
Maxent Modeling for Identifying the Nature Reserve of Cistanche deserticola Ma under Effects of the Host (Haloxylon Bunge) Forest and Climate Changes in Xinjiang, China

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Occurrence Data
2.3. Environmental Data
2.4. Distribution Modeling
2.5. Assessing the Conservation Areas
3. Results
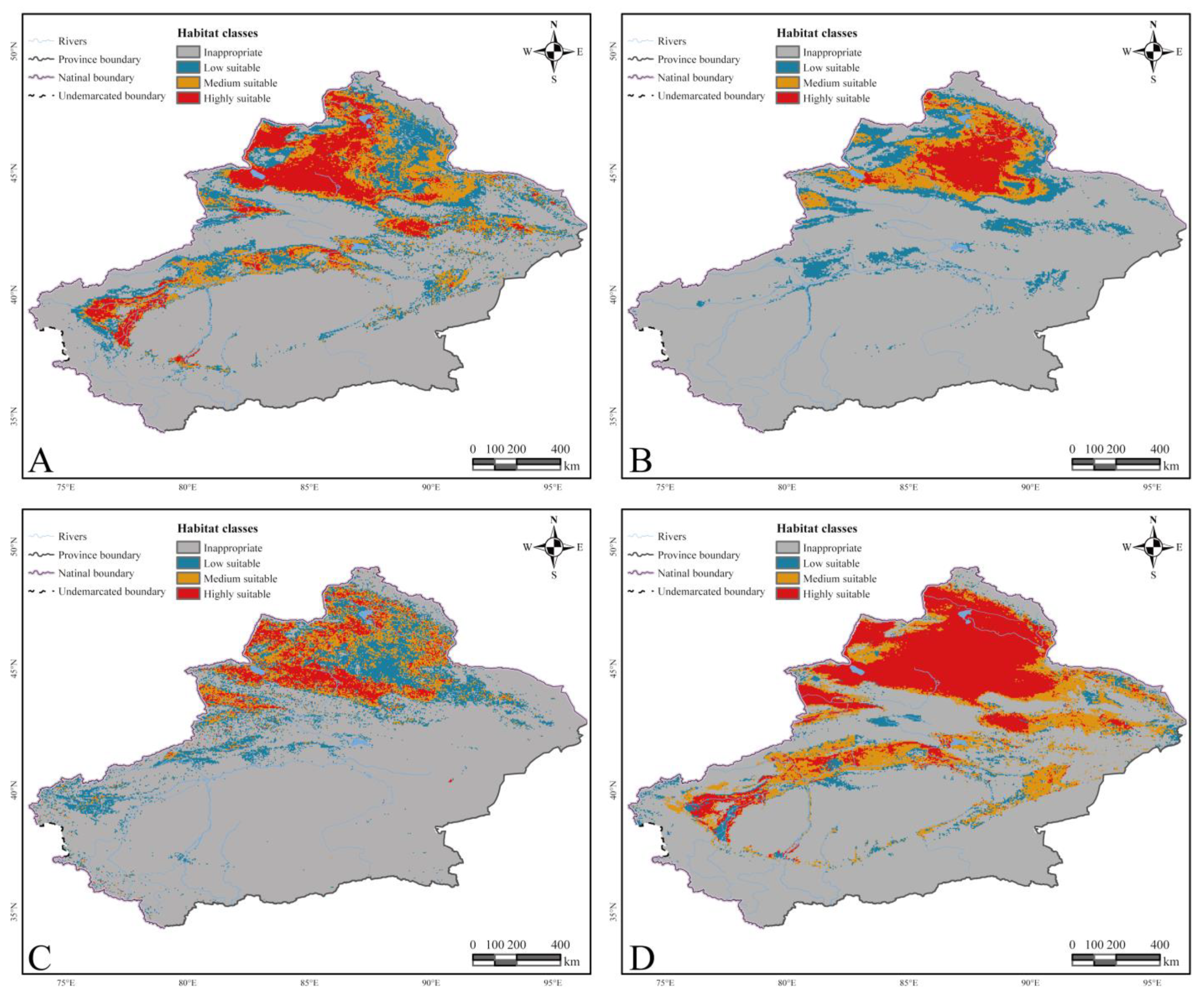
3.1. Current Distribution
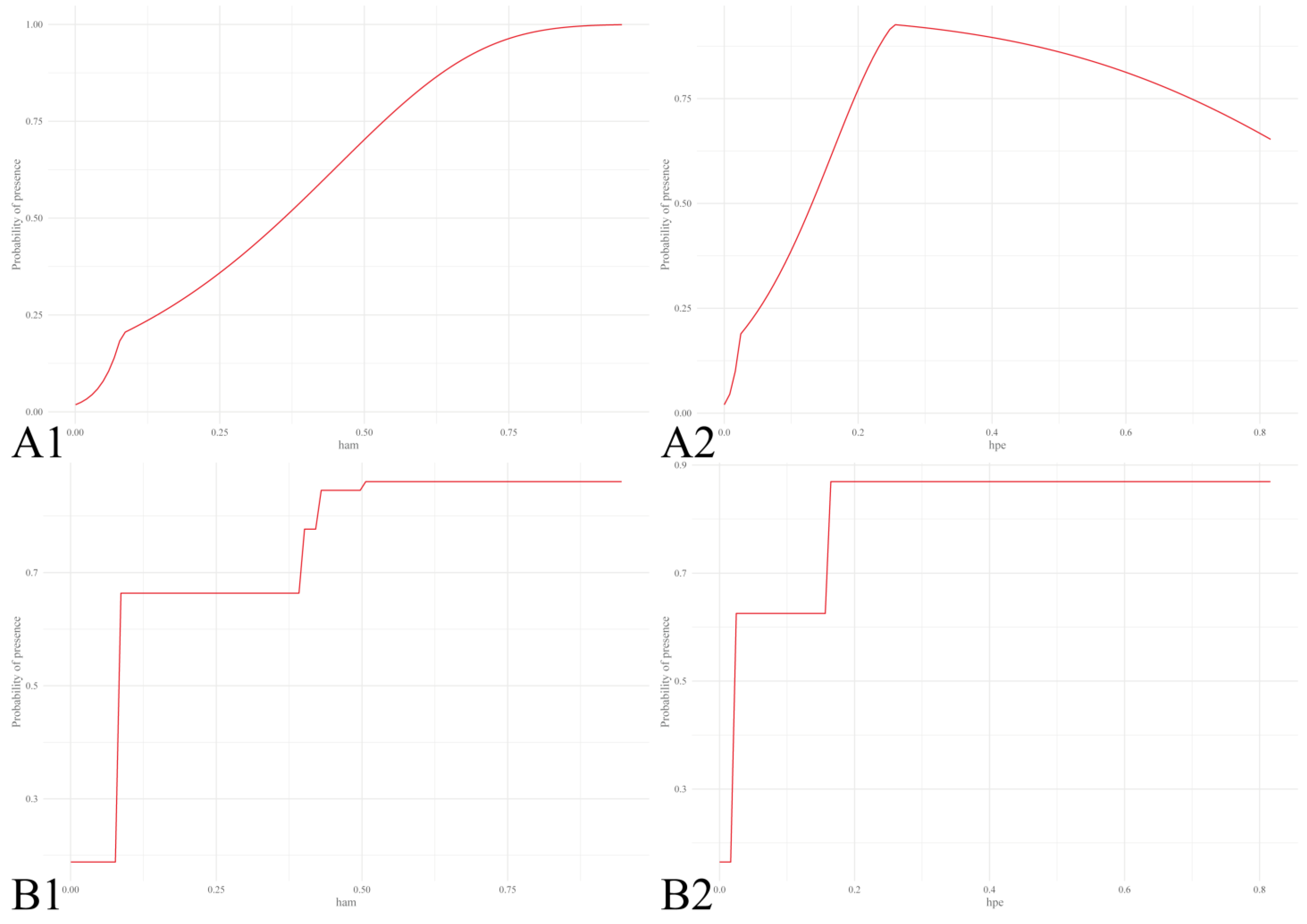
3.1.1. Abiotic and Host Factors
3.1.2. Host Factors Only
3.2. Future Distribution
3.3. The Result of Evaluation and Conservation Identifying
4. Discussion
4.1. The Better Model
4.2. The Conservation Areas of C. deserticola
5. Conclusions
- By incorporating model predictions and actual parasitic conditions, the spatial distribution of C. deserticola with the HO strategy is more realistic and accurate than AH. The HO strategy outperformed the AH strategy in terms of the effect on host distribution. It can better reflect the actual parasitic situation.
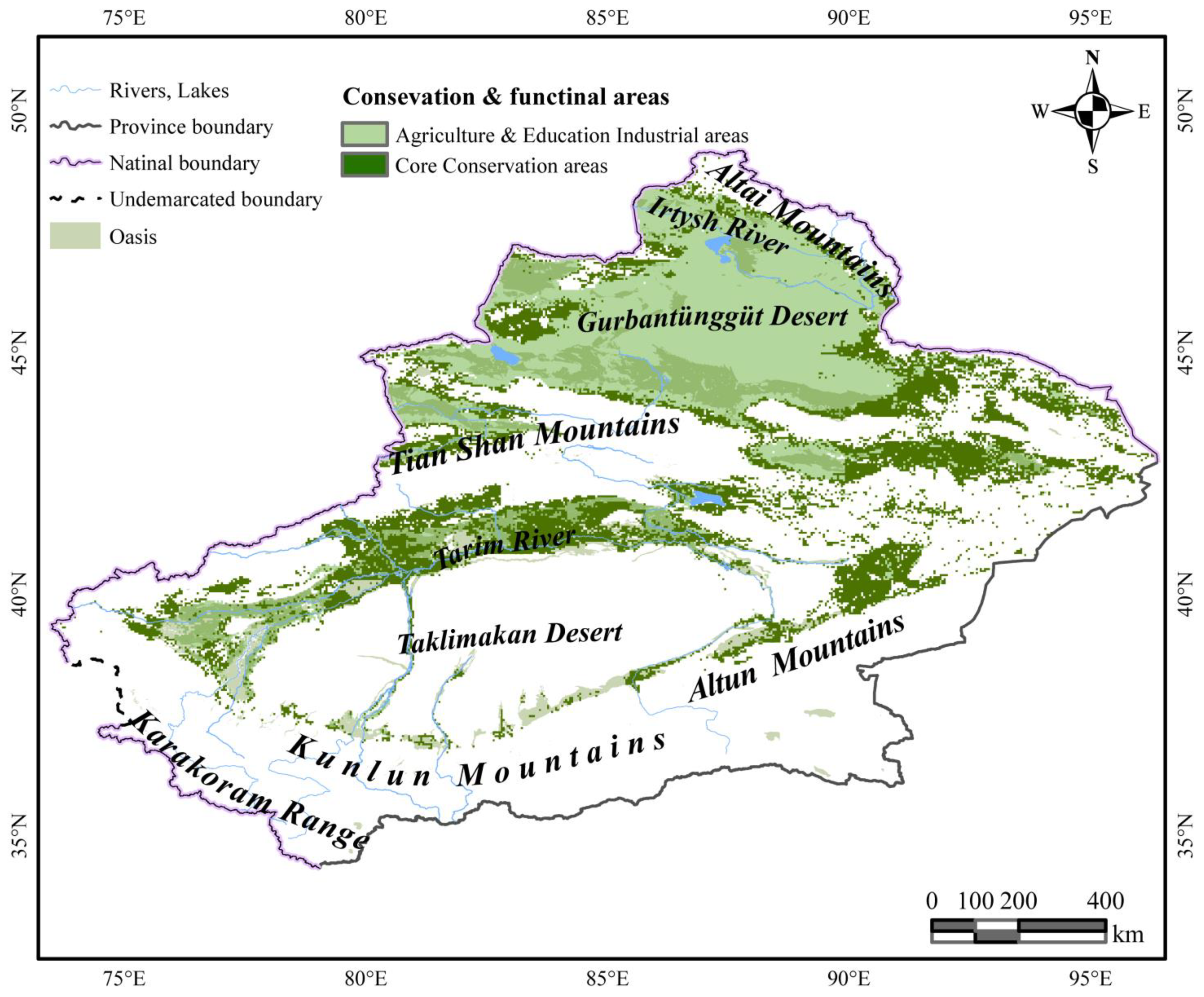
- Under various climate scenarios, the HSH is constantly increasing, reaching a maximum growth ratio of 27.3%; the LSH is more sensitive and is primarily decreasing, reaching a maximum reduction ratio of 48.2%; and the MSH has the same sensitivity as LSH, reaching a maximum reduction ratio of 26.6%. As a result, while designating the protected areas, the influence of the combination of LSH and MSH was prioritized in all climate scenarios. Additionally, given the economic development and biodiversity protection (Figure 8), the HSH is defined as agriculture and education industrial areas, aiming to promote the development of cultivated C. deserticola industry and curb the loss of biodiversity in cultivated HCCF by receiving inspiration from the natural host–parasite relationship (HCCF). The core conservation area in SSP126, SSP245, SSP370, and SSP585 is 317,315.118 km2. The HSH, which is always growing, is used as agricultural and educational industrial areas. The industrial zones cover a total area of 319,489.874 km2.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Zhou, Y.; Guo, Y.; Liu, Y.; Wu, W.; Li, Y. Targeted poverty alleviation and land policy innovation: Some practice and policy implications from China. Land Use Policy 2018, 74, 53–65. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Wu, H.; Zhu, Y.; Shi, X. Targeted poverty reduction under new structure: A perspective from mental health of older adults in rural China. China Agric. Econ. Rev. 2019, 11, 555–566. [Google Scholar] [CrossRef]
- Ge, Y.; Hu, S.; Ren, Z.; Jia, Y.; Wang, J.; Liu, M.; Zhang, D.; Zhao, W.; Luo, Y.; Fu, Y.; et al. Mapping annual land use changes in China’s poverty-stricken areas from 2013 to 2018. Remote Sens. Environ. 2019, 232, 111285. [Google Scholar] [CrossRef]
- Zhou, P.; Wang, Z.; Zhang, J.; Yang, Z.; Li, X. Study on the hydrochemical characteristics of groundwater along the Taklimakan Desert Highway. Environ. Earth Sci. 2016, 75, 1378. [Google Scholar] [CrossRef]
- Yan, W.; Qi, Y.; Yingxia, G. Current Situation and Challenges of Health Poverty Alleviation of Provinces along the Silk Road: Survey Findings in Shaanxi, Gansu and Xinjiang. Mod. China Stud. 2019, 26, 84–96. [Google Scholar]
- Yang, P.; Xia, J.; Zhang, Y.; Zhan, C.; Cai, W.; Zhang, S.; Wang, W. Quantitative study on characteristics of hydrological drought in arid area of Northwest China under changing environment. J. Hydrol. 2021, 597, 126343. [Google Scholar] [CrossRef]
- He, T.; Wang, C.; Wang, Z.; He, X.; Liu, H.; Zhang, J. Assessing the agricultural water savings–economy–ecological environment system in an arid area of northwest china using a water rights transaction model. Water 2021, 13, 1233. [Google Scholar] [CrossRef]
- Li, Y.; Lu, R.; Li, W.; Xie, Z.; Song, Y. Concentrations and size distributions of viable bioaerosols under various weather conditions in a typical semi-arid city of Northwest China. J. Aerosol. Sci. 2017, 106, 83–92. [Google Scholar] [CrossRef]
- Jia, Y.; Liu, Y.; Zhang, S. Evaluation of Agricultural Ecosystem Service Value in Arid and Semiarid Regions of Northwest China Based on the Equivalent Factor Method. Environ. Process. 2021, 8, 713–727. [Google Scholar] [CrossRef]
- Liu, S.; Huang, G. Rural Construction and Ecological Environment Optimization in Northwest China Based on Sustainability. IOP Conf. Ser. Earth Environ. Sci. 2020, 514, 32056. [Google Scholar] [CrossRef]
- Zhang, G.; Shen, D.; Ming, B.; Xie, R.; Jin, X.; Liu, C.; Hou, P.; Xue, J.; Chen, J.; Zhang, W.; et al. Using irrigation intervals to optimize water-use efficiency and maize yield in Xinjiang, northwest China. Crop J. 2019, 7, 322–334. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, X.; Xie, W. Cistanche deserticola Y. C. Ma, ‘desert Ginseng’: A review. Am. J. Chin. Med. 2012, 40, 1123–1141. [Google Scholar] [CrossRef] [PubMed]
- Announcement of National Forestry and Grassland Administration and Ministry of Agriculture and Rural Affairs (List of National Key Protected Wild Plants in China. Available online: http://www.forestry.gov.cn/main/5461/20210908/162515850572900.html (accessed on 20 November 2021).
- Jiang, Y.; Tu, P.F. Analysis of chemical constituents in Cistanche species. J. Chromatogr. A 2009, 1216, 1970–1979. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Q.B.; Kadota, S.; Tani, T.; Namba, T. Antioxidative effects of phenylethanoids from Cistanche deserticola. Biol. Pharm. Bull. 1996, 19, 1580–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, C.C.; Li, H.B.; Cheng, K.W.; Chen, F. A systematic survey of antioxidant activity of 30 Chinese medicinal plants using the ferric reducing antioxidant power assay. Food Chem. 2006, 97, 705–711. [Google Scholar] [CrossRef]
- Yue, H.; Zeng, H.; Ding, K. A review of isolation methods, structure features and bioactivities of polysaccharides from Dendrobium species. Chin. J. Nat. Med. 2020, 18, 1–27. [Google Scholar] [CrossRef]
- Dong, W.; Xu, C.; Li, D.; Jin, X.; Li, R.; Lu, Q.; Suo, Z. Comparative analysis of the complete chloroplast genome sequences in psammophytic Haloxylon species (Amaranthaceae). PeerJ 2016, 4, e2699. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Chang, H.; Liu, T.; Zhang, C. The potential geographical distribution of Haloxylon across Central Asia under climate change in the 21st century. Agric. For. Meteorol. 2019, 275, 243–254. [Google Scholar] [CrossRef]
- Song, J.; Feng, G.; Tian, C.; Zhang, F. Strategies for adaptation of Suaeda physophora, Haloxylon ammodendron and Haloxylon persicum to a saline environment during seed-germination stage. Ann. Bot. 2005, 96, 399–405. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Liu, S.; Yan, K.; Tian, L.; Li, P.; Li, X.; He, X. Effect of Drip Irrigation with Brackish Water on the Soil Chemical Properties for a Typical Desert Plant (Haloxylon Ammodendron) in the Manas River Basin. Irrig. Drain. 2020, 69, 460–471. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Mohammadi, F.; Fakhireh, A.; Bameri, A. Effects of Haloxylon spp. of Different Age Classes on Vegetation Cover and Soil Properties on an Arid Desert Steppe in Iran. Pedosphere 2019, 29, 619–631. [Google Scholar] [CrossRef]
- Khosravi Mashizi, A.; Sharafatmandrad, M. Assessing the effects of shrubs on ecosystem functions in arid sand dune ecosystems. Arid Land Res. Manag. 2020, 34, 171–187. [Google Scholar] [CrossRef]
- Di Marco, M.; Venter, O.; Possingham, H.P.; Watson, J.E.M. Changes in human footprint drive changes in species extinction risk. Nat. Commun. 2018, 9, 4621. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Qiu, H.; Guo, J.; Xu, X.; Wu, D.; Zhong, L.; Jiang, B.; Jiao, J.; Yuan, W.; Huang, Y. Modeling the potential distribution of Zelkova schneideriana under different human activity intensities and climate change patterns in China. Glob. Ecol. Conserv. 2020, 21, e00840. [Google Scholar] [CrossRef]
- Galante, P.J.; Alade, B.; Muscarella, R.; Jansa, S.A.; Goodman, S.M.; Anderson, R.P. The challenge of modeling niches and distributions for data-poor species: A comprehensive approach to model complexity. Ecography 2018, 41, 726–736. [Google Scholar] [CrossRef] [Green Version]
- Gherghel, I.; Brischoux, F.; Papeş, M. Using biotic interactions in broad-scale estimates of species’ distributions. J. Biogeogr. 2018, 45, 2216–2225. [Google Scholar] [CrossRef]
- Moreno-Amat, E.; Mateo, R.G.; Nieto-Lugilde, D.; Morueta-Holme, N.; Svenning, J.C.; García-Amorena, I. Impact of model complexity on cross-temporal transferability in Maxent species distribution models: An assessment using paleobotanical data. Ecol. Modell. 2015, 312, 308–317. [Google Scholar] [CrossRef]
- Wiese, D.; Escalante, A.A.; Murphy, H.; Henry, K.A.; Gutierrez-Velez, V.H. Integrating environmental and neighborhood factors in MaxEnt modeling to predict species distributions: A case study of Aedes albopictus in southeastern Pennsylvania. PLoS ONE 2019, 14, e0223821. [Google Scholar] [CrossRef] [Green Version]
- Mandakh, U.; Battseren, M.; Ganbat, D.; Ayanga, T.; Adiya, Z.; Borjigidai, A.; Long, C. Folk nomenclature of plants in Cistanche deserticola-associated community in South Gobi, Mongolia. Plant Divers. 2020, 42, 434–442. [Google Scholar] [CrossRef]
- Song, Y.; Zeng, K.; Jiang, Y.; Tu, P. Cistanches Herba, from an endangered species to a big brand of Chinese medicine. Med. Res. Rev. 2021, 41, 1539–1577. [Google Scholar] [CrossRef]
- Sun, X.; Pei, J.; Lin, Y.L.; Li, B.L.; Zhang, L.; Ahmad, B.; Huang, L.F. Revealing the Impact of the Environment on Cistanche salsa: From Global Ecological Regionalization to Soil Microbial Community Characteristics. J. Agric. Food Chem. 2020, 68, 8720–8731. [Google Scholar] [CrossRef] [PubMed]
- Da Cunha, H.F.; Ferreira, É.D.; Tessarolo, G.; Nabout, J.C. Host plant distributions and climate interact to affect the predicted geographic distribution of a Neotropical termite. Biotropica 2018, 50, 625–632. [Google Scholar] [CrossRef]
- Dibong, S.; Taffouo, D.V.; Boussim, J.I.; Akoa, A. Effects of host specificity in parasitic Loranthaceae for sodium and potassium distribution in coastal region of Cameroon. Int. J. Bot. 2012, 8, 159–164. [Google Scholar] [CrossRef] [Green Version]
- Sayad, E.; Boshkar, E.; Gholami, S. Different role of host and habitat features in determining spatial distribution of mistletoe infection. For. Ecol. Manage. 2017, 384, 323–330. [Google Scholar] [CrossRef]
- Hu, L.; Wang, J.; Yang, C.; Islam, F.; Bouwmeester, H.J.; Muños, S.; Zhou, W. The effect of virulence and resistance mechanisms on the interactions between parasitic plants and their hosts. Int. J. Mol. Sci. 2020, 21, 9013. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.D.; Mescher, M.C.; De Moraes, C.M. Implications of bioactive solute transfer from hosts to parasitic plants. Curr. Opin. Plant Biol. 2013, 16, 464–472. [Google Scholar] [CrossRef]
- Wang, X.M.; Yang, D.Y.; Tian, Y.Z.; Tu, P.F.; Sun, Q.S.; Li, X.B. Genetic relationship between parasitized and non-parasitized Haloxylon ammodendron in the Alxa Desert. J. Syst. Evol. 2009, 47, 255–262. [Google Scholar] [CrossRef]
- Li, N.; Lin, H.; Wang, T.; Li, Y.; Liu, Y.; Chen, X.; Hu, X. Impact of climate change on cotton growth and yields in Xinjiang, China. Field Crops Res. 2020, 247, 107590. [Google Scholar] [CrossRef]
- Li, Q.; Chen, Y.; Shen, Y.; Li, X.; Xu, J. Spatial and temporal trends of climate change in Xinjiang, China. J. Geogr. Sci. 2011, 21, 1007–1018. [Google Scholar] [CrossRef]
- Li, Y.; Sun, C. Impacts of the superimposed climate trends on droughts over 1961–2013 in Xinjiang, China. Theor. Appl. Climatol. 2017, 129, 977–994. [Google Scholar] [CrossRef]
- Heatwole, H.; Grech, A.; Marsh, H. Paleoclimatology, Paleogeography, and the Evolution and Distribution of Sea Kraits (Serpentes; Elapidae; Laticauda). Herpetol. Monogr. 2017, 31, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J. raster: Geographic Data Analysis and Modeling. 2021. Available online: https://cran.r-project.org/web/packages/raster/index.html (accessed on 24 January 2022).
- Perpiñán, O.; Hijmans, R. rasterVis. 2021. Available online: https://cran.r-project.org/web/packages/rasterVis/index.html (accessed on 24 January 2022).
- Warren, D.L.; Glor, R.E.; Turelli, M. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 2010, 33, 607–611. [Google Scholar] [CrossRef]
- Warren, D.L.; Matzke, N.J.; Cardillo, M.; Baumgartner, J.B.; Beaumont, L.J.; Turelli, M.; Glor, R.E.; Huron, N.A.; Simões, M.; Iglesias, T.L.; et al. ENMTools 1.0: An R package for comparative ecological biogeography. Ecography 2021, 44, 504–511. [Google Scholar] [CrossRef]
- Vignali, S.; Barras, A.G.; Arlettaz, R.; Braunisch, V. SDMtune: An R package to tune and evaluate species distribution models. Ecol. Evol. 2020, 10, 11488–11506. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P. Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Elith, J.H.; Graham, C.P.; Anderson, R.; Dudík, M.; Ferrier, S.; Guisan, A.J.; Hijmans, R.; Huettmann, F.R.; Leathwick, J.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Cobos, M.E.; Townsend Peterson, A.; Barve, N.; Osorio-Olvera, L. kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef] [Green Version]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Araujo, M.; Pearson, R.; Thuiller, W.; Erhard, M. Validation of species-climate impact models under climate change. Glob. Chang. Biol. 2005, 11, 1504–1513. [Google Scholar] [CrossRef] [Green Version]
- Swets, J. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzpatrick, B.M.; Turelli, M. The Geography of Mammalian Speciation: Mixed Signals from Phylogenies and Range Maps. Evolution 2006, 60, 601–615. [Google Scholar] [CrossRef]
- Lira-Noriega, A.; Peterson, A.T. Range-wide ecological niche comparisons of parasite, hosts and dispersers in a vector-borne plant parasite system. J. Biogeogr. 2014, 41, 1664–1673. [Google Scholar] [CrossRef]
- Li, Q.; Grossenbacher, D.L.; Angert, A.L. The effect of range overlap on ecological niche divergence depends on spatial scale in monkeyflowers. Evolution 2018, 72, 2100–2113. [Google Scholar] [CrossRef]
- Pastore, A.I.; Barabás, G.; Bimler, M.D.; Mayfield, M.M.; Miller, T.E. The evolution of niche overlap and competitive differences. Nat. Ecol. Evol. 2021, 5, 330–337. [Google Scholar] [CrossRef]
- Kornis, M.S.; Bunnell, D.B.; Swanson, H.K.; Bronte, C.R. Spatiotemporal patterns in trophic niche overlap among five salmonines in Lake Michigan, USA. Can. J. Fish. Aquat. Sci. 2020, 77, 1059–1075. [Google Scholar] [CrossRef]
- Chen, J.; Yang, S.; Li, H.; Zhang, B.; Lv, J. Research on geographical environment unit division based on the method of natural breaks (Jenks). In Proceedings of the ISPRS/IGU/ICA Joint Workshop on Borderlands Modelling and Understanding for Global Sustainability 2013, Beijing, China, 5–6 December 2013; Volume 40, pp. 47–50. [Google Scholar]
- Smith, J.D.; Woldemariam, M.G.; Mescher, M.C.; Jander, G.; de Moraes, C.M. Glucosinolates from host plants influence growth of the parasitic plant Cuscuta gronovii and its susceptibilityto aphid feeding. Plant Physiol. 2016, 172, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Rowntree, J.K.; Craig, H. The contrasting roles of host species diversity and parasite population genetic diversity in the infection dynamics of a keystone parasitic plant. J. Ecol. 2019, 107, 23–33. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.B.; Santos, M.J. Testing the ability of species distribution models to infer variable importance. Ecography 2020, 43, 1801–1813. [Google Scholar] [CrossRef]
- Bradie, J.; Leung, B. A quantitative synthesis of the importance of variables used in MaxEnt species distribution models. J. Biogeogr. 2017, 44, 1344–1361. [Google Scholar] [CrossRef]
- Zhu, G.; Qiao, H. Effect of the Maxent model’s complexity on the prediction of species potential distributions. Biodivers. Sci. 2016, 24, 1189–1196. [Google Scholar] [CrossRef] [Green Version]
- Westwood, J.H.; Yoder, J.I.; Timko, M.P.; dePamphilis, C.W. The evolution of parasitism in plants. Trends Plant Sci. 2010, 15, 227–235. [Google Scholar] [CrossRef]
- Wisz, M.S.; Pottier, J.; Kissling, W.D.; Pellissier, L.; Lenoir, J.; Damgaard, C.F.; Dormann, C.F.; Forchhammer, M.C.; Grytnes, J.A.; Guisan, A.; et al. The role of biotic interactions in shaping distributions and realised assemblages of species: Implications for species distribution modelling. Biol. Rev. 2013, 88, 15–30. [Google Scholar] [CrossRef] [Green Version]
- Searcy, C.A.; Bradley Shaffer, H. Do ecological niche models accurately identify climatic determinants of species ranges? Am. Nat. 2016, 187, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Konowalik, K.; Nosol, A. Evaluation metrics and validation of presence-only species distribution models based on distributional maps with varying coverage. Sci. Rep. 2021, 11, 1482. [Google Scholar] [CrossRef]
- Hirzel, A.H.; Le Lay, G.; Helfer, V.; Randin, C.; Guisan, A. Evaluating the ability of habitat suitability models to predict species presences. Ecol. Modell. 2006, 199, 142–152. [Google Scholar] [CrossRef]
- Somodi, I.; Lepesi, N.; Botta-Dukát, Z. Prevalence dependence in model goodness measures with special emphasis on true skill statistics. Ecol. Evol. 2017, 7, 863–872. [Google Scholar] [CrossRef] [Green Version]
- Sofaer, H.R.; Hoeting, J.A.; Jarnevich, C.S. The area under the precision-recall curve as a performance metric for rare binary events. Methods Ecol. Evol. 2019, 10, 565–577. [Google Scholar] [CrossRef]
- Liu, C.; White, M.; Newell, G. Measuring and comparing the accuracy of species distribution models with presence–absence data. Ecography 2011, 34, 232–243. [Google Scholar] [CrossRef]
- Araújo, M.B.; Anderson, R.P.; Barbosa, A.M.; Beale, C.M.; Dormann, C.F.; Early, R.; Garcia, R.A.; Guisan, A.; Maiorano, L.; Naimi, B. Standards for distribution models in biodiversity assessments. Sci. Adv. 2019, 5, eaat4858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valavi, R.; Guillera-Arroita, G.; Lahoz-Monfort, J.J.; Elith, J. Predictive performance of presence-only species distribution models: A benchmark study with reproducible code. Ecol Monogr. 2021, 1, e01486. [Google Scholar] [CrossRef]
- Ma, M.X.; Zhang, H.; Gao, J.X.; Ju, C.; Wang, Y.; Liu, D. Different methods comparison of delineating the ecological protection red line for biodiversity conservation. Acta Ecol. Sin. 2019, 39, 6959–6965. [Google Scholar]
- Wang, X.; Jiang, J.; Wang, Y.; Luo, W.; Song, C.; Chen, J. Responses of ephemeral plant germination and growth to water and heat conditions in the southern part of Gurbantunggut Desert. Chinese Sci. Bull. 2006, 51, 110–116. [Google Scholar] [CrossRef]
- Qin, J.; Su, B.; Tao, H.; Wang, Y.; Huang, J.; Jiang, T. Projection of temperature and precipitation under SSPs-RCPs Scenarios over northwest China. Front. Earth Sci. 2021, 15, 23–37. [Google Scholar] [CrossRef]
- Yue, P.; Cui, X.; Wu, W.; Gong, Y.; Li, K.; Misselbrook, T.; Liu, X. Are annual nitrous oxide fluxes sensitive to warming and increasing precipitation in the Gurbantunggut Desert? L. Degrad. Dev. 2021, 32, 1213–1223. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Bioclimatic Variables | Information |
|---|---|---|
| Cistanche deserticola Ma | Bio07 | Temperature annual range |
| Bio19 | Precipitation of coldest quarter | |
| aspect | Aspect | |
| slope | Slope | |
| Water_b100 | Soil water content at 100 cm depth | |
| Organic_b200 | Soil organic carbon at 200 cm depth | |
| Texture_b100 | Soil texture class at 100 cm depth | |
| Texture_b200 | Soil texture class at 200 cm depth | |
| Haloxylon ammodendron (C. A. Mey.) Bunge | Bio06 | Min Temperature of coldest month |
| Bio15 | Precipitation seasonality | |
| aspect | Aspect | |
| slope | Slope | |
| elevation | Elevation | |
| Water_b100 | Soil water content at 100 cm depth | |
| Organic_b100 | Soil organic carbon at 100 cm depth | |
| Texture_b100 | Soil texture class at 100 cm depth | |
| Texture_b200 | Soil texture class at 200 cm depth | |
| H. persicum Bunge ex Boiss | Bio07 | Temperature annual range |
| Bio13 | Precipitation of wettest month | |
| Bio17 | Precipitation of driest quarter | |
| aspect | Aspect | |
| slope | Slope | |
| Sand_b200 | Soil sand content at 200 cm depth | |
| Organic_b200 | Soil organic carbon at 200 cm depth | |
| Texture_b100 | Soil texture class at 100 cm depth | |
| Texture_b200 | Soil texture class at 200 cm depth |
| Category | AUC | Evaluation |
|---|---|---|
| H. ammodendron | 0.920 | Excellent (>0.9) |
| H. persicum | 0.960 | Excellent (>0.9) |
| C. deserticola (AH) | 0.951 | Excellent (>0.9) |
| C. deserticola (HO) | 0.922 | Excellent (>0.9) |
| Category | H. ammodendron | H. persicum | AH | HO |
|---|---|---|---|---|
| H. ammodendron | 1 | |||
| H. persicum | 0.0216 | 1 | ||
| AH | 0.540 | 0.034 | 1 | |
| HO | 0.876 | 0.291 | 0.798 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, M.; Wang, L.; Li, B.; Li, S.; Fan, J.; Li, C. Maxent Modeling for Identifying the Nature Reserve of Cistanche deserticola Ma under Effects of the Host (Haloxylon Bunge) Forest and Climate Changes in Xinjiang, China. Forests 2022, 13, 189. https://doi.org/10.3390/f13020189
Shao M, Wang L, Li B, Li S, Fan J, Li C. Maxent Modeling for Identifying the Nature Reserve of Cistanche deserticola Ma under Effects of the Host (Haloxylon Bunge) Forest and Climate Changes in Xinjiang, China. Forests. 2022; 13(2):189. https://doi.org/10.3390/f13020189
Chicago/Turabian StyleShao, Minghao, Lei Wang, Bingwen Li, Shengyu Li, Jinglong Fan, and Congjuan Li. 2022. "Maxent Modeling for Identifying the Nature Reserve of Cistanche deserticola Ma under Effects of the Host (Haloxylon Bunge) Forest and Climate Changes in Xinjiang, China" Forests 13, no. 2: 189. https://doi.org/10.3390/f13020189
APA StyleShao, M., Wang, L., Li, B., Li, S., Fan, J., & Li, C. (2022). Maxent Modeling for Identifying the Nature Reserve of Cistanche deserticola Ma under Effects of the Host (Haloxylon Bunge) Forest and Climate Changes in Xinjiang, China. Forests, 13(2), 189. https://doi.org/10.3390/f13020189