
Climate Change Impact on Peruvian Biomes



Abstract
:1. Introduction
2. Materials and Methods
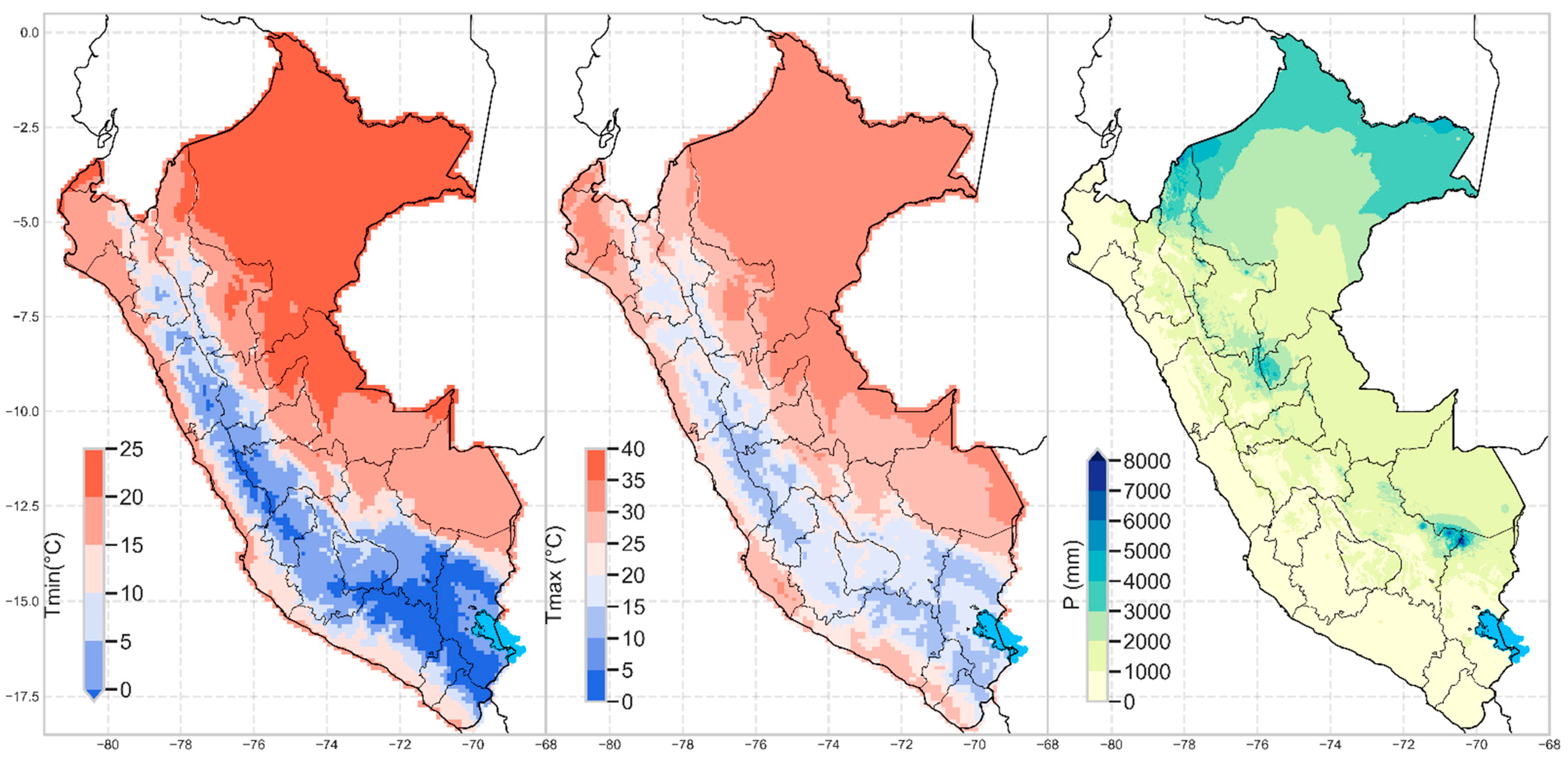
2.1. Precipitation and Temperature
2.2. Bioclimatic and Geographic Variables
2.3. Regional Models of Climate Change
2.4. Modeling of Biomes
2.5. Correction of Climate Change Scenarios
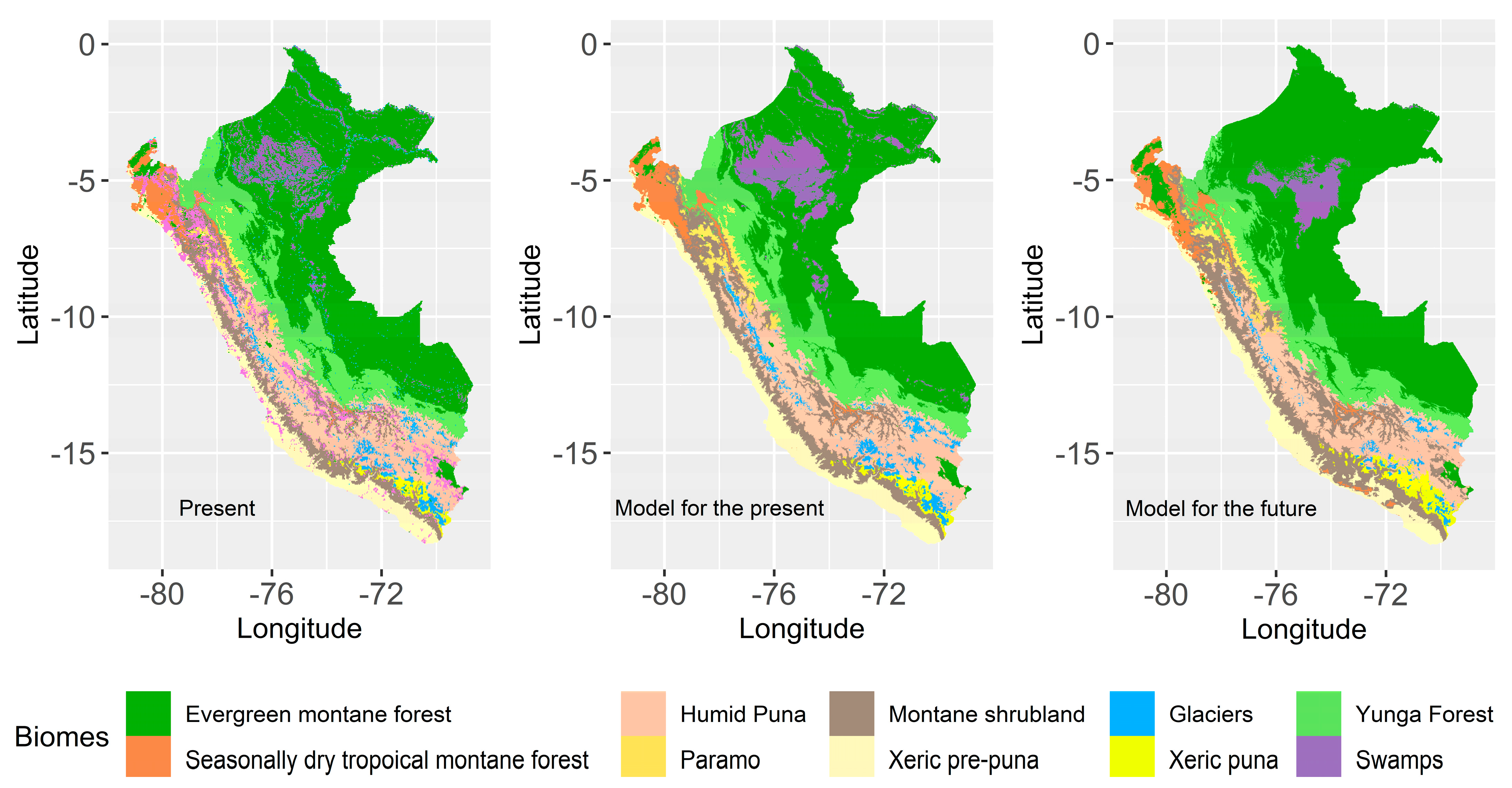
2.6. Future Biomes
2.6.1. Evaluation of Changes in Future Biomes
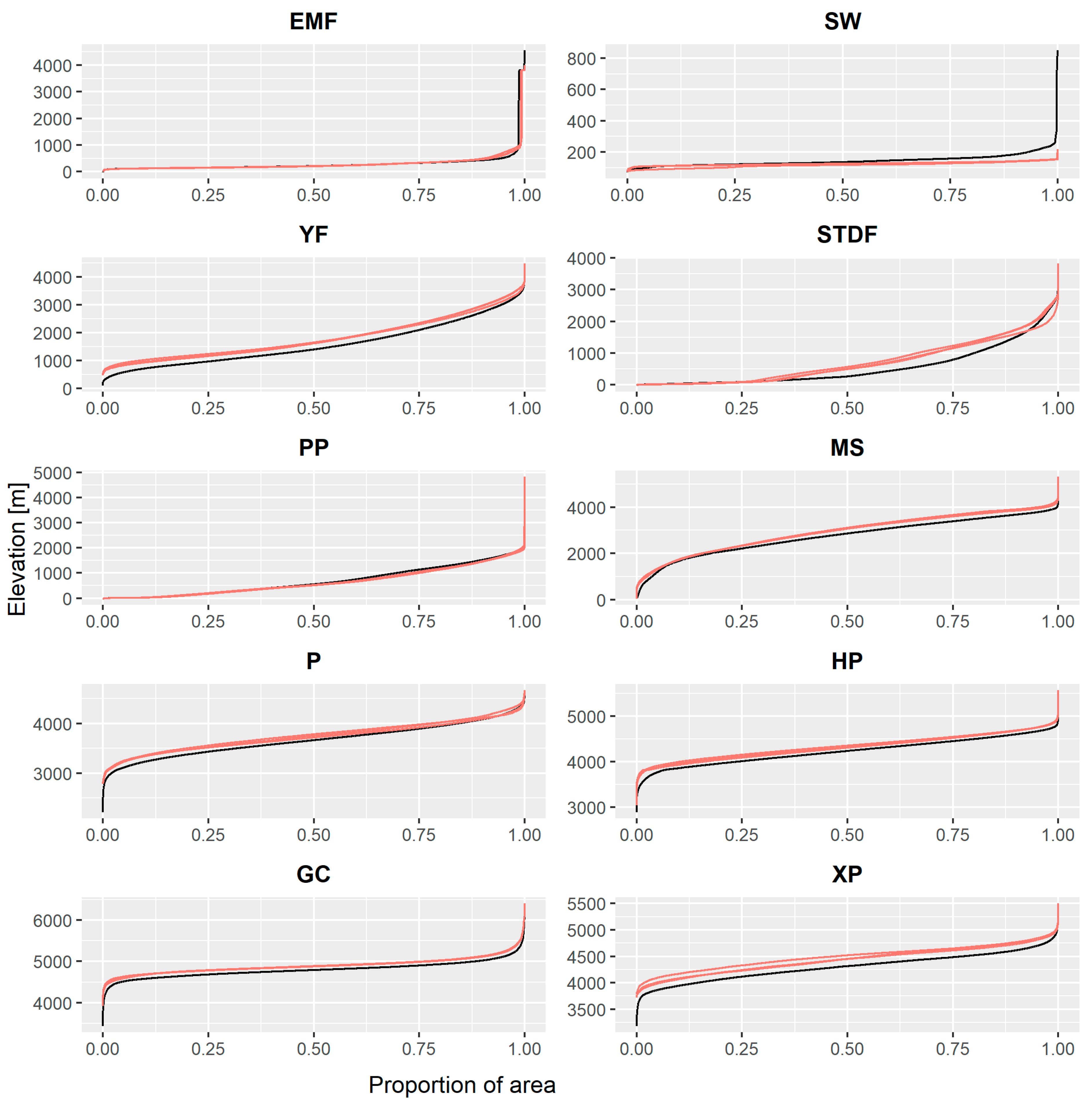
2.6.2. Vertical Displacements in Biomes
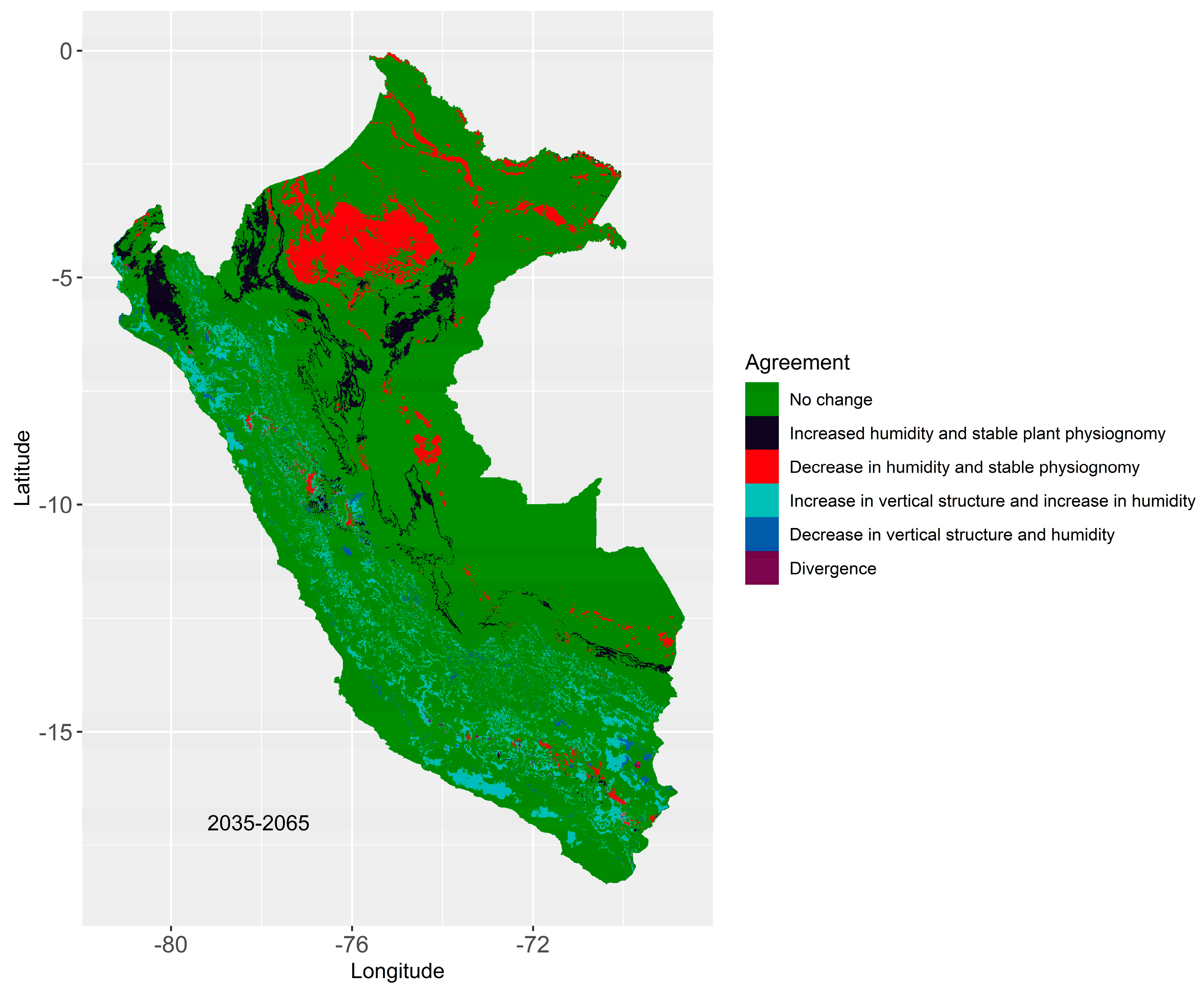
2.6.3. Regions Most Susceptible to Changes in Biomes
3. Results
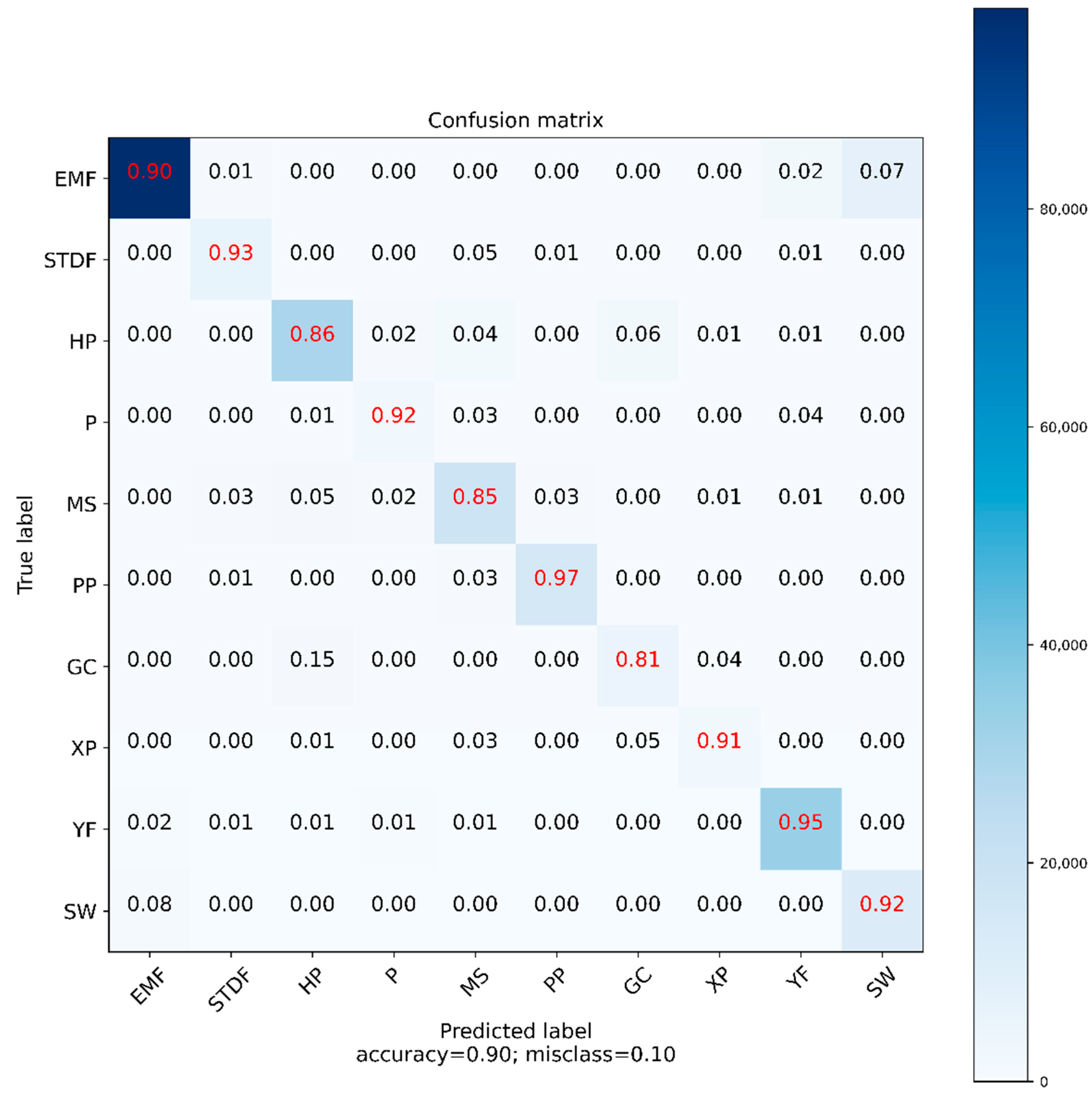
3.1. Selection of the Biome Classification Model
3.2. Vertical Displacements
3.3. Extension Changes in Biomes
3.4. Regions Most Susceptible to Changes in Their Biomes
4. Discussion
4.1. Changes in Biomes
4.2. Most Susceptible Biomes and Projected Changes
4.3. Analysis of the Modeling of Biomes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
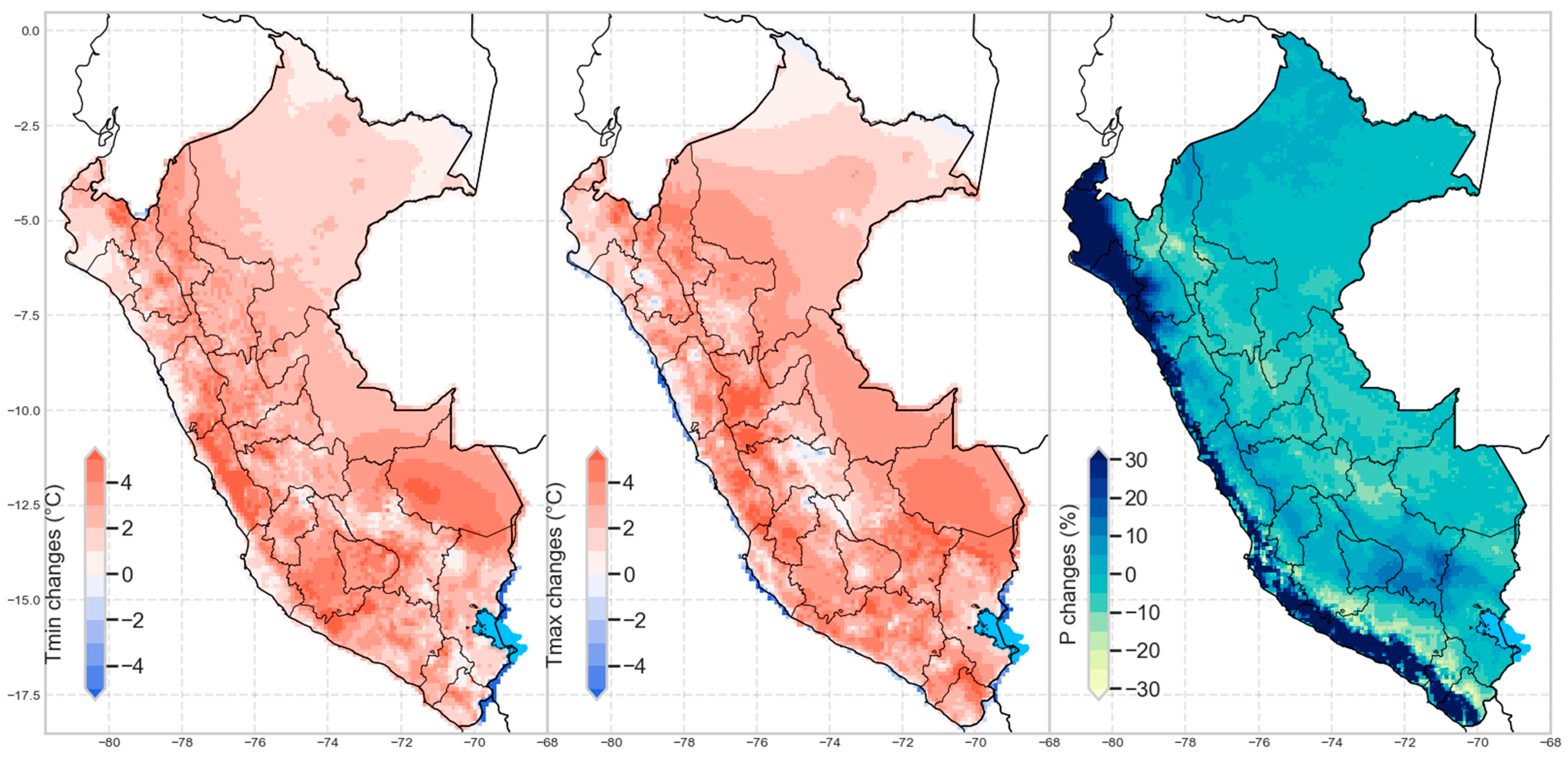
Change in Precipitation and Temperature

Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioclimatic variables |
| BIO1 = Annual Mean Temperature |
| BIO2 = Mean Diurnal Range (Mean of monthly (max temp—min temp)) |
| BIO3 = Isothermality (BIO2/BIO7) (×100) |
| BIO4 = Temperature Seasonality (standard deviation ×100) |
| BIO5 = Max Temperature of Warmest Month |
| BIO6 = Min Temperature of Coldest Month |
| BIO7 = Temperature Annual Range (BIO5-BIO6) |
| BIO8 = Mean Temperature of Wettest Quarter |
| BIO9 = Mean Temperature of Driest Quarter |
| BIO10 = Mean Temperature of Warmest Quarter |
| BIO11 = Mean Temperature of Coldest Quarter |
| BIO12 = Annual Precipitation |
| BIO13 = Precipitation of Wettest Month |
| BIO14 = Precipitation of Driest Month |
| BIO15 = Precipitation Seasonality (Coefficient of Variation) |
| BIO16 = Precipitation of Wettest Quarter |
| BIO17 = Precipitation of Driest Quarter |
| BIO18 = Precipitation of Warmest Quarter |
| BIO19 = Precipitation of Coldest Quarter |
Appendix C
| EMF | SDTF | HP | P | MS | PP | GC | XP | YF | SW | |
|---|---|---|---|---|---|---|---|---|---|---|
| EMF | 506.69 | 0.59 | 1.27 | 0.02 | 0.93 | 0.25 | 0 | 0.04 | 1.26 | 14.85 |
| SDTF | 13.44 | 31 | 0 | 0 | 1.33 | 0.65 | 0 | 0 | 0.18 | 0 |
| HP | 0.07 | 0 | 133.67 | 3.31 | 21.29 | 0 | 1.26 | 2.03 | 2.89 | 0 |
| P | 0 | 0 | 2.05 | 15.74 | 5.34 | 0 | 0.01 | 0 | 2.53 | 0 |
| MS | 0.07 | 5.91 | 1.07 | 0.66 | 116.55 | 2.05 | 0 | 0.06 | 3.31 | 0 |
| PP | 0.74 | 6.95 | 0 | 0 | 8.91 | 74.5 | 0 | 0 | 0 | 0 |
| GC | 0 | 0 | 15.24 | 0.12 | 0.03 | 0 | 16.31 | 4.57 | 0 | 0 |
| XP | 0 | 0 | 1.77 | 0 | 2.49 | 0.01 | 0.09 | 10.07 | 0 | 0 |
| YF | 30.35 | 1.46 | 0.08 | 0.16 | 1.58 | 0 | 0 | 0 | 151.61 | 0 |
| SW | 68.73 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 30.61 |
Appendix D


References
- Cramer, W.; Bondeau, A.; Woodward, F.I.; Prentice, I.C.; Betts, R.A.; Brovkin, V.; Cox, P.M.; Fisher, V.; Foley, J.A.; Friend, A.D.; et al. Global response of terrestrial ecosystem structure and function to CO2 and climate change: Results from six dynamic global vegetation models. Glob. Chang. Biol. 2001, 7, 357–373. [Google Scholar] [CrossRef] [Green Version]
- Moncrieff, G.R.; Bond, W.J.; Higgins, S.I. Revising the biome concept for understanding and predicting global change impacts. J. Biogeogr. 2016, 863–873. [Google Scholar] [CrossRef]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; George, V.N.; Underwood, E.C.; Jennifer, A.D.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the World: A New Map of Life on Earth. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Prentice, I.C.; Sykes, M.T.; Cramer, W. The possible dynamic response of northern forests to global warming. Glob. Ecol. Biogeogr. Lett. 1991, 1, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, A.; Joshi, P.K.; Ghosh, A.; Areendran, G. Assessing biome boundary shifts under climate change scenarios in India. Ecol. Indic. 2013, 34, 536–547. [Google Scholar] [CrossRef]
- Holdridge, L.R. Determination of World Plant Formations From Simple Climatic Data. Science 1947, 105, 367–368. [Google Scholar] [CrossRef]
- Walter, H.; Richards, P.W.; Wieser, J. Vegetation of the Earth in Relation to Climate and the Eco-Physiological Conditions. J. Ecol. 1973, 63, 1014–1015. [Google Scholar] [CrossRef]
- Lopes, M.S.; Veettil, B.K.; Saldanha, D.L. Buffer zone delimitation of conservation units based on map algebra and AHP technique: A study from Atlantic Forest Biome (Brazil). Biol. Conserv. 2021, 253, 108905. [Google Scholar] [CrossRef]
- Gallego-Sala, A.V.; Colin Prentice, I. Blanket peat biome endangered by climate change. Nat. Clim. Chang. 2013, 3, 152–155. [Google Scholar] [CrossRef]
- Rasquinha, D.N.; Sankaran, M. Modelling biome shifts in the Indian subcontinent under scenarios of future climate change. Curr. Sci. 2016, 111, 147–156. [Google Scholar] [CrossRef]
- Heubes, J.; Kühn, I.; König, K.; Wittig, R.; Zizka, G.; Hahn, K. Modelling biome shifts and tree cover change for 2050 in West Africa. J. Biogeogr. 2011, 38, 2248–2258. [Google Scholar] [CrossRef]
- Costa, G.C.; Hampe, A.; Pablo, M.L.; Mazzochini, G.G.; Shepard, D.B.; Werneck, F.P.; Moritz, C.; Carolina, A. Biome stability in South America over the last 30 kyr: Inferences from long-term vegetation dynamics and habitat modelling. Glob. Ecol. Biogeogr. 2018, 27, 285–297. [Google Scholar] [CrossRef]
- Giudicelli, G.C.; Turchetto, C.; Silva-Arias, G.A.; Freitas, L.B. Influence of climate changes on the potential distribution of a widespread grassland species in South America. Perspect. Plant Ecol. Evol. Syst. 2019, 41, 125496. [Google Scholar] [CrossRef]
- MINAM (Ministerio del Ambiente). Mapa Nacional de Ecosistemas del Perú—Memoria Descriptiva. 2019. Available online: https://redd.unfccc.int/files/nref_peru_final.pdf (accessed on 30 September 2021).
- Scullion, J.J.; Vogt, K.A.; Sienkiewicz, A.; Gmur, S.J.; Trujillo, C. Assessing the influence of land-cover change and conflicting land-use authorizations on ecosystem conversion on the forest frontier of Madre de Dios, Peru. Biol. Conserv. 2014, 171, 247–258. [Google Scholar] [CrossRef]
- Brêda, J.P.L.F.; de Paiva, R.C.D.; Collischon, W.; Bravo, J.M.; Siqueira, V.A.; Steinke, E.B. Climate change impacts on South American water balance from a continental-scale hydrological model driven by CMIP5 projections. Clim. Chang. 2020, 159, 503–522. [Google Scholar] [CrossRef]
- Boit, A.; Sakschewski, B.; Boysen, L.; Cano-Crespo, A.; Clement, J.; Garcia-alaniz, N.; Kok, K.; Kolb, M.; Langerwisch, F.; Rammig, A.; et al. Large-scale impact of climate change vs. land-use change on future biome shifts in Latin America. Glob. Chang. Biol. 2016, 22, 3689–3701. [Google Scholar] [CrossRef]
- Tovar, C.; Arnillas, C.A.; Cuesta, F.; Buytaert, W. Diverging Responses of Tropical Andean Biomes under Future Climate Conditions. PLoS ONE 2013, 8, e63634. [Google Scholar] [CrossRef] [Green Version]
- Bax, V.; Castro-Nunez, A.; Francesconi, W. Assessment of potential climate change impacts on montane forests in the peruvian andes: Implications for conservation prioritization. Forests 2021, 12, 375. [Google Scholar] [CrossRef]
- Ramirez-Villegas, J.; Cuesta, F.; Devenish, C.; Peralvo, M.; Jarvis, A.; Arnillas, C.A. Using species distributions models for designing conservation strategies of Tropical Andean biodiversity under climate change. J. Nat. Conserv. 2014, 22, 391–404. [Google Scholar] [CrossRef] [Green Version]
- Garreaud, R.D.; Vuille, M.; Compagnucci, R.; Marengo, J. Present-day South American climate. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 281, 180–195. [Google Scholar] [CrossRef]
- Lavado Casimiro, W.S.; Ronchail, J.; Labat, D.; Espinoza, J.C.; Guyot, J.L. Basin-scale analysis of rainfall and runoff in Peru (1969–2004): Pacific, Titicaca and Amazonas drainages. Hydrol. Sci. J. 2012, 57, 625–642. [Google Scholar] [CrossRef] [Green Version]
- Castro, A.; Davila, C.; Laura, W.; Cubas, F.; Ávalos, G.; Ocaña, C.L.; Villena, D.; Valdez, M.; Urbiola, J.; Trebejo, I.; et al. Climas del Perú. In Mapa de Clasificación Climática Nacional; 2020; Available online: https://www.senamhi.gob.pe/load/file/01404SENA-4.pdf (accessed on 30 September 2021).
- Funk, C.; Peterson, P.; Landsfeld, M.; Pedreros, D.; Verdin, J.; Shukla, S.; Husak, G.; Rowland, J.; Harrison, L.; Hoell, A.; et al. The climate hazards infrared precipitation with stations—A new environmental record for monitoring extremes. Sci. Data 2015, 2, 150066. [Google Scholar] [CrossRef] [Green Version]
- Vancutsem, C.; Ceccato, P.; Dinku, T.; Connor, S.J. Evaluation of MODIS land surface temperature data to estimate air temperature in different ecosystems over Africa. Remote Sens. Environ. 2010, 114, 449–465. [Google Scholar] [CrossRef]
- Aybar, C.; Fernández, C.; Huerta, A.; Lavado, W.; Vega, F. Construction of a high-resolution gridded rainfall dataset for Peru from 1981 to the present day. Hydrol. Sci. J. 2020, 65, 770–785. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Elith, J. Species distribution modeling with R Introduction. R CRAN Proj. 2013, 6, 71. [Google Scholar]
- Skamarock, W.C.; Klemp, J.B.; Dudhia, J.B.; Gill, D.O.; Barker, D.M.; Duda, M.G.; Huang, X.-Y.; Wang, W.; Powers, J.G. A description of the Advanced Research WRF Version 3, NCAR Technical Note TN-475+STR. Tech. Rep. 2008. [Google Scholar] [CrossRef]
- Jungclaus, J.H.; Fischer, N.; Haak, H.; Lohmann, K.; Marotzke, J.; Matei, D.; Mikolajewicz, U.; Notz, D.; Von Storch, J.S. Characteristics of the ocean simulations in the Max Planck Institute Ocean Model (MPIOM) the ocean component of the MPI-Earth system model. J. Adv. Model. Earth Syst. 2013, 5, 422–446. [Google Scholar] [CrossRef]
- Bi, D.; Dix, M.; Marsland, S.J.; O’Farrell, S.; Rashid, H.A.; Uotila, P.; Hirst, A.C.; Kowalczyk, E.; Golebiewski, M.; Sullivan, A.; et al. The ACCESS coupled model: Description, control climate and evaluation. Aust. Meteorol. Oceanogr. J. 2013, 63, 41–64. [Google Scholar] [CrossRef]
- Collins, W.J.; Bellouin, N.; Doutriaux-Boucher, M.; Gedney, N.; Halloran, P.; Hinton, T.; Hughes, J.; Jones, C.D.; Joshi, M.; Liddicoat, S.; et al. Development and evaluation of an Earth-System model—HadGEM2. Geosci. Model Dev. 2011, 4, 1051–1075. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Liu, T.; Meng, F.; Duan, Y.; Frankl, A.; Bao, A.; De Maeyer, P. Comparing bias correction methods used in downscaling precipitation and temperature from regional climate models: A case study from the Kaidu River Basin in Western China. Water 2018, 10, 1046. [Google Scholar] [CrossRef] [Green Version]
- Blaustein, A.R. Predicting climate-induced range shifts: Model differences and model reliability. Glob. Chang. Biol. 2006, 12, 1568–1584. [Google Scholar] [CrossRef] [Green Version]
- Breiman, L.E.O. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Cutler, D.R.; Edwards, T.C.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random forests for classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef]
- Duchesnay, F.P.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Maraun, D.; Widmann, M. Statistical Downscaling and Bias Correction for Climate Research. Stat. Downscal. Bias Correct. Clim. Res. 2018. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, J.M.; Maraun, D.; Widmann, M.; Huth, R.; Hertig, E.; Benestad, R.; Roessler, O.; Wibig, J.; Wilcke, R.; Kotlarski, S.; et al. An intercomparison of a large ensemble of statistical downscaling methods over Europe: Results from the VALUE perfect predictor cross-validation experiment. Int. J. Climatol. 2019, 39, 3750–3785. [Google Scholar] [CrossRef]
- Chen, J.; Brissette, F.P.; Leconte, R. Coupling statistical and dynamical methods for spatial downscaling of precipitation. Clim. Chang. 2012, 114, 509–526. [Google Scholar] [CrossRef]
- Ovalle-Rivera, O.; Läderach, P.; Bunn, C.; Obersteiner, M.; Schroth, G. Projected shifts in Coffea arabica suitability among major global producing regions due to climate change. PLoS ONE 2015, 10, e0124155. [Google Scholar] [CrossRef] [Green Version]
- Rehm, E.M.; Feeley, K.J. The inability of tropical cloud forest species to invade grasslands above treeline during climate change: Potential explanations and consequences. Ecography 2015, 38, 1167–1175. [Google Scholar] [CrossRef] [Green Version]
- Sekercioglu, C.H.; Schneider, S.H.; Fay, J.P.; Loarie, S.R. Climate change, elevational range shifts, and bird extinctions. Conserv. Biol. 2008, 22, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Feeley, K.J.; Silman, M.R. Land-use and climate change effects on population size and extinction risk of Andean plants. Glob. Chang. Biol. 2010, 16, 3215–3222. [Google Scholar] [CrossRef]
- Valencia, B.G.; Urrego, D.H.; Silman, M.R.; Bush, M.B. From ice age to modern: A record of landscape change in an Andean cloud forest. J. Biogeogr. 2010, 37, 1637–1647. [Google Scholar] [CrossRef]
- Bush, M.B.; Silman, M.R.; Urrego, D.H. 48,000 Years of Climate and Forest Change in a Biodiversity Hot Spot. Science 2004, 303, 827–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malcolm, J.R.; Markham, A.; Neilson, R.P.; Garaci, M. Estimated migration rates under scenarios of global climate change. J. Biogeogr. 2002, 29, 835–849. [Google Scholar] [CrossRef] [Green Version]
- Buytaert, W.; Bievre, B. De Water for cities: The impact of climate change and demographic growth in the tropical Andes. Water Resour. Res. 2012, 48, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Park, H.; Im, J.; Yoo, C.; Rhee, J.; Lee, B.; Kwon, C.G. Delineation of high resolution climate regions over the Korean Peninsula using machine learning approaches. PLoS ONE 2019, 14, e0223362. [Google Scholar] [CrossRef]
- Buytaert, W.; Cuesta-Camacho, F.; Tobón, C. Potential impacts of climate change on the environmental services of humid tropical alpine regions. Glob. Ecol. Biogeogr. 2011, 20, 19–33. [Google Scholar] [CrossRef]
- Balvanera, P.; Uriarte, M.; Almeida-Leñero, L.; Altesor, A.; DeClerck, F.; Gardner, T.; Hall, J.; Lara, A.; Laterra, P.; Peña-Claros, M.; et al. Ecosystem services research in Latin America: The state of the art. Ecosyst. Serv. 2012, 2, 56–70. [Google Scholar] [CrossRef]
- Woodward, F.I.; Lomas, M.R.; Kelly, C.K. Global climate and the distribution of plant biomes. Philos. Trans. R. Soc. B Biol. Sci. 2004, 359, 1465–1476. [Google Scholar] [CrossRef] [Green Version]
- Salazar, L.F.; Nobre, C.A.; Oyama, M.D. Climate change consequences on the biome distribution in tropical South America. Geophys. Res. Lett. 2007, 34, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Foley, J.A.; Asner, G.P.; Costa, M.H.; Coe, M.T.; Defries, R.; Gibbs, H.K.; Howard, E.A.; Olson, S.; Patz, J.; Ramankutty, N.; et al. Amazonia revealed: Forest degradation and loss of ecosystem goods and services in the Amazon Basin. Front. Ecol. Environ. 2007, 5, 25–32. [Google Scholar] [CrossRef]





| Peruvian Biomes | Area % | Plant Life Form | Humidity Level |
|---|---|---|---|
| EMF | 42.44 | forest | 8 |
| SDTF | 2.42 | forest | 7 |
| HP | 12.84 | grassland | 4 |
| P | 1.08 | grassland | 5 |
| MS | 8.06 | shrubland | 6 |
| PP | 5.85 | desert | 1 |
| GC | 2.32 | desert | 2 |
| XP | 0.91 | grassland | 3 |
| YF | 13.53 | forest | 7 |
| SW | 4.84 | forest | 9 |
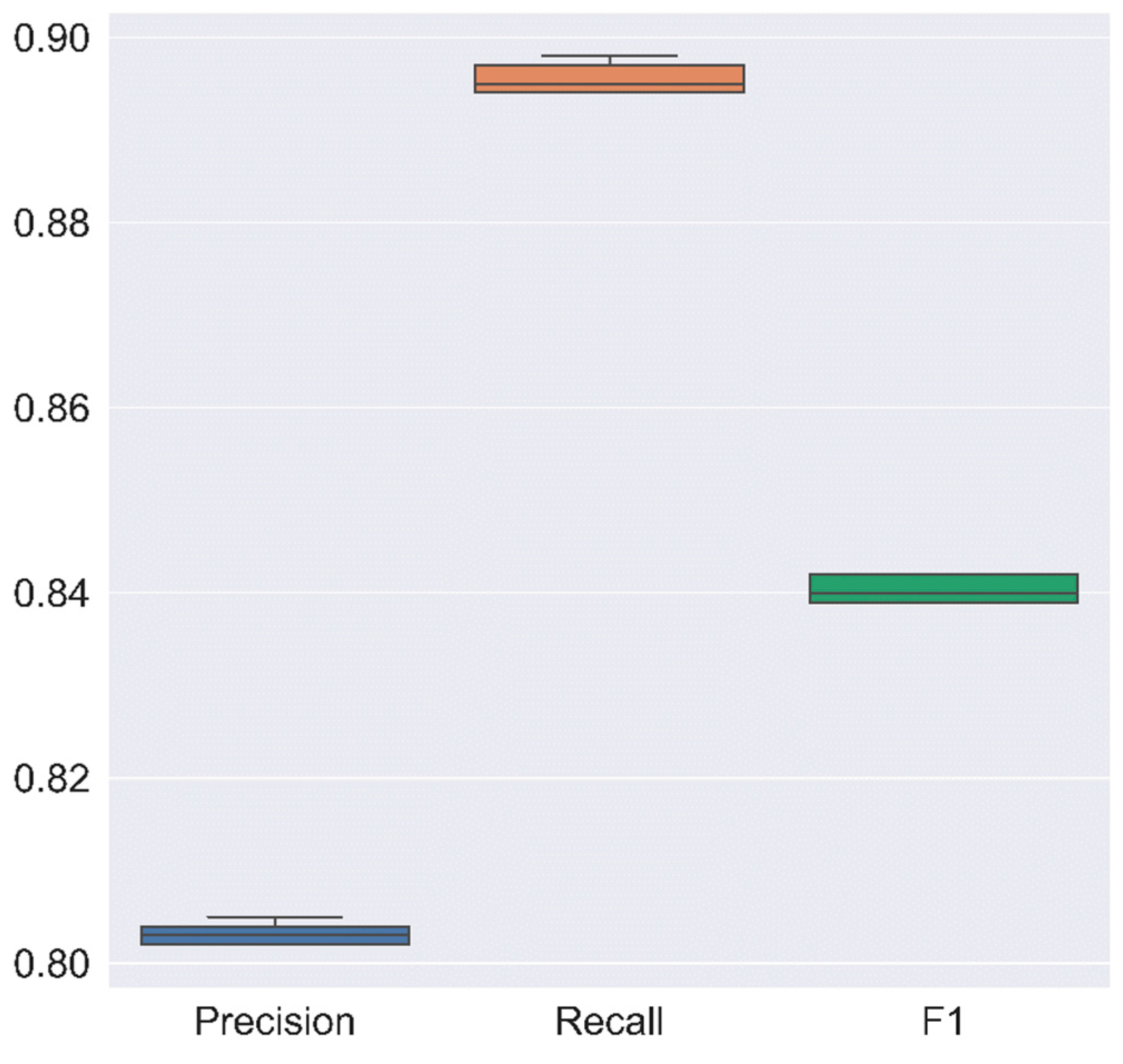
| K-Fold | Precision | Recall | F1 Score |
|---|---|---|---|
| RF | RF | RF | |
| 1 | 0.805 | 0.897 | 0.842 |
| 2 | 0.802 | 0.894 | 0.839 |
| 3 | 0.803 | 0.894 | 0.840 |
| 4 | 0.804 | 0.898 | 0.842 |
| 5 | 0.802 | 0.895 | 0.839 |
| Mean | 0.803 | 0.896 | 0.841 |
| Standard deviation | 0.001 | 0.002 | 0.001 |
| EMF | SDTF | HP | P | MS | PP | GC | XP | YF | SW | Recall | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| EMF | 98432 | 700 | 287 | 96 | 269 | 98 | 71 | 26 | 2155 | 7568 | 0.90 |
| SDTF | 16 | 5819 | 0 | 0 | 319 | 80 | 0 | 0 | 44 | 0 | 0.93 |
| HP | 13 | 1 | 28851 | 725 | 1217 | 0 | 2017 | 226 | 351 | 0 | 0.86 |
| P | 0 | 0 | 29 | 2521 | 69 | 0 | 5 | 0 | 105 | 0 | 0.92 |
| MS | 1 | 556 | 1127 | 450 | 17792 | 545 | 15 | 244 | 230 | 0 | 0.85 |
| PP | 7 | 113 | 0 | 0 | 389 | 14776 | 0 | 0 | 0 | 0 | 0.97 |
| GC | 0 | 0 | 879 | 23 | 6 | 0 | 4744 | 231 | 0 | 0 | 0.81 |
| XP | 0 | 0 | 34 | 0 | 69 | 0 | 116 | 2275 | 0 | 0 | 0.91 |
| YF | 526 | 181 | 349 | 505 | 275 | 0 | 0 | 0 | 33226 | 2 | 0.95 |
| SW | 980 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 11591 | 0.92 |
| Precision | 0.98 | 0.79 | 0.91 | 0.58 | 0.87 | 0.95 | 0.68 | 0.76 | 0.92 | 0.60 |
| EMF | SDTF | HP | P | MS | PP | GC | XP | YF | SW | |
|---|---|---|---|---|---|---|---|---|---|---|
| EMF | 96.4 (95.7–98.4) | 0.11 | 0.24 | 0 | 0.18 | 0.05 | 0 | 0.01 | 0.24 | 2.83 (0.6–3.5) |
| SDTF | 28.85 (24.6–32.6) | 66.55 (60.8–70.7) | 0 | 0 | 2.86 (1.9–5.5) | 1.4 (0.8–1.9) | 0 | 0 | 0.39 | 0 |
| HP | 0.04 | 0 | 78.35 (75.5–86.7) | 1.94 (0.7–4.9) | 12.48 (9.1–17.1) | 0 | 0.74 (0.3–1.0) | 1.19 (0.7–4.7) | 1.69 (0.5–1.7) | 0 |
| P | 0 | 0 | 7.82 (5.9–9.6) | 60.01 (59.9–62.1) | 20.36 (20.2–25.2) | 0 | 0.03 | 0 | 9.65 (8.7–10.0) | 0 |
| MS | 0.05 | 4.56 (4.2–4.9) | 0.82 | 0.51 | 89.82 (89.2–89.9) | 1.58 | 0 | 0.05 | 2.55 (1.2–3.3) | 0 |
| PP | 0.81 | 7.62 (4.2–10.8) | 0 | 0 | 9.78 (8.6–11.9) | 81.79 (76.2–86.9) | 0 | 0 | 0 | 0 |
| GC | 0 | 0 | 41.99 (39.4–44.4) | 0.34 | 0.09 | 0 | 44.95 (43.3–45.1) | 12.58 | 0 | 0 |
| XP | 0 | 0 | 11.65 (0.5–16.1) | 0 | 16.41 (11.5–21.2) | 0.06 | 0.59 (0.5–1.3) | 66.34 (66–87.3) | 0 | 0 |
| YF | 16.38 (14.8–22.92) | 0.79 (0.7–1.5) | 0.04 | 0.09 | 0.85 (0.7–4.1) | 0 | 0 | 0 | 81.85 (70.9–83.6) | 0 |
| SW | 69.19 (56.6–94.2) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 30.81 (5.7–43.3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zevallos, J.; Lavado-Casimiro, W. Climate Change Impact on Peruvian Biomes. Forests 2022, 13, 238. https://doi.org/10.3390/f13020238
Zevallos J, Lavado-Casimiro W. Climate Change Impact on Peruvian Biomes. Forests. 2022; 13(2):238. https://doi.org/10.3390/f13020238
Chicago/Turabian StyleZevallos, Jose, and Waldo Lavado-Casimiro. 2022. "Climate Change Impact on Peruvian Biomes" Forests 13, no. 2: 238. https://doi.org/10.3390/f13020238
APA StyleZevallos, J., & Lavado-Casimiro, W. (2022). Climate Change Impact on Peruvian Biomes. Forests, 13(2), 238. https://doi.org/10.3390/f13020238