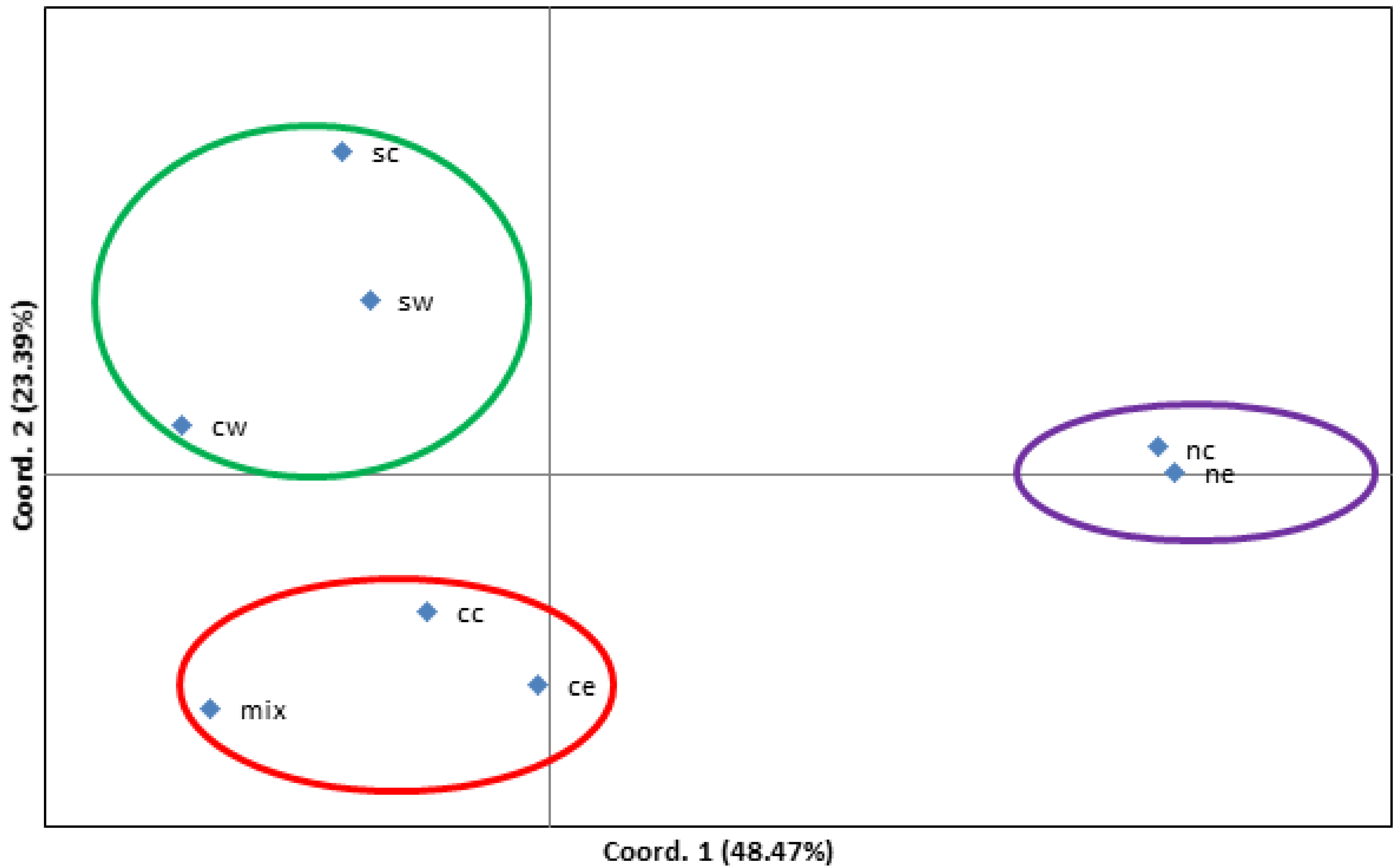
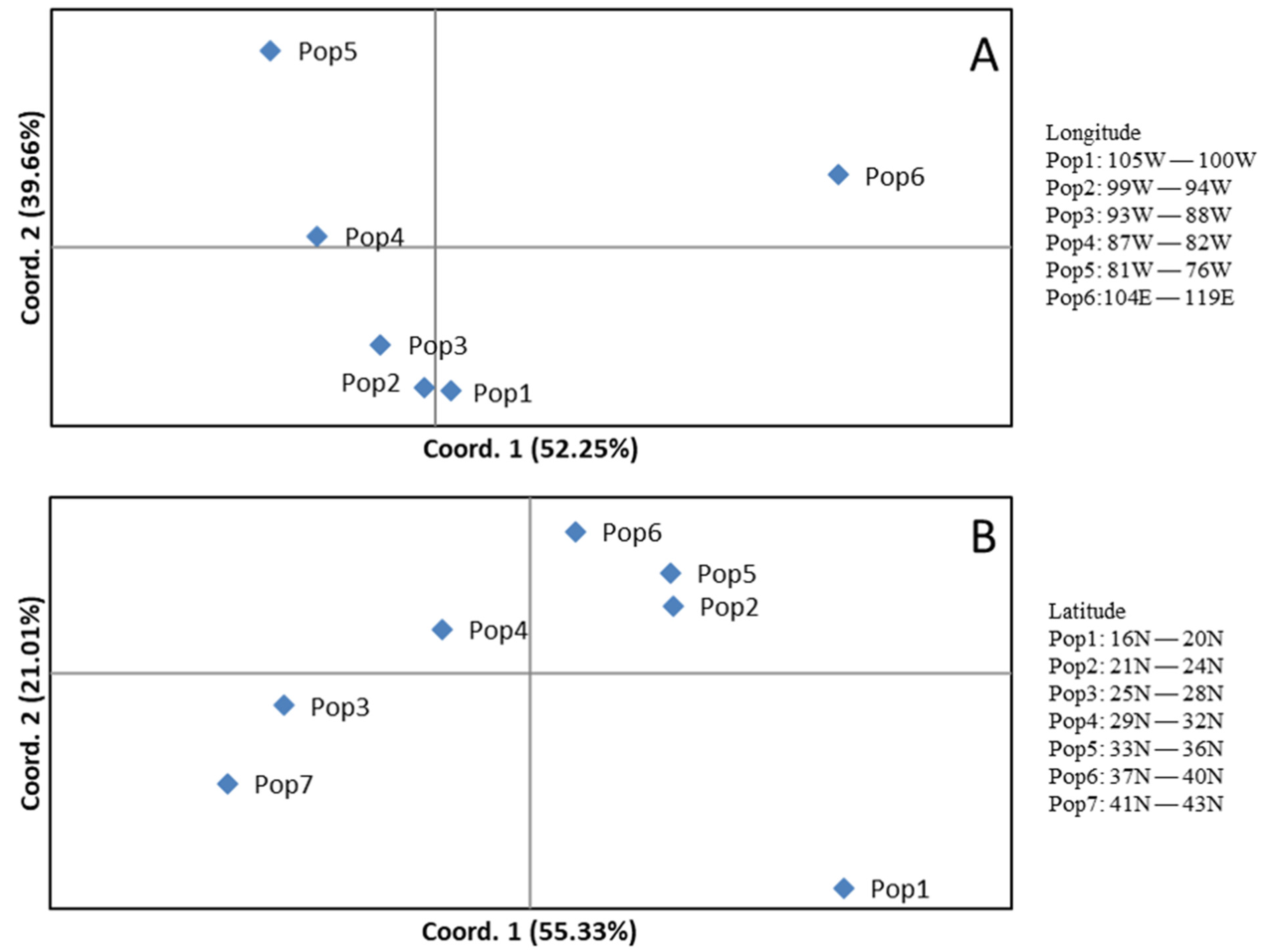
3.3. Genetic Differentiation of Carya Species Based on Geographic Origins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Manos, P.S.; Stone, D.E. Evolution, Phylogeny, and Systematics of the Juglandaceae. Ann. Mo. Bot. Gard. 2001, 88, 231. [Google Scholar] [CrossRef]
- Thompson, T.E.; Grauke, L. Pecans and Other Hickories (Carya). Acta Hortic. 1991, 290, 839–906. [Google Scholar] [CrossRef]
- Grauke, L.; Price, H.; Johnston, J. Genome Size of Pecan as Determined by Flow Cytometry. HortScience 2001, 36, 814. [Google Scholar] [CrossRef] [Green Version]
- Grauke, L.; Mendoza-Herrera, M. Population Structure in the Genus Carya. Acta Hortic. 2012, 948, 143–158. [Google Scholar] [CrossRef]
- Chang, R.-H.; Lu, A.-M. A Study of the Genus Carya Nutt. in China. J. Syst. Evol. 1979, 17, 40–44. [Google Scholar]
- Grauke, L.; Wood, B.W.; Harris, M.K. Crop Vulnerability: Carya. HortScience 2016, 51, 653–663. [Google Scholar] [CrossRef] [Green Version]
- Grauke, L.; Thompson, T.E.; Marquard, R.D. Evaluation of Pecan (Carya illinoinensis (Wangenh.) K. Koch) Germplasm Collections and Designation of a Core Subset. HortScience 1995, 30, 950–954. [Google Scholar] [CrossRef]
- Grauke, L.; Iqbal, M.J.; Reddy, A.S.; Thompson, T.E. Developing Microsatellite DNA Markers in Pecan. J. Am. Soc. Hortic. Sci. 2003, 128, 374–380. [Google Scholar] [CrossRef] [Green Version]
- Grauke, L.; Klein, R.; Grusak, M.; Klein, P. The Forest and the Trees: Applications for Molecular Markers in the Repository and Pecan Breeding Program. Acta Hortic. 2015, 1070, 109–126. [Google Scholar] [CrossRef]
- Emanuelli, F.; Lorenzi, S.; Grzeskowiak, L.; Catalano, V.; Stefanini, M.; Troggio, M.; Myles, S.; Martinez-Zapater, J.M.; Zyprian, E.; Moreira, F.M.; et al. Genetic diversity and population structure assessed by SSR and SNP markers in a large germplasm collection of grape. BMC Plant Biol. 2013, 13, 39. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Choudhury, D.R.; Singh, A.; Kumar, S.; Srinivasan, K.; Tyagi, R.K.; Singh, R. Comparison of SSR and SNP Markers in Estimation of Genetic Diversity and Population Structure of Indian Rice Varieties. PLoS ONE 2013, 8, e84136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montes-Borrego, M.; Lopes, J.R.S. Combined use of a new SNP-based assay and multilocus SSR markers to assess genetic diversity of Xylella fastidiosa subsp. pauca infecting citrus and coffee plants. Int. Microbiol. 2015, 18, 13–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chistiakov, D.A.; Hellemans, B.; Volckaert, F.A. Microsatellites and their genomic distribution, evolution, function and applications: A review with special reference to fish genetics. Aquaculture 2006, 255, 1–29. [Google Scholar] [CrossRef]
- Varshney, R.; Graner, A.; Sorrells, M.E. Genic microsatellite markers in plants: Features and applications. Trends Biotechnol. 2005, 23, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Tóth, G.; Gáspári, Z.; Jurka, J. Microsatellites in Different Eukaryotic Genomes: Survey and Analysis. Genome Res. 2000, 10, 967–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weising, K.; Gardner, R.C. A Set of Conserved PCR Primers for the Analysis of Simple Sequence Repeat Polymorphisms in Chloroplast Genomes of Dicotyledonous Angiosperms. Genome 1999, 42, 9–19. [Google Scholar] [CrossRef]
- Marquard, R.D.; Grauke, L.J.; Thompson, T.E.; Janos, R.S. Identifying Pecan Cultivars by Isozymes and Inheritance of Leucine Aminopeptidase. J. Am. Soc. Hortic. Sci. 1995, 120, 661–666. [Google Scholar] [CrossRef] [Green Version]
- Grauke, L.; Mendoza-Herrera, M.A.; Loopstra, C.; Thompson, T.E. Microsatellite Markers for Verifying Parentage of Pecans. HortScience 2006, 40, 515. [Google Scholar] [CrossRef] [Green Version]
- Rüter, B.; Hamrick, J.; Wood, B. Genetic diversity within provenance and cultivar germplasm collections versus natural populations of pecan (Carya illinoinensis). J. Hered. 1999, 90, 521–528. [Google Scholar] [CrossRef]
- Conner, P.J.; Wood, B.W. Identification of Pecan Cultivars and Their Genetic Relatedness as Determined by Randomly Am-plified Polymorphic DNA Analysis. J. Am. Soc. Hortic. Sci. 2001, 126, 474–480. [Google Scholar] [CrossRef]
- Hampe, J.; Marczinek, K.; Preuss, A.; Nürnberg, P. Marker pattern instabilities as a major cause of reproducibility problems in two-dimensional DNA fingerprinting. Electrophoresis 1996, 17, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.; Edwards, K.; Castaglione, S.; Winfield, M.; Sala, F.; Van De Wiel, C.; Bredemeijer, G.; Vosman, B.; Matthes, M.; Daly, A.; et al. Reproducibility testing of RAPD, AFLP and SSR markers in plants by a network of European laboratories. Mol. Breed. 1997, 3, 381–390. [Google Scholar] [CrossRef]
- Bian, Y.; Yang, Q.; Balint-Kurti, P.J.; Wisser, R.J.; Holland, J.B. Limits on the reproducibility of marker associations with southern leaf blight resistance in the maize nested association mapping population. BMC Genom. 2014, 15, 1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadeem, M.A.; Nawaz, M.A.; Shahid, M.Q.; Doğan, Y.; Comertpay, G.; Yıldız, M.; Hatipoğlu, R.; Ahmad, F.; Alsaleh, A.; Labhane, N.; et al. DNA molecular markers in plant breeding: Current status and recent advancements in genomic selection and genome editing. Biotechnol. Biotechnol. Equip. 2018, 32, 261–285. [Google Scholar] [CrossRef] [Green Version]
- Barchi, L.; Lanteri, S.; Portis, E.; Acquadro, A.; Valè, G.; Toppino, L.; Rotino, G.L. Identification of SNP and SSR markers in eggplant using RAD tag sequencing. BMC Genom. 2011, 12, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalapa, J.E.; Cuevas, H.; Zhu, H.; Steffan, S.; Senalik, D.; Zeldin, E.; McCown, B.; Harbut, R.; Simon, P. Using next-generation sequencing approaches to isolate simple sequence repeat (SSR) loci in the plant sciences. Am. J. Bot. 2012, 99, 193–208. [Google Scholar] [CrossRef] [Green Version]
- Bentley, N.; Grauke, L.J.; Klein, P. Genotyping by sequencing (GBS) and SNP marker analysis of diverse accessions of pecan (Carya illinoinensis). Tree Genet. Genomes 2019, 15, 8. [Google Scholar] [CrossRef]
- Cabezas, J.A.; Ibáñez, J.; Lijavetzky, D.; Vélez, D.; Bravo, G.; Rodríguez, V.; Carreño, I.; Jermakow, A.M.; Carreño, J.; Ruiz-García, L.; et al. A 48 SNP set for grapevine cultivar identification. BMC Plant Biol. 2011, 11, 153. [Google Scholar] [CrossRef] [Green Version]
- Bryan, G.J.; McNicoll, J.; Ramsay, G.; Meyer, R.C.; de Jong, W.S. Polymorphic simple sequence repeat markers in chloroplast genomes of Solanaceous plants. Theor. Appl. Genet. 1999, 99, 859–867. [Google Scholar] [CrossRef]
- Grauke, L.; Mendoza-Herrera, M.; Binzel, M. Plastid Microsatellite Markers in Carya. Acta Hortic. 2010, 859, 237–246. [Google Scholar] [CrossRef]
- Jia, X.-D.; Wang, T.; Zhai, M.; Li, Y.-R.; Guo, Z.-R. Genetic Diversity and Identification of Chinese-Grown Pecan Using ISSR and SSR Markers. Molecules 2011, 16, 10078–10092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grauke, L.J.; Mendoza-Herrera, M.A.; Miller, A.J.; Wood, B.W. Geographic patterns of genetic variation in native pecans. Tree Genet. Genomes 2011, 7, 917–932. [Google Scholar] [CrossRef]
- Woeste, K.; Burns, R.; Rhodes, O.; Michler, C. Thirty polymorphic nuclear microsatellite loci from black walnut. J. Hered. 2002, 93, 58–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amos, W.; Hoffman, J.I.; Frodsham, A.; Zhang, L.; Best, S.; Hill, A.V.S. Automated binning of microsatellite alleles: Problems and solutions. Mol. Ecol. Notes 2006, 7, 10–14. [Google Scholar] [CrossRef]
- Rinehart, T.A. AFLP analysis using GeneMapper® software and an Excel® macro that aligns and converts output to binary. BioTechniques 2004, 37, 186–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SAS Institute Inc. SAS/STAT® 9.3 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2011. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, J.; Wilson, B.; Grimwood, J.; Schmutz, J.; Grauke, L. Towards a Reference Pecan Genome Sequence. Acta Hortic. 2015, 1070, 101–108. [Google Scholar] [CrossRef]
- Wang, X.; Rhein, H.S.; Jenkins, J.; Schmutz, J.; Grimwood, J.; Grauke, L.J.; Randall, J.J. Chloroplast genome sequences of Carya illinoinensis from two distinct geographic populations. Tree Genet. Genomes 2020, 16, 48. [Google Scholar] [CrossRef]
- Moeller, D.A.; Tiffin, P. Geographic Variation in Adaptation at the Molecular Level: A Case Study of Plant Immunity Genes. Evol. Int. J. Org. Evol. 2008, 62, 3069–3081. [Google Scholar] [CrossRef] [PubMed]
- Jump, A.S.; Marchant, R.; Peñuelas, J. Environmental change and the option value of genetic diversity. Trends Plant Sci. 2009, 14, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Lovell, J.T.; Bentley, N.B.; Bhattarai, G.; Jenkins, J.W.; Sreedasyam, A.; Alarcon, Y.; Bock, C.; Boston, L.B.; Carlson, J.; Cervantes, K.; et al. Four chromosome scale genomes and a pan-genome annotation to accelerate pecan tree breeding. Nat. Commun. 2021, 12, 4125. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Section/Species | Common Name | Genus | Ploidy | Number | Abbrev. | Origin/Collection Site * |
|---|---|---|---|---|---|---|
| Section Apocarya | Pecan Hickories | 275 | apo | |||
| C. aquatica (F. Michx.) Nutt. | Water hickory | Carya | 2n = 2x = 32 | 11 | aqu | Louisiana, Mississippi, and Texas |
| C. cordiformis (Wangenh.) K. Koch | Bitternut hickory | Carya | 2n = 2x = 32 | 7 | cor | Kansas, Louisiana, New York, and North Carolina |
| C. illinoinensis (Wangenh.) K. Koch | Pecan | Carya | 2n = 2x = 32 | 251 | ill | 185 from across US (ill-US) and 66 from Mexico (ill-MX) |
| C. palmeri (Manning) | Mexican hickory | Carya | 2n = 2x = 32 | 6 | plm | Mexico |
| Section Carya | True Hickories | 108 | car | |||
| C. floridana Sarg. | Scrub hickory | Carya | 2n = 4x = 64 | 37 | flo | Florida |
| C. glabra (Mill.) Sweet | Pignut hickory | Carya | 2n = 4x = 64 | 20 | gla | Florida, Louisiana, and New York |
| C. laciniosa (F. Michx.) Loudon | Shellbark hickory | Carya | 2n = 2x = 32 | 5 | lac | Kansas, Ohio, and West Virginia |
| C. myristiciformis (F. Michx.) Nutt. | Nutmeg hickory | Carya | 2n = 2x = 32 | 11 | myr | 5 from Alabama, Louisiana, and Texas (myr-US) and 6 from Mexico (myr-MX) |
| C. ovata (Mill.) K. Koch | Shagbark hickory | Carya | 2n = 2x = 32 | 22 | ovt | 15 from Alabama, Louisiana, and Texas (ovt-US) and 7 from Mexico (ovt-MX) |
| C. texana (Buckley) | Black hickory | Carya | 2n = 4x = 64 | 9 | tex | Louisiana and Texas |
| C. tomentosa (Poir.) Nutt. | Mockernut | Carya | 2n = 4x = 64 | 4 | tom | Louisiana, Missouri, Ohio, and South Carolina |
| Section Sinocarya | Asian Hickories | Carya | 11 | sin | ||
| C. cathayensis Sarg. | Chinese hickory | Carya | 2n = 2x = 32 | 5 | cat | Zhejiang Province, PRC |
| C. dabieshanensis Liu & Li | Dabie Mountain hickory | Carya | 2n = 2x = 32 | 5 | dab | Anhui Province, PRC |
| C. tonkinensis LeComte | Vietnam hickory | Carya | 2n = 2x = 32 | 1 | ton | Vietnam |
| Hybrids | 17 | hyb | ||||
| C. ovata × C. illinoinensis | Carya | 2n = 2x = 32 | 3 | xio | Kentucky, Texas, and Mexico | |
| C. ovata × mex × mysristiformis? | Carya | 2n = 2x = 32 | 2 | xom | ||
| C. × brownii (C. illinoinensis x C. cordiformis) | Carya | 2n = 2x = 32 | 4 | xbr | Illinois, Oklahoma, Texas | |
| C. × laneyi (C. cordiformis × C.ovata) | Carya | 2n = 2x = 32 | 3 | xla | Iowa, New York Kentucky, Arkansas | |
| C. × lecontei (C. illinoinensis × C. aquatica) | Carya | 2n = 2x = 32 | 3 | xlc | Louisiana, Mississippi, Texas, | |
| C. × nussbaumeri (C. illinoinensis × C. laciniosa) | Carya | 2n = 2x = 32 | 2 | xnu | Georgia, Indiana, Iowa, Missouri, Mississippi, | |
| Rhysocaryon | Black walnuts | 7 | rhy | |||
| J. major | Arizona black walnut | Juglans | 2n = 2x = 32 | 2 | maj | Texas |
| J. microcarpa | Texas black walnut | Juglans | 2n = 2x = 32 | 3 | mic | Texas |
| J. nigra | Eastern black walnut | Juglans | 2n= 2x = 32 | 2 | nig | Texas |
| Population | N | f | p | q | Na | Ne | I | He | uHe | Polymorphic Loci (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| aqu | 11 | 0.687 ± 0.104 | 0.524 ± 0.080 | 0.967 ± 0.029 | 1.951 ± 0.065 | 1.172 ± 0.103 | 0.339 ± 0.053 | 0.576 ± 0.092 | 0.607 ± 0.096 | 23.692 |
| cor | 7 | 0.308 ± 0.087 | 0.534 ± 0.099 | 0.964 ± 0.036 | 1.972 ± 0.062 | 1.077 ± 0.073 | 0.392 ± 0.058 | 0.240 ± 0.047 | 0.260 ± 0.053 | 21.538 |
| ill-MX | 66 | 0.368 ± 0.035 | 0.521 ± 0.034 | 0.962 ± 0.014 | 1.993 ± 0.0102 | 1.166 ± 0.032 | 0.390 ± 0.022 | 0.327 ± 0.028 | 0.325 ± 0.029 | 44.308 |
| ill-US | 185 | 0.397 ± 0.018 | 0.421 ± 0.017 | 0.958 ± 0.009 | 2.000 ± 0.000 | 1.213 ± 0.019 | 0.355 ± 0.016 | 0.379 ± 0.017 | 0.360 ± 0.015 | 53.231 |
| plm | 6 | 0.373 ± 0.107 | 0.618 ± 0.127 | 0.964 ± 0.037 | 1.926 ± 0.3107 | 1.081 ± 0.084 | 0.431 ± 0.062 | 0.267 ± 0.051 | 0.291 ± 0.056 | 19.385 |
| flo | 37 | 0.389 ± 0.039 | 0.398 ± 0.044 | 0.941 ± 0.018 | 1.995 ± 0.012 | 1.253 ± 0.048 | 0.370 ± 0.035 | 0.354 ± 0.034 | 0.367 ± 0.036 | 60.308 |
| gla | 20 | 0.332 ± 0.045 | 0.353 ± 0.048 | 0.935 ± 0.026 | 1.994 ± 0.017 | 1.211 ± 0.056 | 0.314 ± 0.037 | 0.458 ± 0.062 | 0.487 ± 0.064 | 53.846 |
| lac | 5 | 0.394 ± 0.113 | 0.288 ± 0.105 | 0.964 ± 0.042 | 1.921 ± 0.121 | 1.078 ± 0.090 | 0.451 ± 0.060 | 0.285 ± 0.051 | 0.365 ± 0.039 | 17.846 |
| myrMX | 6 | 0.381 ± 0.097 | 0.546 ± 0.130 | 0.966 ± 0.035 | 1.937 ± 0.100 | 1.082 ± 0.086 | 0.457 ± 0.062 | 0.286 ± 0.049 | 0.311 ± 0.054 | 18.154 |
| myrUS | 5 | 0.439 ± 0.113 | 0.313 ± 0.105 | 0.966 ± 0.044 | 1.922 ± 0.120 | 1.074 ± 0.093 | 0.488 ± 0.062 | 0.316 ± 0.054 | 0.389 ± 0.042 | 14.462 |
| ovtMX | 7 | 0.390 ± 0.110 | 0.526 ± 0.114 | 0.971 ± 0.033 | 1.881 ± 0.122 | 1.062 ± 0.067 | 0.413 ± 0.057 | 0.256 ± 0.047 | 0.278 ± 0.052 | 16.000 |
| ovtUS | 15 | 0.427 ± 0.067 | 0.408 ± 0.058 | 0.961 ± 0.027 | 1.979 ± 0.037 | 1.134 ± 0.066 | 0.324 ± 0.044 | 0.370 ± 0.055 | 0.391 ± 0.057 | 28.308 |
| tex | 9 | 0.319 ± 0.081 | 0.398 ± 0.072 | 0.937 ± 0.042 | 1.974 ± 0.053 | 1.133 ± 0.082 | 0.397 ± 0.057 | 0.288 ± 0.036 | 0.264 ± 0.049 | 35.077 |
| tom | 4 | 0.424 ± 0.119 | 0.276 ± 0.114 | 0.958 ± 0.052 | 1.939 ± 0.119 | 1.101 ± 0.120 | 0.496 ± 0.067 | 0.315 ± 0.055 | 0.360 ± 0.063 | 19.077 |
| cat | 5 | 0.958 ± 0.063 | 0.941 ± 0.087 | 0.998 ± 0.011 | 1.083 ± 0.124 | 1.004 ± 0.025 | 0.596 ± 0.028 | 0.407 ± 0.026 | 0.452 ± 0.029 | 0.615 |
| dab | 5 | 0.883 ± 0.118 | 0.865 ± 0.135 | 0.998 ± 0.009 | 1.167 ± 0.167 | 1.005 ± 0.020 | 0.435 ± 0.044 | 0.269 ± 0.036 | 0.343 ± 0.020 | 1.231 |
| xbr/xlc | 7 | 0.321 ± 0.093 | 0.524 ± 0.107 | 0.960 ± 0.036 | 1.941 ± 0.089 | 1.085 ± 0.072 | 0.389 ± 0.054 | 0.236 ± 0.044 | 0.257 ± 0.049 | 24.615 |
| xio/xnu | 5 | 0.441 ± 0.118 | 0.320 ± 0.109 | 0.961 ± 0.048 | 1.915 ± 0.125 | 1.083 ± 0.099 | 0.481 ± 0.064 | 0.311 ± 0.055 | 0.386 ± 0.044 | 16.615 |
| xla | 3 | 0.511 ± 0.126 | 0.341 ± 0.135 | 0.965 ± 0.059 | 1.911 ± 0.164 | 1.080 ± 0.132 | 0.556 ± 0.058 | 0.373 ± 0.053 | 0.448 ± 0.064 | 12.615 |
| Population | N | f | p | q | Na | Ne | I | He | uHe | Polymorphic Loci (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| cc | 45 | 0.393 ± 0.038 | 0.356 ± 0.037 | 0.962 ± 0.016 | 1.977 ± 0.022 | 1.160 ± 0.038 | 0.358 ± 0.029 | 0.349 ± 0.030 | 0.354 ± 0.030 | 39.077 |
| ce | 27 | 0.424 ± 0.049 | 0.441 ± 0.051 | 0.960 ± 0.023 | 1.990 ± 0.020 | 1.134 ± 0.049 | 0.494 ± 0.056 | 0.368 ± 0.037 | 0.377 ± 0.038 | 29.538 |
| cw | 48 | 0.427 ± 0.037 | 0.388 ± 0.038 | 0.959 ± 0.017 | 1.982 ± 0.019 | 1.142 ± 0.037 | 0.393 ± 0.027 | 0.364 ± 0.030 | 0.375 ± 0.030 | 33.231 |
| nc | 19 | 0.425 ± 0.057 | 0.411 ± 0.058 | 0.967 ± 0.023 | 1.940 ± 0.055 | 1.120 ± 0.052 | 0.275 ± 0.040 | 0.343 ± 0.039 | 0.361 ± 0.039 | 28.923 |
| ne | 41 | 0.445 ± 0.045 | 0.351 ± 0.041 | 0.959 ± 0.019 | 1.975 ± 0.024 | 1.140 ± 0.037 | 0.413 ± 0.034 | 0.387 ± 0.038 | 0.393 ± 0.038 | 36.615 |
| sc | 41 | 0.406 ± 0.042 | 0.374 ± 0.042 | 0.965 ± 0.016 | 1.973 ± 0.025 | 1.126 ± 0.035 | 0.362 ± 0.032 | 0.352 ± 0.035 | 0.366 ± 0.037 | 33.846 |
| sw | 23 | 0.450 ± 0.049 | 0.358 ± 0.049 | 0.964 ± 0.023 | 1.990 ± 0.021 | 1.146 ± 0.053 | 0.302 ± 0.036 | 0.397 ± 0.039 | 0.407 ± 0.040 | 30.462 |
| mix | 7 | 0.356 ± 0.110 | 0.598 ± 0.101 | 0.968 ± 0.034 | 1.899 ± 0.115 | 1.068 ± 0.070 | 0.388 ± 0.061 | 0.237 ± 0.050 | 0.257 ± 0.055 | 19.077 |
| Code | ALM | DC | IL | KS | KY | mix | MO | MX1 | MX2 | MX3 | MX4 | MX5 | NLC | Se | TN | TX1 | TX2 | TX3 | TX4 | TX5 | TX6 | TXSD | N |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ALM | 24.27 | 29.10 | 24.15 | 23.95 | 23.65 | 27.23 | 23.18 | 25.14 | 22.84 | 25.31 | 29.00 | 24.33 | 26.25 | 25.26 | 23.45 | 26.19 | 25.80 | 27.28 | 26.42 | 27.77 | 23.87 | 26.68 | 10 |
| DC | 29.10 | 15.49 | 28.81 | 28.18 | 28.70 | 27.91 | 27.60 | 26.38 | 23.83 | 23.77 | 27.44 | 28.17 | 28.21 | 27.36 | 29.69 | 26.97 | 27.87 | 28.55 | 27.04 | 28.65 | 27.72 | 27.33 | 25 |
| IL | 24.15 | 28.81 | 23.77 | 23.05 | 23.03 | 27.89 | 22.41 | 25.25 | 23.82 | 26.51 | 30.01 | 24.01 | 26.60 | 25.53 | 22.40 | 26.29 | 26.61 | 27.71 | 27.38 | 28.17 | 23.61 | 27.11 | 13 |
| KS | 23.95 | 28.18 | 23.05 | 22.77 | 22.03 | 27.20 | 21.72 | 25.62 | 23.52 | 26.03 | 29.54 | 23.53 | 26.41 | 25.47 | 21.99 | 25.60 | 25.78 | 26.83 | 27.03 | 27.37 | 23.34 | 26.94 | 13 |
| KY | 23.65 | 28.70 | 23.03 | 22.03 | 20.04 | 25.84 | 21.61 | 25.52 | 22.10 | 26.88 | 29.90 | 23.48 | 26.11 | 23.97 | 21.88 | 24.64 | 25.59 | 26.08 | 26.33 | 26.72 | 23.01 | 25.88 | 8 |
| mix | 27.23 | 27.91 | 27.89 | 27.20 | 25.84 | 23.52 | 26.77 | 27.47 | 24.56 | 26.16 | 29.26 | 26.40 | 26.19 | 21.98 | 28.21 | 24.38 | 24.05 | 25.40 | 25.11 | 25.87 | 27.02 | 22.69 | 7 |
| MO | 23.18 | 27.60 | 22.41 | 21.72 | 21.61 | 26.77 | 20.00 | 25.10 | 21.82 | 25.87 | 28.91 | 22.97 | 25.62 | 24.95 | 21.44 | 25.59 | 26.18 | 26.72 | 26.43 | 27.43 | 22.75 | 26.03 | 13 |
| MX1 | 25.14 | 26.38 | 25.25 | 25.62 | 25.52 | 27.47 | 25.10 | 15.33 | 17.48 | 19.59 | 22.55 | 24.93 | 26.87 | 25.81 | 25.57 | 26.73 | 25.84 | 26.66 | 25.43 | 28.62 | 24.16 | 27.49 | 7 |
| MX2 | 22.84 | 23.83 | 23.82 | 23.52 | 22.10 | 24.56 | 21.82 | 17.48 | 11.44 | 19.13 | 22.19 | 21.44 | 24.35 | 23.54 | 23.46 | 24.81 | 24.26 | 24.53 | 24.47 | 26.50 | 22.98 | 25.93 | 9 |
| MX3 | 25.31 | 23.77 | 26.51 | 26.03 | 26.88 | 26.16 | 25.87 | 19.59 | 19.13 | 8.29 | 17.64 | 24.58 | 25.35 | 25.54 | 26.81 | 25.57 | 24.95 | 26.20 | 25.00 | 28.04 | 25.52 | 26.91 | 7 |
| MX4 | 29.00 | 27.44 | 30.01 | 29.54 | 29.90 | 29.26 | 28.91 | 22.55 | 22.19 | 17.64 | 16.06 | 28.24 | 28.00 | 28.53 | 29.81 | 28.51 | 28.33 | 29.23 | 28.10 | 30.45 | 28.06 | 29.73 | 12 |
| MX5 | 24.33 | 28.17 | 24.01 | 23.53 | 23.48 | 26.40 | 22.97 | 24.93 | 21.44 | 24.58 | 28.24 | 20.80 | 24.11 | 24.67 | 24.11 | 24.73 | 24.73 | 25.50 | 25.19 | 27.04 | 23.29 | 26.04 | 14 |
| NLC | 26.25 | 28.21 | 26.60 | 26.41 | 26.11 | 26.19 | 25.62 | 26.87 | 24.35 | 25.35 | 28.00 | 24.11 | 24.04 | 23.97 | 26.72 | 24.63 | 23.76 | 25.29 | 24.40 | 26.56 | 25.35 | 24.74 | 17 |
| se | 25.26 | 27.36 | 25.53 | 25.47 | 23.97 | 21.98 | 24.95 | 25.81 | 23.54 | 25.54 | 28.53 | 24.67 | 23.97 | 19.44 | 25.85 | 22.16 | 21.04 | 23.42 | 21.90 | 23.89 | 24.11 | 21.16 | 18 |
| TN | 23.45 | 29.69 | 22.40 | 21.99 | 21.88 | 28.21 | 21.44 | 25.57 | 23.46 | 26.81 | 29.81 | 24.11 | 26.72 | 25.85 | 20.91 | 26.82 | 26.42 | 27.07 | 27.88 | 28.05 | 23.11 | 27.30 | 12 |
| TX1 | 26.19 | 26.97 | 26.29 | 25.60 | 24.64 | 24.38 | 25.59 | 26.73 | 24.81 | 25.57 | 28.51 | 24.73 | 24.63 | 22.16 | 26.82 | 22.12 | 22.17 | 24.85 | 24.06 | 24.15 | 24.88 | 23.08 | 16 |
| TX2 | 25.80 | 27.87 | 26.61 | 25.78 | 25.59 | 24.05 | 26.18 | 25.84 | 24.26 | 24.95 | 28.33 | 24.73 | 23.76 | 21.04 | 26.42 | 22.17 | 20.75 | 22.13 | 20.93 | 23.82 | 24.07 | 22.48 | 8 |
| TX3 | 27.28 | 28.55 | 27.71 | 26.83 | 26.08 | 25.40 | 26.72 | 26.66 | 24.53 | 26.20 | 29.23 | 25.50 | 25.29 | 23.42 | 27.07 | 24.85 | 22.13 | 23.00 | 22.20 | 25.49 | 26.42 | 23.40 | 5 |
| TX4 | 26.42 | 27.04 | 27.38 | 27.03 | 26.33 | 25.11 | 26.43 | 25.43 | 24.47 | 25.00 | 28.10 | 25.19 | 24.40 | 21.90 | 27.88 | 24.06 | 20.93 | 22.20 | 19.33 | 24.46 | 25.20 | 22.56 | 10 |
| TX5 | 27.77 | 28.65 | 28.17 | 27.37 | 26.72 | 25.87 | 27.43 | 28.62 | 26.50 | 28.04 | 30.45 | 27.04 | 26.56 | 23.89 | 28.05 | 24.15 | 23.82 | 25.49 | 24.46 | 26.10 | 26.94 | 23.88 | 13 |
| TX6 | 23.87 | 27.72 | 23.61 | 23.34 | 23.01 | 27.02 | 22.75 | 24.16 | 22.98 | 25.52 | 28.06 | 23.29 | 25.35 | 24.11 | 23.11 | 24.88 | 24.07 | 26.42 | 25.20 | 26.94 | 22.06 | 25.42 | 9 |
| TXSD | 26.68 | 27.33 | 27.11 | 26.94 | 25.88 | 22.69 | 26.03 | 27.49 | 25.93 | 26.91 | 29.73 | 26.04 | 24.74 | 21.16 | 27.30 | 23.08 | 22.48 | 23.40 | 22.56 | 23.88 | 25.42 | 21.40 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Chatwin, W.; Hilton, A.; Kubenka, K. Genetic Diversity Revealed by Microsatellites in Genus Carya. Forests 2022, 13, 188. https://doi.org/10.3390/f13020188
Wang X, Chatwin W, Hilton A, Kubenka K. Genetic Diversity Revealed by Microsatellites in Genus Carya. Forests. 2022; 13(2):188. https://doi.org/10.3390/f13020188
Chicago/Turabian StyleWang, Xinwang, Warren Chatwin, Angelyn Hilton, and Keith Kubenka. 2022. "Genetic Diversity Revealed by Microsatellites in Genus Carya" Forests 13, no. 2: 188. https://doi.org/10.3390/f13020188
APA StyleWang, X., Chatwin, W., Hilton, A., & Kubenka, K. (2022). Genetic Diversity Revealed by Microsatellites in Genus Carya. Forests, 13(2), 188. https://doi.org/10.3390/f13020188