
The Functional Structure of Tropical Plant Communities and Soil Properties Enhance Ecosystem Functioning and Multifunctionality in Different Ecosystems in Ghana

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
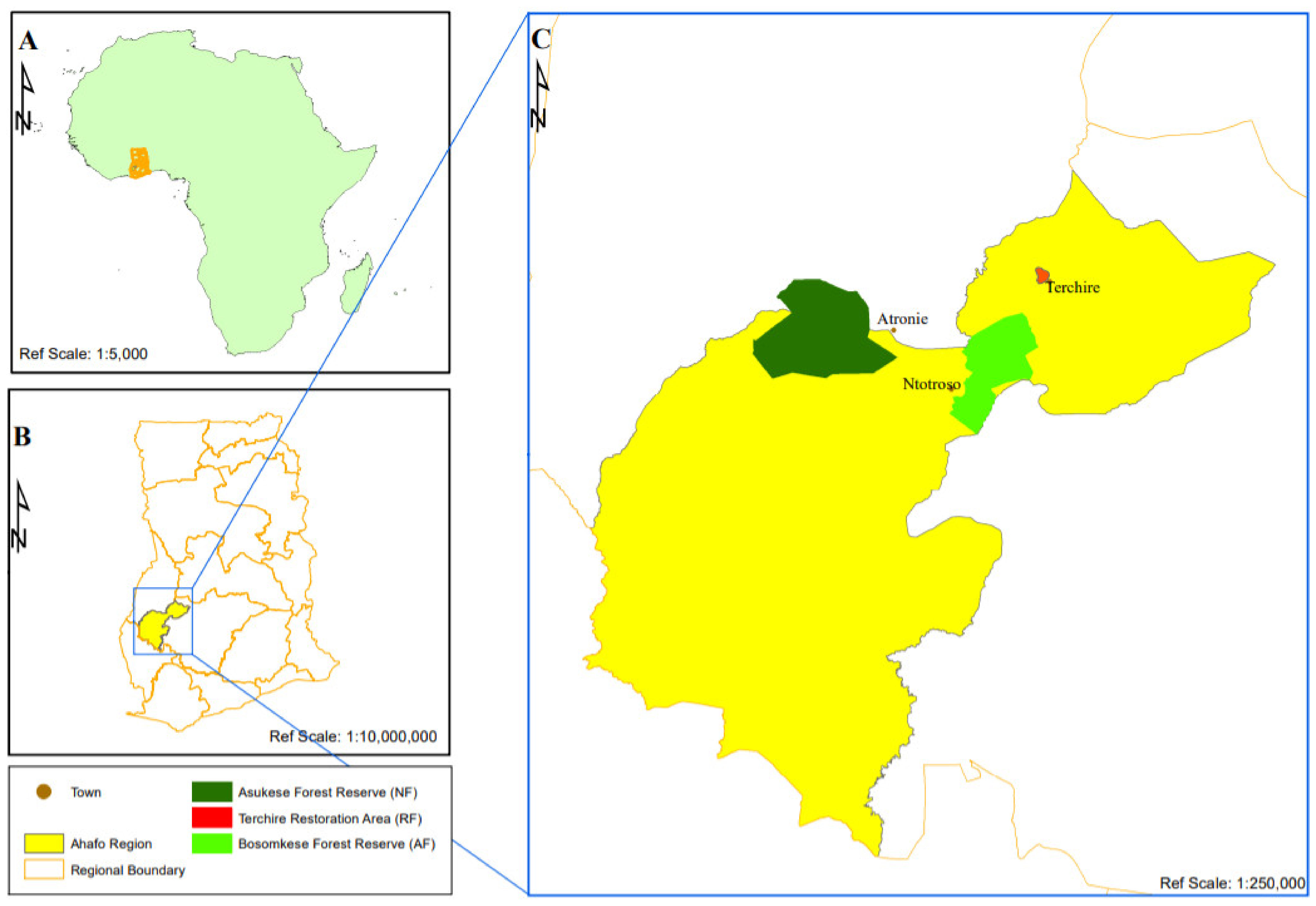
2.1. Study Site Description
2.2. Data Collection
2.3. Ecosystem Functions and Multifunctionality
2.4. Functional Composition and Diversity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ | Trait | Abb | Unit | Functional Role | References |
|---|---|---|---|---|---|
| Leaf | Specific leaf area | SLA | m2 kg−1 | Reflects whole-plant growth and photosynthetic efficiency of a species | [61,62] |
| Leaf carbon and nitrogen ratio | C/N | It is an indicator of nitrogen-use efficiency reflecting the metabolic status of C and N in leaves | [63,64] | ||
| Isotopic carbon fraction | δ13C | ‰ | Reflects gas exchange and water-use efficiency | [65] | |
| Stem | Stem dry matter content | SDMC | mg g−1 | Relates above-ground storage of carbon and stimulates plants’ resistance to physical hazards | [33,66] |
| Seed | Seed mass | Smass | mg | Moderates a trade-off between plants’ rate of seed production per unit mass invested in reproduction and the mass of each offspring | [67] |
| Whole plant | Plant height | Phg | m | Reflects a species ability to utilise light and above the ground competition | [68,69] |
2.5. Soil Properties Measurement
2.6. Data Analysis
3. Results
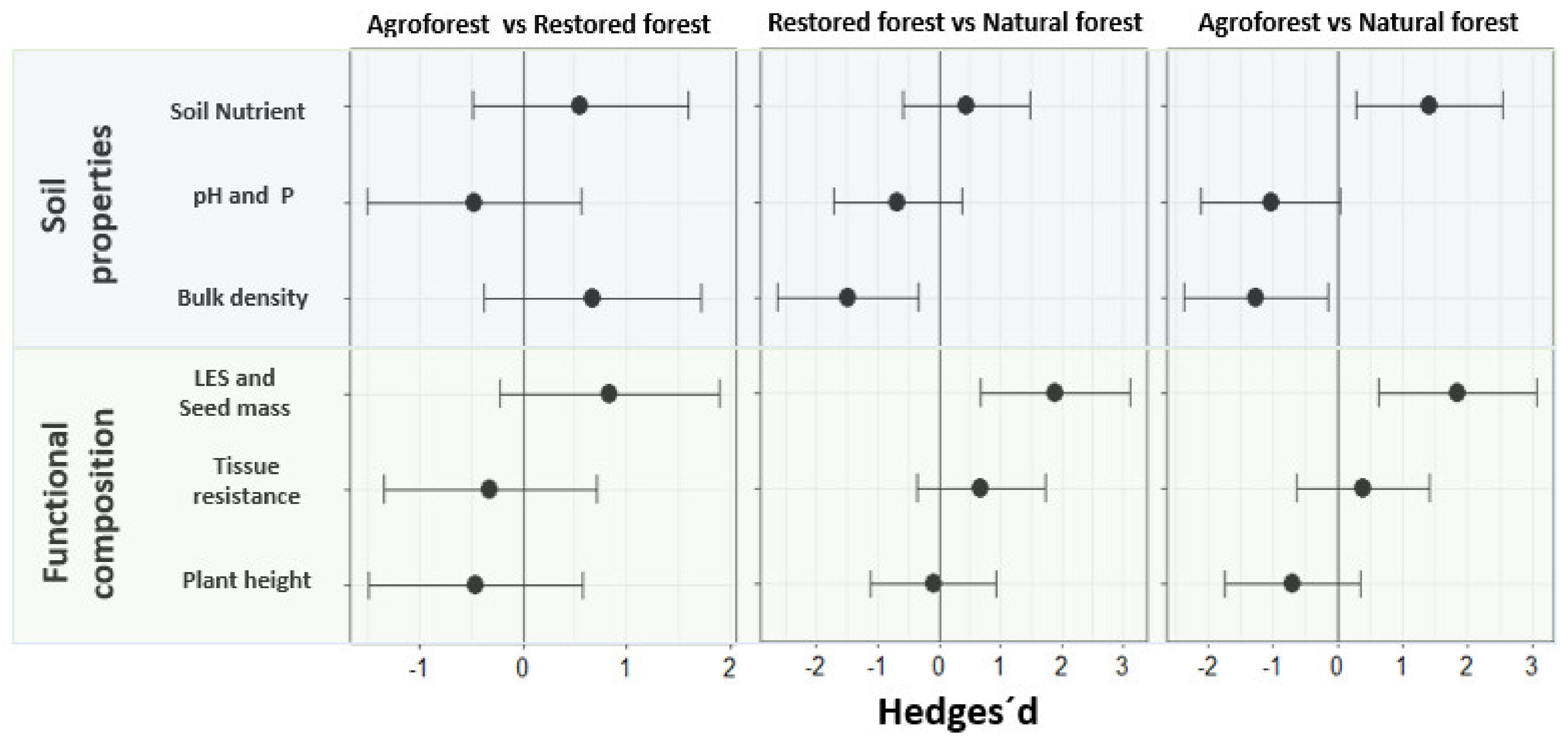
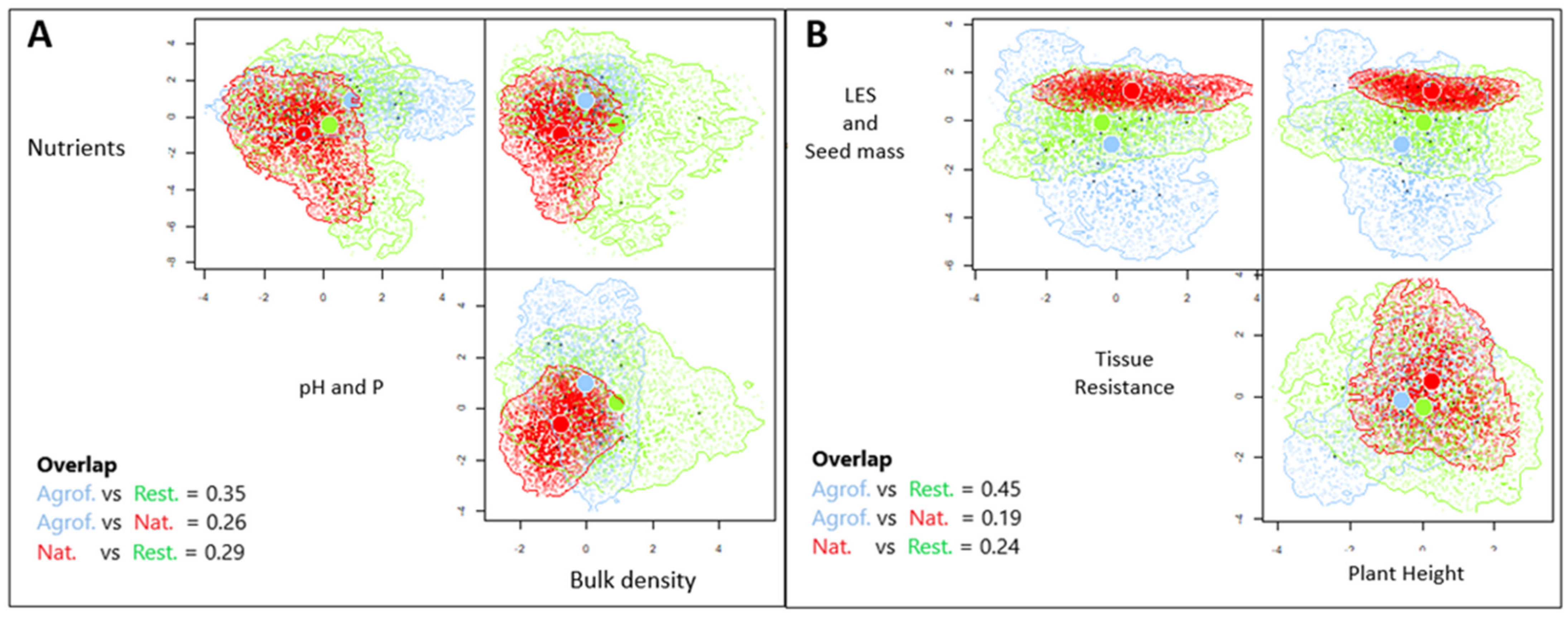
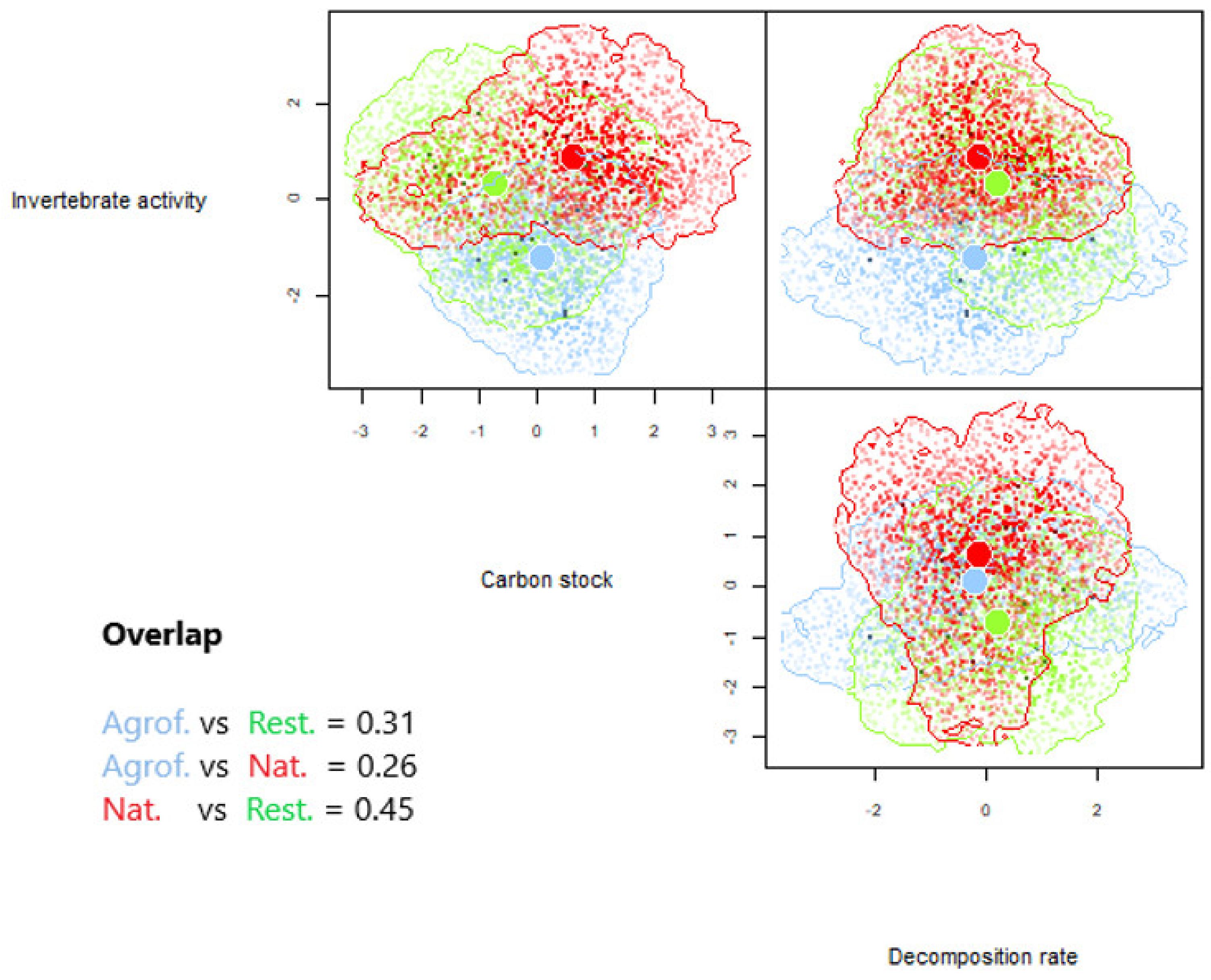
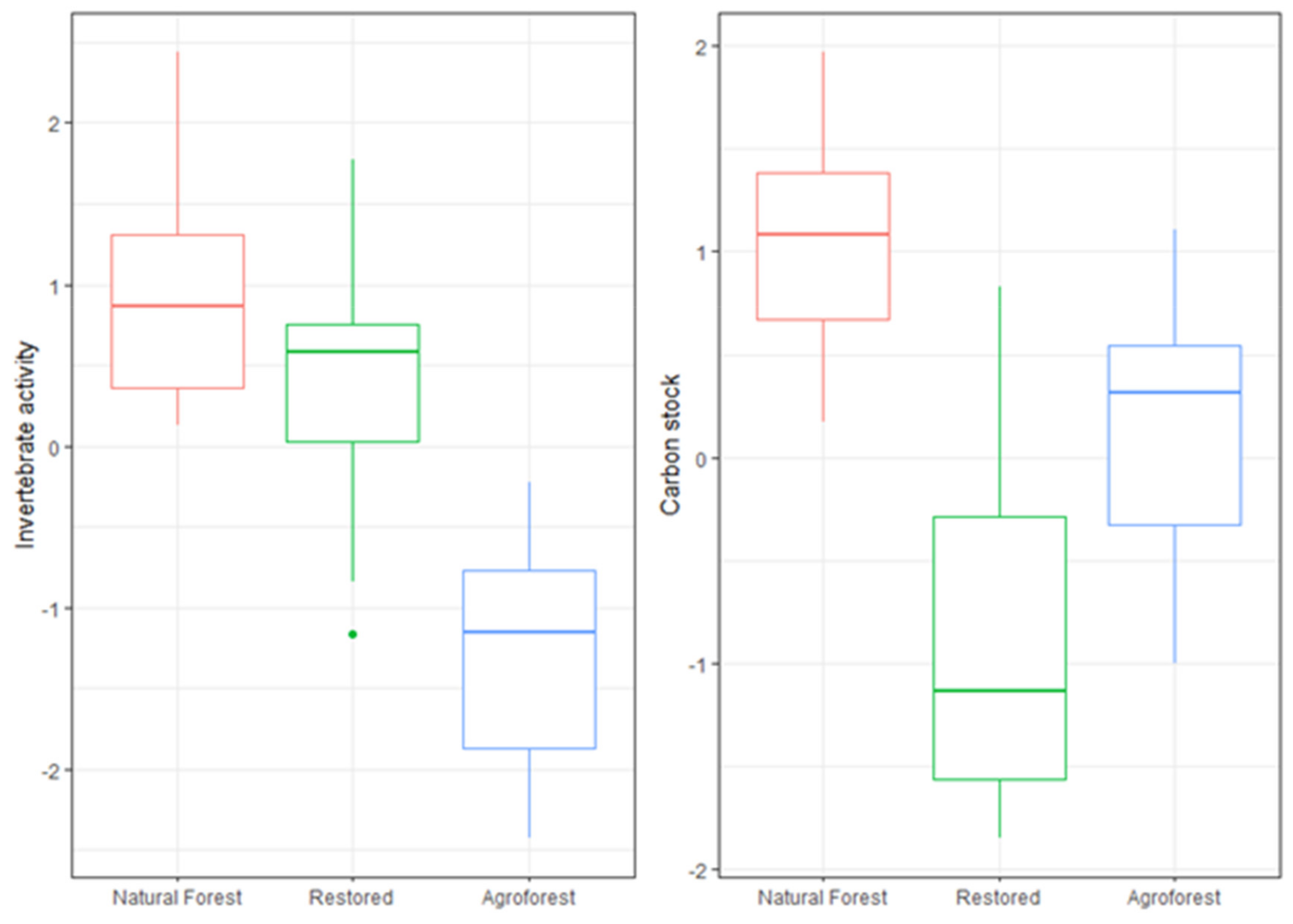
3.1. The Relationship of Ecosystem Types with Soil Parameters and Functional Composition
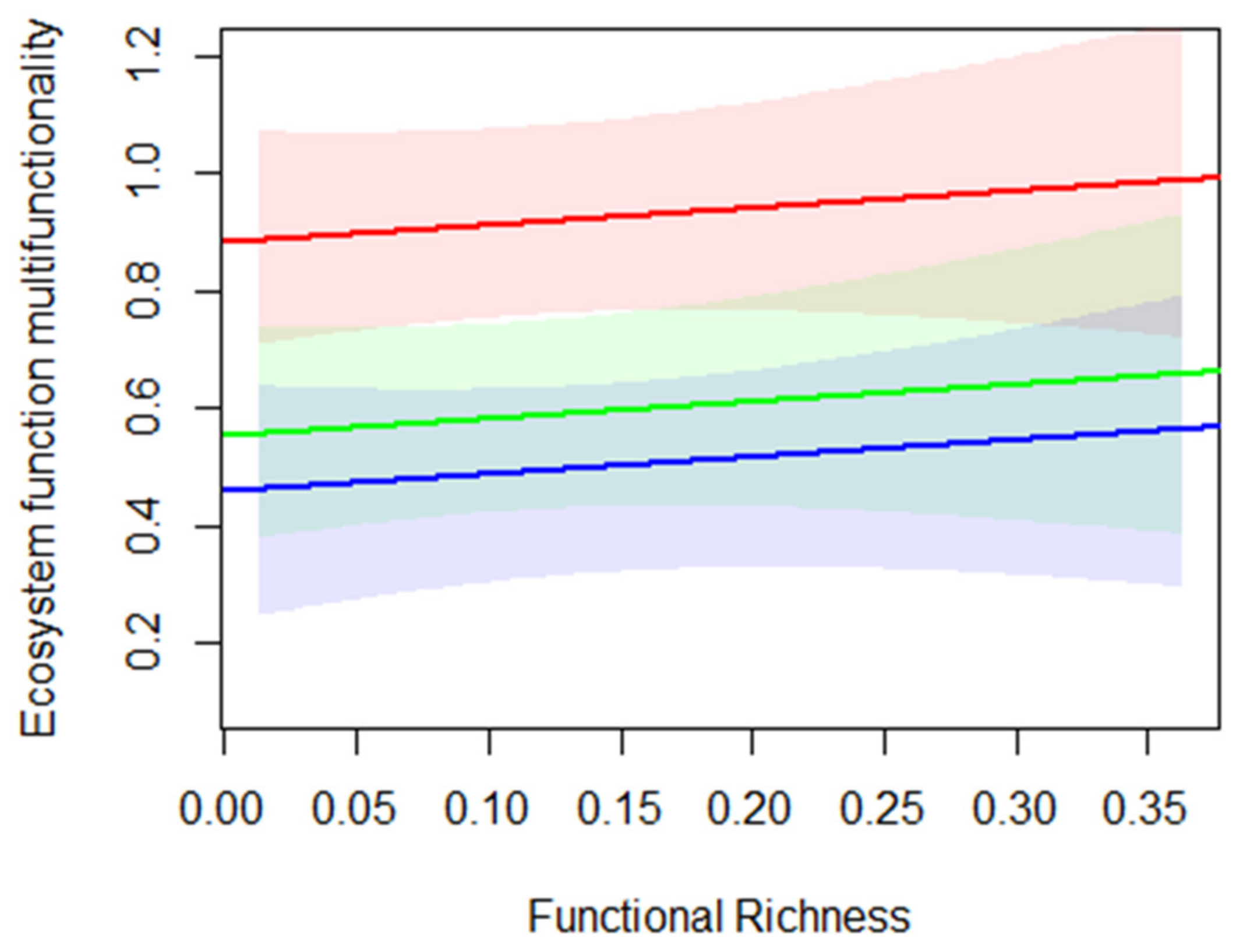
3.2. The Influence of Functional Structure and Soil Properties on Ecosystem Services and Multifunctionality across Ecosystems
4. Discussion
4.1. Ecosystem Type Relationship with Soil Parameters and Functional Composition
4.2. Functional Structure and Soil Properties Influence Ecosystem Services and Multifunctionality across Ecosystems
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| Species | Family |
|---|---|
| Antrocaryon micraster A. Chev. & Guillaumin | Anacardiaceae |
| Alstonia boonei De Wild. | Apocynaceae |
| Funtumia elastica (Preuss) Stapf | Apocynaceae |
| Holarrhena floribunda (G. Don) T. Durand & Schinz | Apocynaceae |
| Rauvolfia vomitoria Afzel. | Apocynaceae |
| Spathodea campanulata P. Beauv. | Bignoniaceae |
| Distemonanthus benthamianus Baill. | Leguminosae |
| Terminalia ivorensis A. Chev. | Combretaceae |
| Terminalia suberba Engl. & Diels | Combretaceae |
| Antidesma laciniatum Müll. Arg. | Phyllanthaceae |
| Macaranga barteri Müll. Arg | Euphorbiaceae |
| Ricinodendron heudelotii (Baill.) Heckel | Euphorbiaceae |
| Albizia zygia (DC.) J. F. Macbr. | Leguminosae |
| Anthocleista nobilis G. Don | Gentianaceae |
| Amphimas pterocarpoides Harms | Leguminosae |
| Bombax buonopozense P. Beauv. | Malvaceae |
| Ceiba pentandra (L.) Gaertn. | Malvaceae |
| Cola gigantea A. Chev. | Malvaceae |
| Mansonia altissima (A Chev.) A Chev. | Malvaceae |
| Pterygota macrocarpa K. Schum. | Malvaceae |
| Triplochiton scleroxylon K. Schum. | Malvaceae |
| Azadirachta indica A. Juss. | Meliaceae |
| Entandrophragma angolense (Welw.) C. DC. | Meliaceae |
| Entandrophragma utile (Dawe & Sprague) Sprague | Meliaceae |
| Khaya anthotheca (Welw.) C. DC. | Meliaceae |
| Piptadeniastrum africanum (Hook.f.) Brenan | Leguminosae |
| Antiaris toxicaria (Lesch.) | Moraceae |
| Ficus exasperata Vahl | Moraceae |
| Milicia excelsa (Welw.) C. C. Berg | Moraceae |
| Morus mesozygia Stapf | Moraceae |
| Pycnanthus angolensis (Welw.) Warb. | Myristicaceae |
| Margaritaria discoidea (Baill.) G. L. Webster | Phyllanthaceae |
| Morinda lucida Benth. | Rubiaceae |
| Blighia sapida K.D. Koenig | Sapindaceae |
| Chrysophyllum albidum G. Don | Sapotaceae |
| Sterculia rhinopetala K. Schum. | Malvaceae |
| Celtis adolfi-friderici Engl. | Cannabaceae |
| Celtis mildbraedii Engl. | Cannabaceae |


| Plant Abundance and Measured Functional Traits | Natural Forest | Restored Forest | Agroforestry System |
|---|---|---|---|
| Plant abundance (n/ha) | 157 | 119 | 153 |
| Specific leaf area (m/kg2) | 80.646 | 66.326 | 47.018 |
| Leaf carbon and nitrogen ratio | 13.292 | 12.192 | 12.480 |
| Isotopic carbon fraction (‰) | 31.117 | 31.174 | 30.523 |
| Stem dry matter content (mg/g) | 0.423 | 0.411 | 0.418 |
| Seed mass (mg) | 0.907 | 0.569 | 0.666 |
| Plant height (m) | 14.532 | 14.655 | 16.120 |
References
- Malhi, Y.; Franklin, J.; Seddon, N.; Solan, M.; Turner, M.G.; Field, C.B.; Knowlton, N. Climate change and ecosystems: Threats, opportunities and solutions. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Geest, K.; de Sherbinin, A.; Kienberger, S.; Zommers, Z.; Sitati, A.; Roberts, E.; James, R. The impacts of climate change on ecosystem services and resulting losses and damages to people and society. In Loss and Damage from Climate Change; Mechler, R., Bouwer, L.M., Schinko, T., Surminski, S., Linnerooth-Bayer, J., Eds.; Springer: Cham, Swtizerland, 2019; pp. 221–236. [Google Scholar]
- Lavorel, S. Plant functional effects on ecosystem services. J. Ecol. 2013, 101, 4–8. [Google Scholar] [CrossRef]
- Hillman, J.R.; Lundquist, C.J.; Thrush, S.F. The challenges associated with connectivity in ecosystem processes. Front. Mar. Sci. 2018, 5, 364. [Google Scholar] [CrossRef] [Green Version]
- Balvanera, P.; Pfisterer, A.B.; Buchmann, N.; He, J.S.; Nakashizuka, T.; Raffaelli, D.; Schmid, B. Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecol. Lett. 2006, 9, 1146–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loreau, M.; Naeem, S.; Inchausti, P.; Bengtsson, J.; Grime, J.P.; Hector, A.; Wardle, D.A. Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science 2001, 294, 804–808. [Google Scholar] [CrossRef] [Green Version]
- Hanisch, M.; Schweiger, O.; Cord, A.F.; Volk, M.; Knapp, S. Plant functional traits shape multiple ecosystem services, their trade-offs and synergies in grasslands. J. Appl. Ecol. 2020, 57, 1535–1550. [Google Scholar] [CrossRef]
- Birkhofer, K.; Andersson, G.K.; Bengtsson, J.; Bommarco, R.; Dänhardt, J.; Ekbom, B.; Ekroos, J.; Hahn, T.; Hedlund, K.; Jönsson, A.M.; et al. Relationships between multiple biodiversity components and ecosystem services along a landscape complexity gradient. Biol. Conserv. 2018, 218, 247–253. [Google Scholar] [CrossRef]
- Maestre, F.T.; Castillo-Monroy, A.P.; Bowker, M.A.; Ochoa-Hueso, R. Species richness effects on ecosystem multifunctionality depend on evenness, composition and spatial pattern. J. Ecol. 2012, 100, 317–330. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Hillebrand, H.; Jonsson, P.R. Multiple functions increase the importance of biodiversity for overall ecosystem functioning. Ecology 2008, 89, 1223–1231. [Google Scholar] [CrossRef]
- Mouillot, D.; Villéger, S.; Scherer-Lorenzen, M.; Mason, N.W. Functional structure of biological communities predicts ecosystem multifunctionality. PLoS ONE 2011, 6, e17476. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.H.; Cadotte, M.W.; Burgess, K.S.; Liu, J.; Tan, S.L.; Zou, J.Y.; Xu, K.; Li, D.Z.; Gao, L.M. Greater than the sum of the parts: How the species composition in different forest strata influence ecosystem function. Ecol. Lett. 2019, 22, 1449–1461. [Google Scholar] [CrossRef]
- Gross, N.; Bagousse-Pinguet, Y.L.; Liancourt, P.; Berdugo, M.; Gotelli, N.J.; Maestre, F.T. Functional trait diversity maximizes ecosystem multifunctionality. Nat. Ecol. Evol. 2017, 1, 0132. [Google Scholar] [CrossRef] [PubMed]
- Violle, C.; Navas, M.L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Nock, C.A.; Vogt, R.J.; Beisner, B.E. Functional traits. In eLS; John Wiley & Sons, Ltd.: Chichester, UK, 2016; pp. 1–8. [Google Scholar] [CrossRef]
- Adler, P.B.; Salguero-Gómez, R.; Compagnoni, A.; Hsu, J.S.; Ray-Mukherjee, J.; Mbeau-Ache, C.; Franco, M. Functional traits explain variation in plant life history strategies. Proc. Natl. Acad. Sci. USA 2014, 111, 740–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaturvedi, R.K.; Tripathi, A.; Raghubanshi, A.S.; Singh, J.S. Functional traits indicate a continuum of tree drought strategies across a soil water availability gradient in a tropical dry forest. For. Ecol. Manag. 2021, 482, 118740. [Google Scholar] [CrossRef]
- Feizabadi, M.F.; Tahmasebi, P.; Broujeni, E.A.; Ebrahimi, A.; Omidipour, R. Functional diversity, functional composition and functional β diversity drive aboveground biomass across different bioclimatic rangelands. Basic Appl. Ecol. 2021, 52, 68–81. [Google Scholar] [CrossRef]
- Vargas-Hernández, J.G.; Zdunek-Wielgołaska, J. Urban green infrastructure as a tool for controlling the resilience of urban sprawl. Environ. Dev. Sustain. 2021, 23, 1335–1354. [Google Scholar] [CrossRef] [Green Version]
- Hanif, M.; Yu, Q.; Rao, X.; Shen, W. Disentangling the contributions of plant taxonomic and functional diversities in shaping aboveground biomass of a restored forest landscape in Southern China. Plants 2019, 8, 612. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Jiang, L.; Zhang, Y. Relationships between functional diversity and aboveground biomass production in the Northern Tibetan alpine grasslands. Sci. Rep. 2016, 6, 34105. [Google Scholar] [CrossRef] [Green Version]
- Finegan, B.; Peña-Claros, M.; de Oliveira, A.; Ascarrunz, N.; Bret-Harte, M.S.; Carreño-Rocabado, G.; Casanoves, F.; Dıaz, S.; Velepucha, P.E.; Fernandez, F.; et al. Does functional trait diversity predict above-ground biomass and productivity of tropical forests? Testing three alternative hypotheses. J. Ecol. 2015, 103, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Su, J.; Li, S.; Liu, W.; Lang, X. Functional diversity drives ecosystem multifunctionality in a Pinus yunnanensis natural secondary forest. Sci. Rep. 2019, 9, 6979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, W.; Huang, J.; Xu, H.; Zang, R.; Ding, Y.; Li, Y.; Li, Y.; Lin, M.; Wang, J.; Zhang, C. Plant functional traits are the mediators in regulating effects of abiotic site conditions on aboveground carbon stock-evidence from a 30 ha tropical forest plot. Front. Plant Sci. 2019, 9, 1958. [Google Scholar] [CrossRef] [PubMed]
- Fyllas, N.M.; Michelaki, C.; Galanidis, A.; Evangelou, E.; Zaragoza-Castells, J.; Dimitrakopoulos, P.G.; Tsadilas, C.; Arianoutsou, M.; Lloyd, J. Functional trait variation among and within species and plant functional types in mountainous mediterranean forests. Front. Plant Sci. 2020, 11, 212. [Google Scholar] [CrossRef] [Green Version]
- Westoby, M.; Falster, D.S.; Moles, A.T.; Vesk, P.A.; Wright, I.J. Plant ecological strategies: Some leading dimensions of variation between species. Annu. Rev. Ecol. Syst. 2002, 33, 125–159. [Google Scholar] [CrossRef] [Green Version]
- Díaz, S.; Kattge, J.; Cornelissen, J.H.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Prentice, I.C.; et al. The global spectrum of plant form and function. Nature 2016, 529, 167–171. [Google Scholar] [CrossRef]
- Dıaz, S.; Cabido, M. Vive la différence: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Reich, P.B. The world-wide ‘fast–slow’plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- De la Riva, E.G.; Prieto, I.; Villar, R. The leaf economic spectrum drives leaf litter decomposition in Mediterranean forests. Plant Soil 2019, 435, 353–366. [Google Scholar] [CrossRef]
- Conti, G.; Díaz, S. Plant functional diversity and carbon storage–an empirical test in semi-arid forest ecosystems. J. Ecol. 2013, 101, 18–28. [Google Scholar] [CrossRef]
- Laureto, L.M.O.; Cianciaruso, M.V.; Samia, D.S.M. Functional diversity: An overview of its history and applicability. Nat. Conserv. 2015, 13, 112–116. [Google Scholar] [CrossRef] [Green Version]
- De la Riva, E.G.; Marañón, T.; Violle, C.; Villar, R.; Pérez-Ramos, I.M. Biogeochemical and ecomorphological niche segregation of Mediterranean woody species along a local gradient. Front. Plant Sci. 2017, 8, 1242. [Google Scholar] [CrossRef] [PubMed]
- Lazarova, S.; Coyne, D.; Rodríguez, M.G.; Peteira, B.; Ciancio, A. Functional diversity of soil nematodes in relation to the impact of agriculture—A review. Diversity 2021, 13, 64. [Google Scholar] [CrossRef]
- Valladares, F.; Bastias, C.C.; Godoy, O.; Granda, E.; Escudero, A. Species coexistence in a changing world. Front. Plant Sci. 2015, 6, 866. [Google Scholar] [CrossRef] [PubMed]
- De Bello, F.; Lavorel, S.; Díaz, S.; Harrington, R.; Cornelissen, J.H.; Bardgett, R.D.; Matty, P.B.; Cipriotti, P.; Feld, C.K.; Hering, D.; et al. Towards an assessment of multiple ecosystem processes and services via functional traits. Biodivers. Conserv. 2010, 19, 2873–2893. [Google Scholar] [CrossRef]
- Birkhofer, K.; Fliessbach, A.; Gavín-Centol, M.P.; Hedlund, K.; Ingimarsdóttir, M.; Jørgensen, H.B.; Kozjek, K.; Meyer, S.; Montserrat, M.; Moreno, S.S. Conventional agriculture and not drought alters relationships between soil biota and functions. Sci. Rep. 2021, 11, 23975. [Google Scholar] [CrossRef]
- Birkhofer, K.; Smith, H.G.; Weisser, W.W.; Wolters, V.; Gossner, M.M. Land-use effects on the functional distinctness of arthropod communities. Ecography 2015, 38, 889–900. [Google Scholar] [CrossRef]
- Jayathilake, H.M.; Prescott, G.W.; Carrasco, L.R.; Rao, M.; Symes, W.S. Drivers of deforestation and degradation for 28 tropical conservation landscapes. Ambio 2021, 50, 215–228. [Google Scholar] [CrossRef]
- Jung, M.; Rowhani, P.; Scharlemann, J.P. Impacts of past abrupt land change on local biodiversity globally. Nat. Commun. 2019, 10, 5474. [Google Scholar] [CrossRef] [Green Version]
- Acheampong, E.O.; Macgregor, C.J.; Sloan, S.; Sayer, J. Deforestation is driven by agricultural expansion in Ghana’s forest reserves. Sci. Afr. 2019, 5, e00146. [Google Scholar] [CrossRef]
- Symes, W.S.; Edwards, D.P.; Miettinen, J.; Rheindt, F.E.; Carrasco, L.R. Combined impacts of deforestation and wildlife trade on tropical biodiversity are severely underestimated. Nat. Commun. 2018, 9, 4052. [Google Scholar] [CrossRef] [Green Version]
- Oduro, K.A.; Mohren, G.M.J.; Pena-Claros, M.; Kyereh, B.; Arts, B. Tracing forest resource development in Ghana through forest transition pathways. Land Use Policy 2015, 48, 63–72. [Google Scholar] [CrossRef]
- Damptey, F.G.; de la Riva, E.G.; Birkhofer, K. Trade-offs and synergies between food and fodder production and other ecosystem services in an actively restored forest, natural forest and an agroforestry system in Ghana. Front. For. Glob. Chang. 2021, 4, 47. [Google Scholar] [CrossRef]
- Damptey, F.G.; Birkhofer, K.; Nsiah, P.K.; de la Riva, E.G. Soil properties and biomass attributes in a former gravel mine area after two decades of forest restoration. Land 2020, 9, 209. [Google Scholar] [CrossRef]
- Sharma, S.; MacKenzie, R.A.; Tieng, T.; Soben, K.; Tulyasuwan, N.; Resanond, A.; Blate, G.; Litton, C.M. The impacts of degradation, deforestation and restoration on mangrove ecosystem carbon stocks across Cambodia. Sci. Total Environ. 2020, 706, 135416. [Google Scholar] [CrossRef] [PubMed]
- Bernal, B.; Murray, L.T.; Pearson, T.R. Global carbon dioxide removal rates from forest landscape restoration activities. Carbon Balance Manag. 2018, 13, 22. [Google Scholar] [CrossRef] [PubMed]
- Aponte, C.; García, L.V.; Marañón, T. Tree species effects on nutrient cycling and soil biota: A feedback mechanism favouring species coexistence. For. Ecol. Manag. 2013, 309, 36–46. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Ramos, I.M.; Roumet, C.; Cruz, P.; Blanchard, A.; Autran, P.; Garnier, E. Evidence for a ‘plant community economics spectrum’ driven by nutrient and water limitations in a Mediterranean rangeland of southern France. J. Ecol. 2012, 100, 1315–1327. [Google Scholar] [CrossRef] [Green Version]
- Hawthorne, W.D.N.; Gyakari, N. Photoguide for the Forest Trees of Ghana: A Tree-Spotter’s Field Guide for Identifying the Largest Trees; Oxford Forestry Institute—Department of Plant Sciences: Oxford, UK, 2006. [Google Scholar]
- Gatti, R.C.; Castaldi, S.; Lindsell, J.A.; Coomes, D.A.; Marchetti, M.; Maesano, M.; Di Paola, A.; Paparella, F.; Valentini, R. The impact of selective logging and clearcutting on forest structure, tree diversity and above-ground biomass of African tropical forests. Ecol. Res. 2015, 30, 119–132. [Google Scholar] [CrossRef]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef]
- Zanne, A.E.; Lopez-Gonzalez, G.; Coomes, D.A.; Ilic, J.; Jansen, S.; Lewis, S.L.; Miller, R.B.; Swenson, N.G.; Wiemann, M.C.; Chave, J. Global Wood Density Database. Dryad Digital Repository. 2009. Available online: https://datadryad.org/stash/dataset/doi:10.5061/dryad.234 (accessed on 28 August 2021).
- Lewis, S.L.; Sonké, B.; Sunderland, T.; Begne, S.K.; Lopez-Gonzalez, G.; van der Heijden, G.M.; Philips, O.L.; Affum-Baffoe, K.; Baker, T.R.; Banin, L.; et al. Above-ground biomass and structure of 260 African tropical forests. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120295. [Google Scholar] [CrossRef] [Green Version]
- Brockerhoff, E.G.; Barbaro, L.; Castagneyrol, B.; Forrester, D.I.; Gardiner, B.; González-Olabarria, J.R.; Lyver, P.O.; Meurisse, N.; Oxbrough, A.; Taki, H.; et al. Forest biodiversity, ecosystem functioning and the provision of ecosystem services. Biodivers. Conserv. 2017, 26, 3005–3035. [Google Scholar] [CrossRef] [Green Version]
- Keuskamp, J.A.; Dingemans, B.J.; Lehtinen, T.; Sarneel, J.M.; Hefting, M.M. Tea Bag Index: A novel approach to collect uniform decomposition data across ecosystems. Methods Ecol. Evol. 2013, 4, 1070–1075. [Google Scholar] [CrossRef]
- Kattge, J.; Diaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Bönisch, G.; Garnier, E.; Westoby, M.; Reich, P.B.; Wright, I.J.; et al. TRY—A global database of plant traits. Glob. Chang. Biol. 2011, 17, 2905–2935. [Google Scholar] [CrossRef]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef]
- Glozer, K. Protocol for Leaf Image Analysis—Surface Area. 2008. Available online: http://ucanr.edu/sites/fruittree/files/49325.pdf (accessed on 17 September 2021).
- Perez-Harguindeguy, N.; Diaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. Corrigendum to: New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2016, 64, 715–716. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Wang, Z.; Li, S.; Lü, X.; Wang, X.; Han, X. Changes in specific leaf area of dominant plants in temperate grasslands along a 2500-km transect in northern China. Sci. Rep. 2017, 7, 10780. [Google Scholar] [CrossRef] [PubMed]
- Prado, J.; Schiavini, I.; Vale, V.; Lopes, S.; Arantes, C.; Oliveira, A.P. Functional leaf traits of understory species: Strategies to different disturbance severities. Braz. J. Biol. 2015, 75, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; He, N.; Liu, C.; Xu, L.; Chen, Z.; Li, Y.; Wang, R.; Yu, G.; Sun, W.; Xiao, C.; et al. Variation and evolution of C:N ratio among different organs enable plants to adapt to N-limited environments. Glob. Chang. Biol. 2020, 26, 2534–2543. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yang, G.; Yang, X.; Li, Z.; Feng, H.; Xu, B.; Zhao, X. Monitoring ratio of carbon to nitrogen (C/N) in wheat and barley leaves by using spectral slope features with branch-and-bound algorithm. Sci. Rep. 2018, 8, 10034. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Sang, W.; Bai, F.; Brennan, E.; Diekman, M.; Liu, Y.; Li, L.; Marples, A.; Shi, H.; Sui, Z.; et al. Large-scale α-diversity patterns in plants and ground beetles (Coleoptera: Carabidae) indicate a high biodiversity conservation value of China’s restored temperate forest landscapes. Divers. Distrib. 2019, 25, 1613–1624. [Google Scholar] [CrossRef] [Green Version]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; ter Steege, H.; Morgan, H.D.; van der Heijiden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef] [Green Version]
- Moles, A.T.; Westoby, M. Seed size and plant strategy across the whole life cycle. Oikos 2006, 113, 91–105. [Google Scholar] [CrossRef]
- Galán Díaz, J.; de la Riva, E.G.; Funk, J.L.; Vilà, M. Functional segregation of resource-use strategies of native and invasive plants across Mediterranean biome communities. Biol. Invasions 2021, 23, 253–266. [Google Scholar] [CrossRef]
- Moles, A.T.; Warton, D.I.; Warman, L.; Swenson, N.G.; Laffan, S.W.; Zanne, A.E.; Pitman, A.; Hemmings, F.A.; Leishman, M.R. Global patterns in plant height. J. Ecol. 2009, 97, 923–932. [Google Scholar] [CrossRef]
- Larjavaara, M.; Muller-Landau, H.C. Measuring tree height: A quantitative comparison of two common field methods in a moist tropical forest. Methods Ecol. Evol. 2013, 4, 793–801. [Google Scholar] [CrossRef]
- Garnier, E.; Cortez, J.; Billès, G.; Navas, M.L.; Roumet, C.; Debussche, M.; Laurent, G.; Blanchard, A.; Aubry, D.; Bellmann, A.; et al. Plant functional markers capture ecosystem properties during secondary succession. Ecology 2004, 85, 2630–2637. [Google Scholar] [CrossRef]
- De Bello, F.; Carmona, C.P.; Lepš, J.; Szava-Kovats, R.; Pärtel, M. Functional diversity through the mean trait dissimilarity: Resolving shortcomings with existing paradigms and algorithms. Oecologia 2016, 180, 933–940. [Google Scholar] [CrossRef]
- Karadimou, E.K.; Kallimanis, A.S.; Tsiripidis, I.; Dimopoulos, P. Functional diversity exhibits a diverse relationship with area, even a decreasing one. Sci. Rep. 2016, 6, 35420. [Google Scholar] [CrossRef] [Green Version]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring from multiple traits functional diversity. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Villéger, S.; Mason, N.W.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [Green Version]
- Rao, C.R. Diversity and dissimilarity coefficients: A unified approach. Theor. Popul. Biol. 1982, 21, 24–43. [Google Scholar] [CrossRef]
- Botta-Dukát, Z. Rao’s quadratic entropy as a measure of functional diversity based on multiple traits. J. Veg. Sci. 2005, 16, 533–540. [Google Scholar] [CrossRef]
- Laughlin, D.C. The intrinsic dimensionality of plant traits and its relevance to community assembly. J. Ecol. 2014, 102, 186–193. [Google Scholar] [CrossRef]
- Torchiano, M. Effsize: Efficient Effect Size Computation (R Package); R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Blonder, B. Hypervolume concepts in niche-and trait-based ecology. Ecography 2018, 41, 1441–1455. [Google Scholar] [CrossRef]
- Blonder, B.; Nogués-Bravo, D.; Borregaard, M.K.; Donoghue II, J.C.; Jørgensen, P.M.; Kraft, N.J.; Lessard, J.P.; Morueta-Holme, N.; Sandel, B.; Svenning, J.C.; et al. Linking environmental filtering and disequilibrium to biogeography with a community climate framework. Ecology 2015, 96, 972–985. [Google Scholar] [CrossRef] [Green Version]
- Grueber, C.E.; Nakagawa, S.; Laws, R.J.; Jamieson, I.G. Multimodel inference in ecology and evolution: Challenges and solutions. J. Evol. Biol. 2011, 24, 699–711. [Google Scholar] [CrossRef]
- Barton, K.; Barton, M.K. Package ‘Mumin’; Version 1. 2015. Available online: https://cran.r-project.org/package/MuMIn/MuMIn.pdf (accessed on 17 September 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; ISBN 3-900051-07-0. [Google Scholar]
- Moir, J.; Jordan, P.; Moot, D.; Lucas, R. Phosphorus response and optimum pH ranges of twelve pasture legumes grown in an acid upland New Zealand soil under glasshouse conditions. J. Soil Sci. Plant Nutr. 2016, 16, 438–460. [Google Scholar] [CrossRef] [Green Version]
- Oliveras, I.; Bentley, L.; Fyllas, N.M.; Gvozdevaite, A.; Shenkin, A.F.; Peprah, T.; Morandi, P.; Peixoto, K.S.; Boakye, M.; Adu-Bredu, S.; et al. The influence of taxonomy and environment on leaf trait variation along tropical abiotic gradients. Front. For. Glob. Chang. 2020, 3, 18. [Google Scholar] [CrossRef] [Green Version]
- De la Riva, E.G.; Pérez-Ramos, I.M.; Tosto, A.; Navarro-Fernández, C.M.; Olmo, M.; Marañón, T.; Villar, R. Disentangling the relative importance of species occurrence, abundance and intraspecific variability in community assembly: A trait-based approach at the whole-plant level in Mediterranean forests. Oikos 2016, 125, 354–363. [Google Scholar] [CrossRef] [Green Version]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- De la Riva, E.G.; Violle, C.; Pérez-Ramos, I.M.; Marañón, T.; Navarro-Fernández, C.M.; Olmo, M.; Villar, R. A multidimensional functional trait approach reveals the imprint of environmental stress in Mediterranean woody communities. Ecosystems 2018, 21, 248–262. [Google Scholar] [CrossRef] [Green Version]
- Gvozdevaite, A.; Oliveras, I.; Domingues, T.F.; Peprah, T.; Boakye, M.; Afriyie, L.; Peixoto, K.S.; de Farias, J.; de Oliveira, E.A.; Farias, C.C.A.; et al. Leaf-level photosynthetic capacity dynamics in relation to soil and foliar nutrients along forest–Savanna boundaries in Ghana and Brazil. Tree Physiol. 2018, 38, 1912–1925. [Google Scholar] [CrossRef] [PubMed]
- Neyret, M.; Bentley, L.P.; Oliveras, I.; Marimon, B.S.; Marimon-Junior, B.H.; Almeida de Oliveira, E.; Passos, F.B.; Ccoscco, R.C.; dos Santos, J.; Reis, S.M.; et al. Examining variation in the leaf mass per area of dominant species across two contrasting tropical gradients in light of community assembly. Ecol. Evol. 2016, 6, 5674–5689. [Google Scholar] [CrossRef] [Green Version]
- Sterck, F.; Markesteijn, L.; Schieving, F.; Poorter, L. Functional traits determine trade-offs and niches in a tropical forest community. Proc. Natl. Acad. Sci. USA 2011, 108, 20627–20632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornelissen, J.H.C. A triangular relationship between leaf size and seed size among woody species: Allometry, ontogeny, ecology and taxonomy. Oecologia 1999, 118, 248–255. [Google Scholar] [CrossRef]
- Pérez-Ramos, I.M.; Gómez-Aparicio, L.; Villar, R.; García, L.V.; Maranon, T. Seedling growth and morphology of three oak species along field resource gradients and seed mass variation: A seedling age-dependent response. J. Veg. Sci. 2010, 21, 419–437. [Google Scholar] [CrossRef] [Green Version]
- Quero, J.L.; Villar, R.; Marañón, T.; Zamora, R.; Poorter, L. Seed-mass effects in four Mediterranean Quercus species (Fagaceae) growing in contrasting light environments. Am. J. Bot. 2007, 94, 1795–1803. [Google Scholar] [CrossRef] [Green Version]
- Moles, A.T.; Falster, D.S.; Leishman, M.R.; Westoby, M. Small-seeded species produce more seeds per square metre of canopy per year, but not per individual per lifetime. J. Ecol. 2004, 92, 384–396. [Google Scholar] [CrossRef]
- Moles, A.T.; Westoby, M. Seedling survival and seed size: A synthesis of the literature. J. Ecol. 2004, 92, 372–383. [Google Scholar] [CrossRef]
- Wright, S.J.; Stoner, K.E.; Beckman, N.; Corlett, R.T.; Dirzo, R.; Muller-Landau, H.C.; Nunez-Iturri, G.; Peres, C.A.; Wang, B.C. The plight of large animals in tropical forests and the consequences for plant regeneration. Biotropica 2007, 39, 289–291. [Google Scholar] [CrossRef]
- Opuni-Frimpong, E.; Gabienu, E.; Adusu, D.; Opuni-Frimpong, N.Y.; Damptey, F.G. Plant diversity, conservation significance, and community structure of two protected areas under different governance. Trees For. People 2021, 4, 100082. [Google Scholar] [CrossRef]
- Padmakumar, B.; Sreekanth, N.P.; Shanthiprabha, V.; Paul, J.; Sreedharan, K.; Augustine, T.; Jayasooryan, K.K.; Rameshan, M.; Mohan, M.; Ramasamy, E.V.; et al. Tree biomass and carbon density estimation in the tropical dry forest of Southern Western Ghats, India. iForest-Biogeosci. For. 2018, 11, 534. [Google Scholar] [CrossRef] [Green Version]
- Schuldt, A.; Ebeling, A.; Kunz, M.; Staab, M.; Guimarães-Steinicke, C.; Bachmann, D.; Buchmann, N.; Durka, W.; Fichtner, A.; Fornoff, F.; et al. Multiple plant diversity components drive consumer communities across ecosystems. Nat. Commun. 2019, 10, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenninger, E.J.; Inouye, R.S. Insect community response to plant diversity and productivity in a sagebrush–steppe ecosystem. J. Arid Environ. 2008, 72, 24–33. [Google Scholar] [CrossRef]
- McColloch, J.W. The reciprocal relation of soil and insects. Ecology 1922, 3, 288–301. [Google Scholar] [CrossRef]
- Gunstone, T.; Cornelisse, T.; Klein, K.; Dubey, A.; Donley, N. Pesticides and Soil Invertebrates: A Hazard Assessment. Front. Environ. Sci. 2021, 9, 122. [Google Scholar] [CrossRef]
- Amprako, L.; Stenchly, K.; Wiehle, M.; Nyarko, G.; Buerkert, A. Arthropod Communities in Urban Agricultural Production Systems under Different Irrigation Sources in the Northern Region of Ghana. Insects 2020, 11, 488. [Google Scholar] [CrossRef]
- Raven, P.H.; Wagner, D.L. Agricultural intensification and climate change are rapidly decreasing insect biodiversity. Proc. Natl. Acad. Sci. USA 2021, 118, e2002548117. [Google Scholar] [CrossRef]
- Zhao, Z.H.; Hui, C.; He, D.H.; Li, B.L. Effects of agricultural intensification on ability of natural enemies to control aphids. Sci. Rep. 2015, 5, 8024. [Google Scholar] [CrossRef] [Green Version]
- Schjoerring, J.K.; Cakmak, I.; White, P.J. Plant nutrition and soil fertility: Synergies for acquiring global green growth and sustainable development. Plant Soil 2019, 434, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Chatzistathis, T.; Therios, I. How soil nutrient availability influences plant biomass and how biomass stimulation alleviates heavy metal toxicity in soils: The cases of nutrient use efficient genotypes and phytoremediators, respectively. In Biomass Now-Cultivation and Utilization; Matovic, D.M., Ed.; IntechOpen: Rijeka, Croatia, 2013; pp. 427–448. [Google Scholar]
- Måren, I.E.; Sharma, L.N. Seeing the wood for the trees: Carbon storage and conservation in temperate forests of the Himalayas. For. Ecol. Manag. 2021, 487, 119010. [Google Scholar] [CrossRef]
- Kothandaraman, S.; Dar, J.A.; Sundarapandian, S.; Dayanandan, S.; Khan, M.L. Ecosystem-level carbon storage and its links to diversity, structural and environmental drivers in tropical forests of Western Ghats, India. Sci. Rep. 2020, 10, 13444. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.L.; Kuchma, O.; Krutovsky, K.V. Mixed-species versus monocultures in plantation forestry: Development, benefits, ecosystem services and perspectives for the future. Glob. Ecol. Conserv. 2018, 1, e00419. [Google Scholar] [CrossRef]
- Heilpern, S.A.; Anujan, K.; Osuri, A.; Naeem, S. Positive correlations in species functional contributions drive the response of multifunctionality to biodiversity loss. Proc. R. Soc. B 2020, 287, 20192501. [Google Scholar] [CrossRef]
- Zirbel, C.R.; Bassett, T.; Grman, E.; Brudvig, L.A. Plant functional traits and environmental conditions shape community assembly and ecosystem functioning during restoration. J. Appl. Ecol. 2017, 54, 1070–1079. [Google Scholar] [CrossRef] [Green Version]
- Manning, P.; van der Plas, F.; Soliveres, S.; Allan, E.; Maestre, F.T.; Mace, G.; Whittingham, M.J.; Fischer, M. Redefining ecosystem multifunctionality. Nat. Ecol. Evol. 2018, 2, 427–436. [Google Scholar] [CrossRef]
- Shen, Y.; Yu, S.; Lian, J.; Shen, H.; Cao, H.; Lu, H.; Ye, W. Tree aboveground carbon storage correlates with environmental gradients and functional diversity in a tropical forest. Sci. Rep. 2016, 6, 25304. [Google Scholar] [CrossRef] [Green Version]
- Cavanaugh, K.C.; Gosnell, J.S.; Davis, S.L.; Ahumada, J.; Boundja, P.; Clark, D.B.; Mugerwa, B.; Jansen, P.A.; O’Brien, T.G.; Rovero, F.; et al. Carbon storage in tropical forests correlates with taxonomic diversity and functional dominance on a global scale. Glob. Ecol. Biogeogr. 2014, 23, 563–573. [Google Scholar] [CrossRef]
- Mensah, S.; Veldtman, R.; Assogbadjo, A.E.; Glèlè Kakaï, R.; Seifert, T. Tree species diversity promotes aboveground carbon storage through functional diversity and functional dominance. Ecol. Evol. 2016, 6, 7546–7557. [Google Scholar] [CrossRef]
- Tilman, D.; Isbell, F.; Cowles, J.M. Biodiversity and ecosystem functioning. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 471–493. [Google Scholar] [CrossRef]
- Campos, R.I.; Vasconcelos, H.L.; Ribeiro, S.P.; Neves, F.S.; Soares, J.P. Relationship between tree size and insect assemblages associated with Anadenanthera macrocarpa. Ecography 2006, 29, 442–450. [Google Scholar] [CrossRef]
- Marques, E.S.D.A.; Price, P.W.; Cobb, N.S. Resource abundance and insect herbivore diversity on woody fabaceous desert plants. Environ. Entomol. 2000, 29, 696–703. [Google Scholar] [CrossRef] [Green Version]
- Damptey, F.G.; Frimpong, B.F.; Debrah, D.K.; Agro, P.P.; Wiafe, E.D. Vegetation attributes drive the taxonomic richness and functional composition of beetles and spiders in mountainous urban green spaces. Energy Ecol. Environ. 2022, 7, 1–13. [Google Scholar] [CrossRef]
- Wein, A.; Bauhus, J.; Bilodeau-Gauthier, S.; Scherer-Lorenzen, M.; Nock, C.; Staab, M. Tree species richness promotes invertebrate herbivory on congeneric native and exotic tree saplings in a young diversity experiment. PLoS ONE 2016, 11, e0168751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulnik, J.; Plantureux, S.; Thery, M.; Baude, M.; Delattre, M.; van Reeth, C.; Villerd, J.; Michelot-Antalik, A. Floral trait functional diversity is related to soil characteristics and positively influences pollination function in semi-natural grasslands. Agric. Ecosyst. Environ. 2020, 301, 107033. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Carscadden, K.; Mirotchnick, N. Beyond species: Functional diversity and the maintenance of ecological processes and services. J. Appl. Ecol. 2011, 48, 1079–1087. [Google Scholar] [CrossRef]
- Ola, A.; Lovelock, C.E. Decomposition of mangrove roots depends on the bulk density they grew in. Plant Soil 2021, 460, 177–187. [Google Scholar] [CrossRef]
- Nosalewicz, A.; Lipiec, J. The effect of compacted soil layers on vertical root distribution and water uptake by wheat. Plant Soil 2014, 375, 229–240. [Google Scholar] [CrossRef]
- Kozlowski, T.T. Soil compaction and growth of woody plants. Scand. J. For. Res. 1999, 14, 596–619. [Google Scholar] [CrossRef]
- Blakely, J.K.; Neher, D.A.; Spongberg, A.L. Soil invertebrate and microbial communities, and decomposition as indicators of polycyclic aromatic hydrocarbon contamination. Appl. Soil Ecol. 2002, 21, 71–88. [Google Scholar] [CrossRef]
- Dissanayaka, D.M.; Dhananjaya, V.P.; Kosgollegedara, E.J.; Karthigayini, S. Impact of Improved Aeration on Decomposition Rate of Enriched Compost. Int. J. Trend Sci. Res. Dev. 2021, 5, 852–857. [Google Scholar]
- Ola, A.; Gauthier, A.R.; Xiong, Y.; Lovelock, C.E. The roots of blue carbon: Responses of mangrove stilt roots to variation in soil bulk density. Biol. Lett. 2019, 15, 20180866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrick, J.E.; Lal, R. Soil physical property changes during dung decomposition in a tropical pasture. Soil Sci. Soc. Am. J. 1995, 59, 908–912. [Google Scholar] [CrossRef]
- Mensah, S.; van der Plas, F.; Noulèkoun, F. Do functional identity and divergence promote aboveground carbon differently in tropical semi-arid forests and savannas? Ecosphere 2021, 12, e03563. [Google Scholar] [CrossRef]
- Le Bagousse-Pinguet, Y.; Soliveres, S.; Gross, N.; Torices, R.; Berdugo, M.; Maestre, F.T. Phylogenetic, functional, and taxonomic richness have both positive and negative effects on ecosystem multifunctionality. Proc. Natl. Acad. Sci. USA 2019, 116, 8419–8424. [Google Scholar] [CrossRef] [Green Version]
- Grman, E.; Zirbel, C.R.; Bassett, T.; Brudvig, L.A. Ecosystem multifunctionality increases with beta diversity in restored prairies. Oecologia 2018, 188, 837–848. [Google Scholar] [CrossRef]
- The Plant List. Version 1.1. 2013. Available online: http://www.theplantlist.org/ (accessed on 1 January 2022).




| Soil Properties | Source of Variance | ||
|---|---|---|---|
| PC1 | PC2 | PC3 | |
| pH | 4.0 | 61.0 | 7.0 |
| N | 50.0 | 14.0 | 23.0 |
| P | 15.0 | 62.0 | 15.0 |
| K | 55.0 | 2.0 | 4.0 |
| Na | 47.0 | 14.0 | 8.0 |
| Mg | 14.0 | 17.0 | 38.0 |
| SOC | 42.0 | 21.0 | 49.0 |
| BD | 4.0 | 37.0 | 72.0 |
| % variance | 32.3 | 22.9 | 14.8 |
| Functional composition | |||
| SLA | 53.0 | 24.0 | 12.0 |
| SDMC | 2.0 | 64.0 | 44.0 |
| Seed mass | 46.0 | 33.0 | 33.0 |
| C:N | 37.0 | 56.0 | 23.0 |
| δ13C | 52.0 | 30.0 | 36.0 |
| Plant height | 29.0 | 13.0 | 71.0 |
| % variance | 34.3 | 20.7 | 16.7 |
| Ecosystem service proxies | |||
| Carbon stock | 10.0 | 78.0 | 46.0 |
| Predator no. | 68.0 | 27.0 | 26.0 |
| Decomposer no. | 72.0 | 13.0 | 20.0 |
| Decomposition rate | 2.0 | 55.0 | 83.0 |
| % variance | 39.7 | 30.3 | 23.0 |
| Ecosystem Services | Factors | Relation | R2 | AICc | ΔAICc |
|---|---|---|---|---|---|
| Invertebrate activity | Null Model Site+Frich | + | 0.52 | 80.36 71.42 | 0.00 |
| Carbon stock | Null Model Site+Frich | + | 0.58 | 72.28 60.94 | 0.00 |
| Decomposition Rate | Null Model Soil BD | - | 0.24 | 67.97 64.38 | 0.00 |
| Multifunctionality | Null Model Site+Frich | + | 0.47 | 10.57 3.82 | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damptey, F.G.; Birkhofer, K.; Oliveras Menor, I.; de la Riva, E.G. The Functional Structure of Tropical Plant Communities and Soil Properties Enhance Ecosystem Functioning and Multifunctionality in Different Ecosystems in Ghana. Forests 2022, 13, 297. https://doi.org/10.3390/f13020297
Damptey FG, Birkhofer K, Oliveras Menor I, de la Riva EG. The Functional Structure of Tropical Plant Communities and Soil Properties Enhance Ecosystem Functioning and Multifunctionality in Different Ecosystems in Ghana. Forests. 2022; 13(2):297. https://doi.org/10.3390/f13020297
Chicago/Turabian StyleDamptey, Frederick Gyasi, Klaus Birkhofer, Imma Oliveras Menor, and Enrique G. de la Riva. 2022. "The Functional Structure of Tropical Plant Communities and Soil Properties Enhance Ecosystem Functioning and Multifunctionality in Different Ecosystems in Ghana" Forests 13, no. 2: 297. https://doi.org/10.3390/f13020297
APA StyleDamptey, F. G., Birkhofer, K., Oliveras Menor, I., & de la Riva, E. G. (2022). The Functional Structure of Tropical Plant Communities and Soil Properties Enhance Ecosystem Functioning and Multifunctionality in Different Ecosystems in Ghana. Forests, 13(2), 297. https://doi.org/10.3390/f13020297