
Diversity of Endophytes in the Botryosphaeriaceae Differs on Anacardiaceae in Disturbed and Undisturbed Ecosystems in South Africa


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Sites, Tree Health Assessment and Sampling
2.2. Fungal Isolations
2.3. DNA Extraction, PCR Amplification and Sequencing
2.4. DNA Sequence and Phylogenetic Analyses
3. Results
3.1. Tree Health Assessment
3.2. Fungal Isolation and Preliminary Identification
3.3. Phylogenetic Analyses and Confirmation of Species Identification
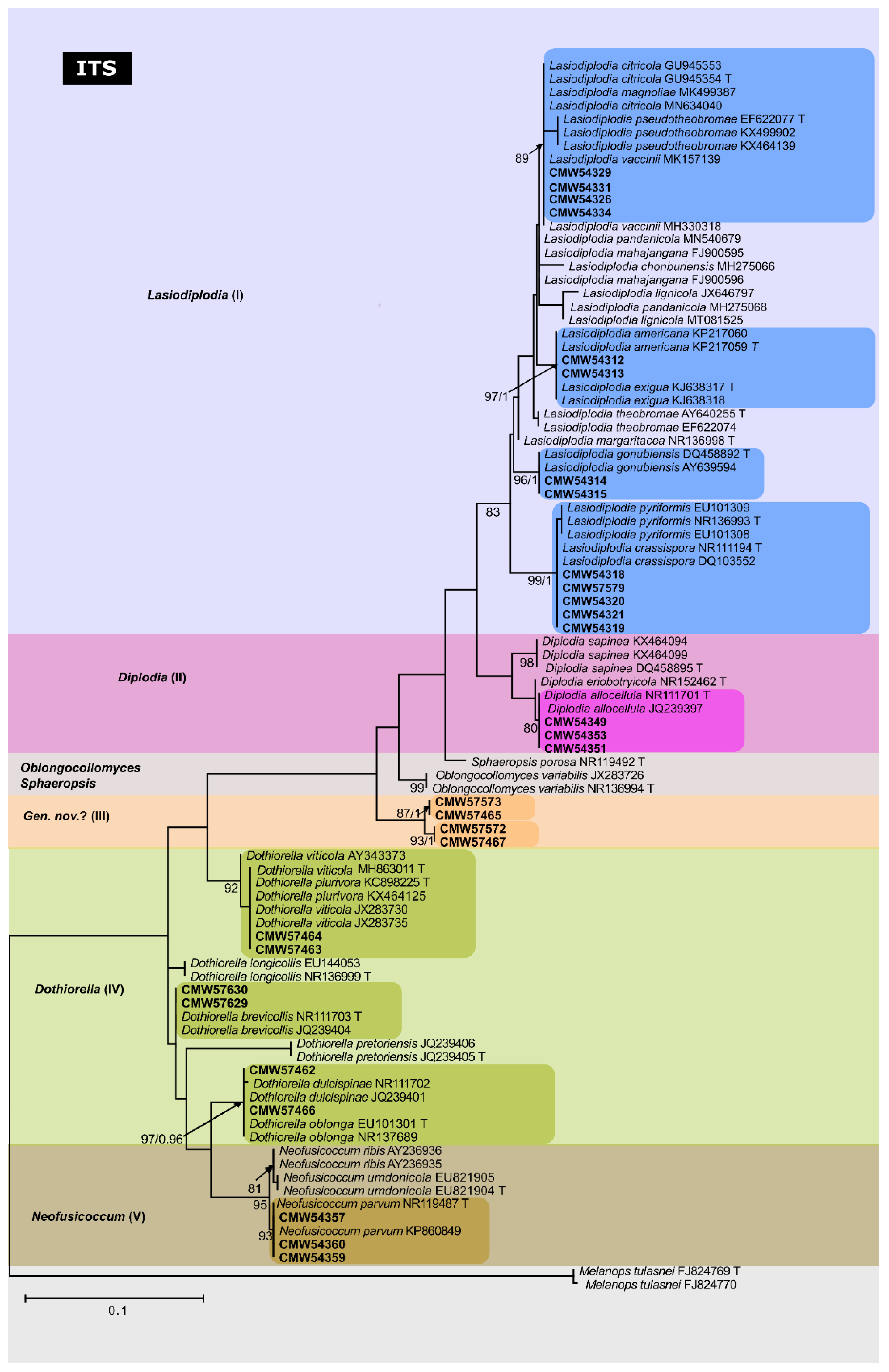
3.3.1. ITS Phylogeny
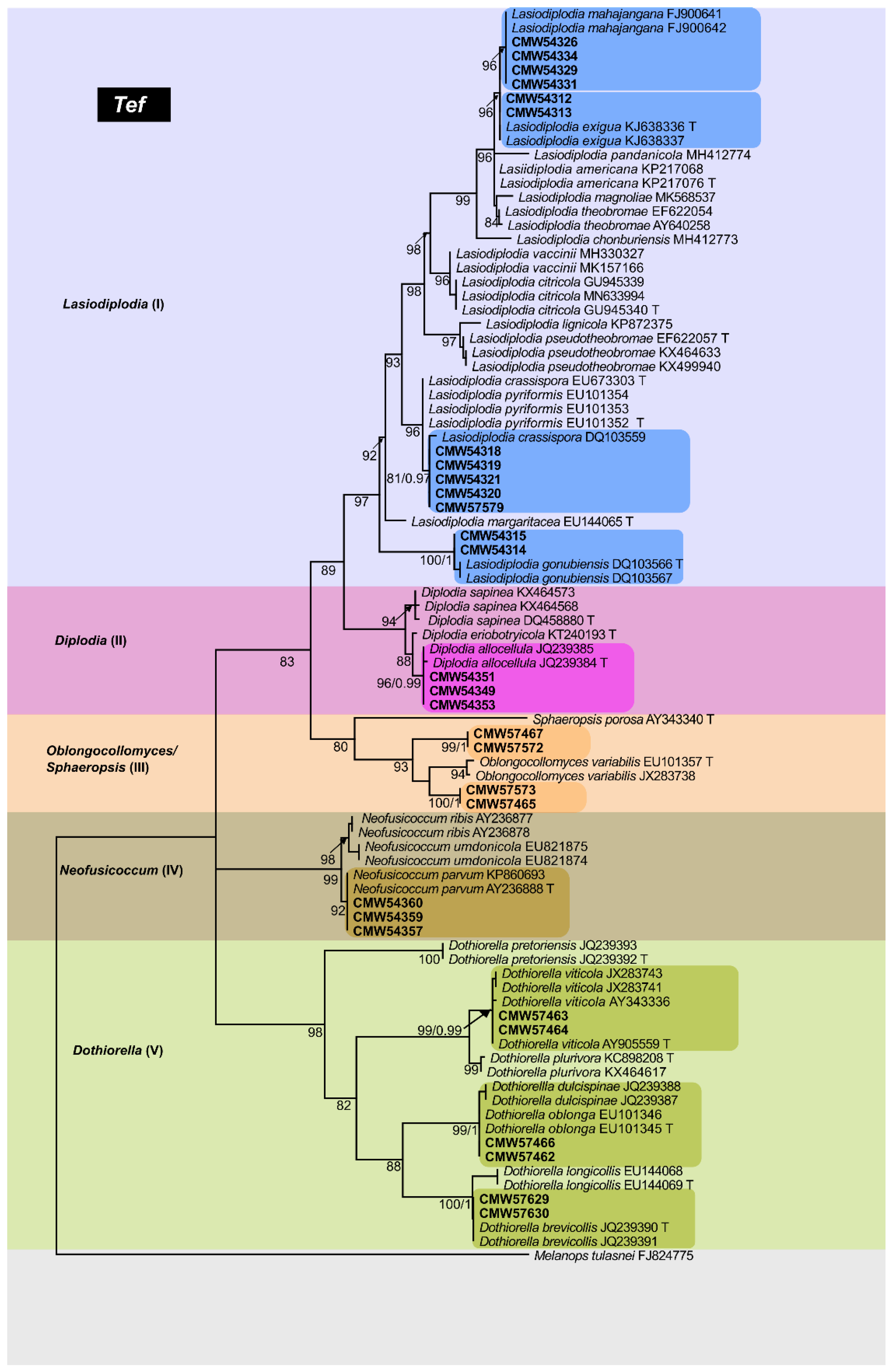
3.3.2. Tef-1α Phylogeny
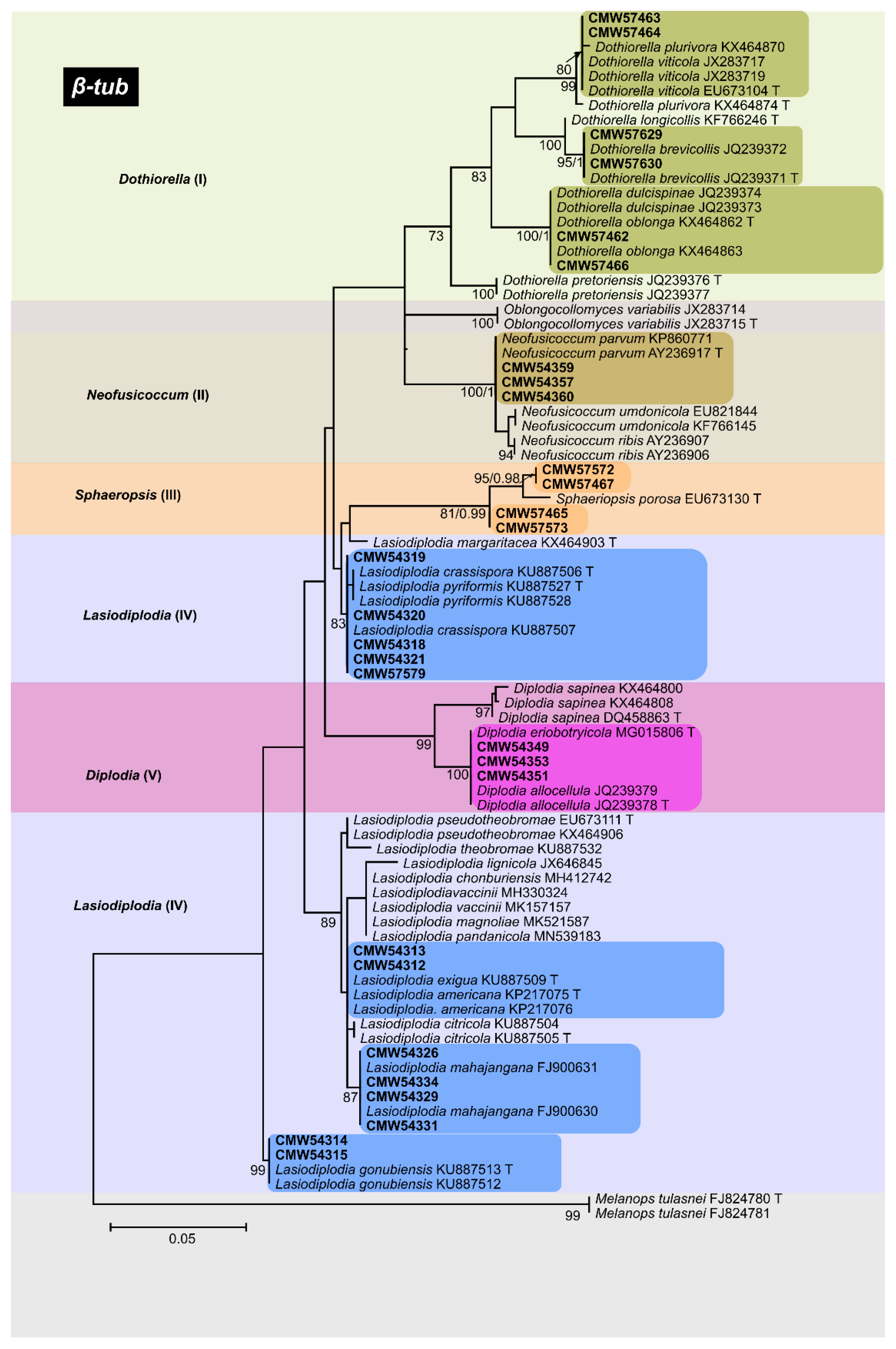
3.3.3. β-Tub Phylogeny
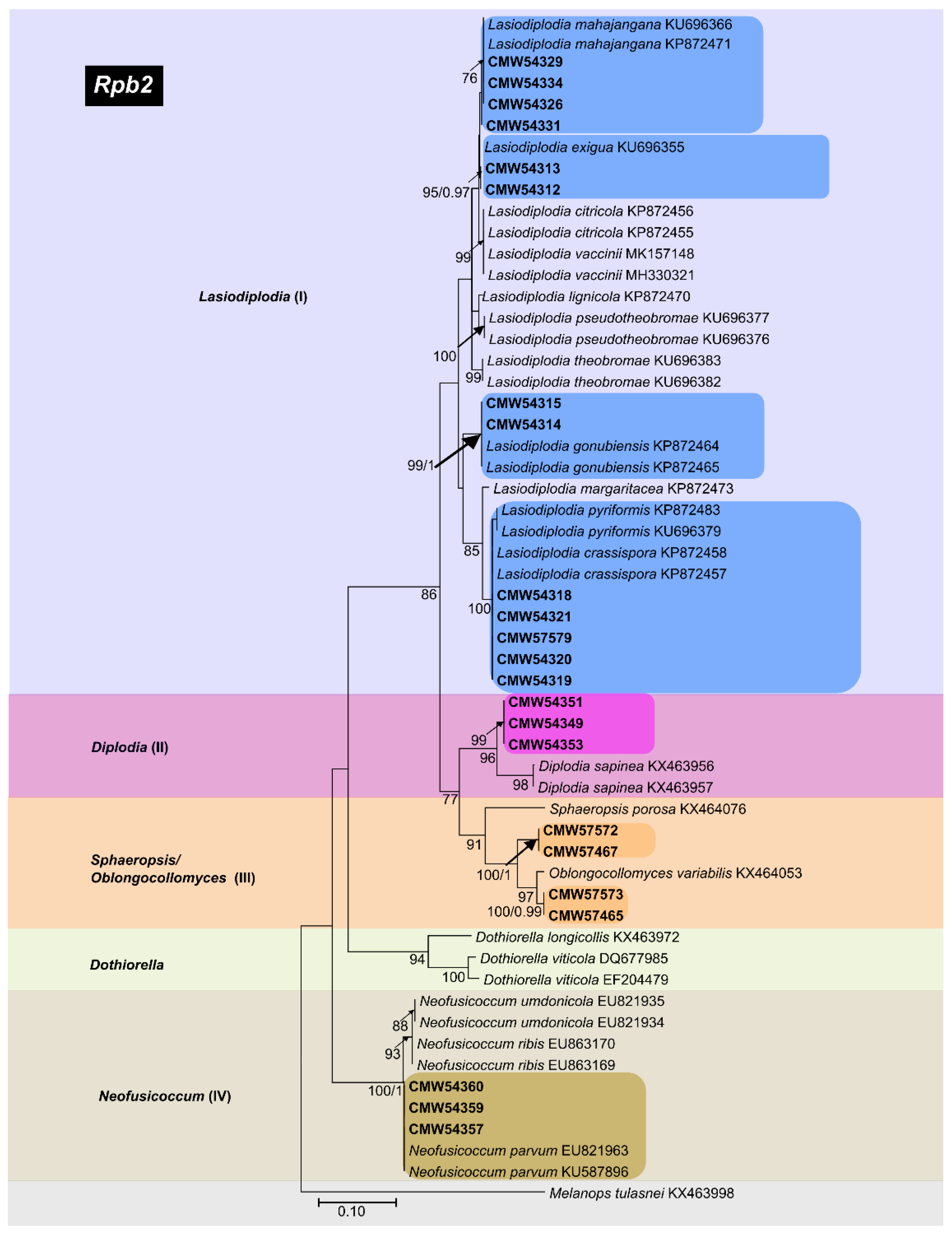
3.3.4. rpb2 Phylogeny
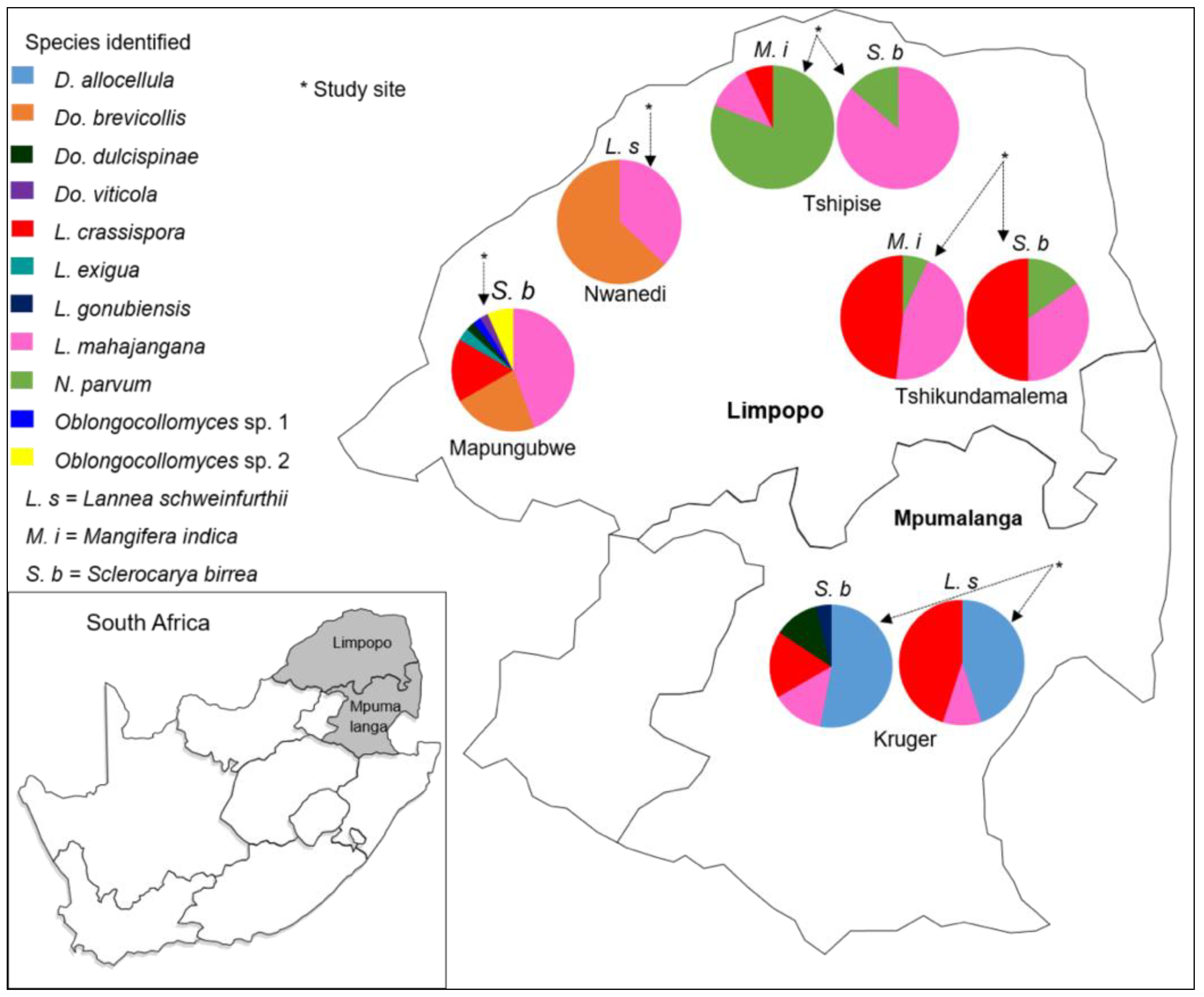
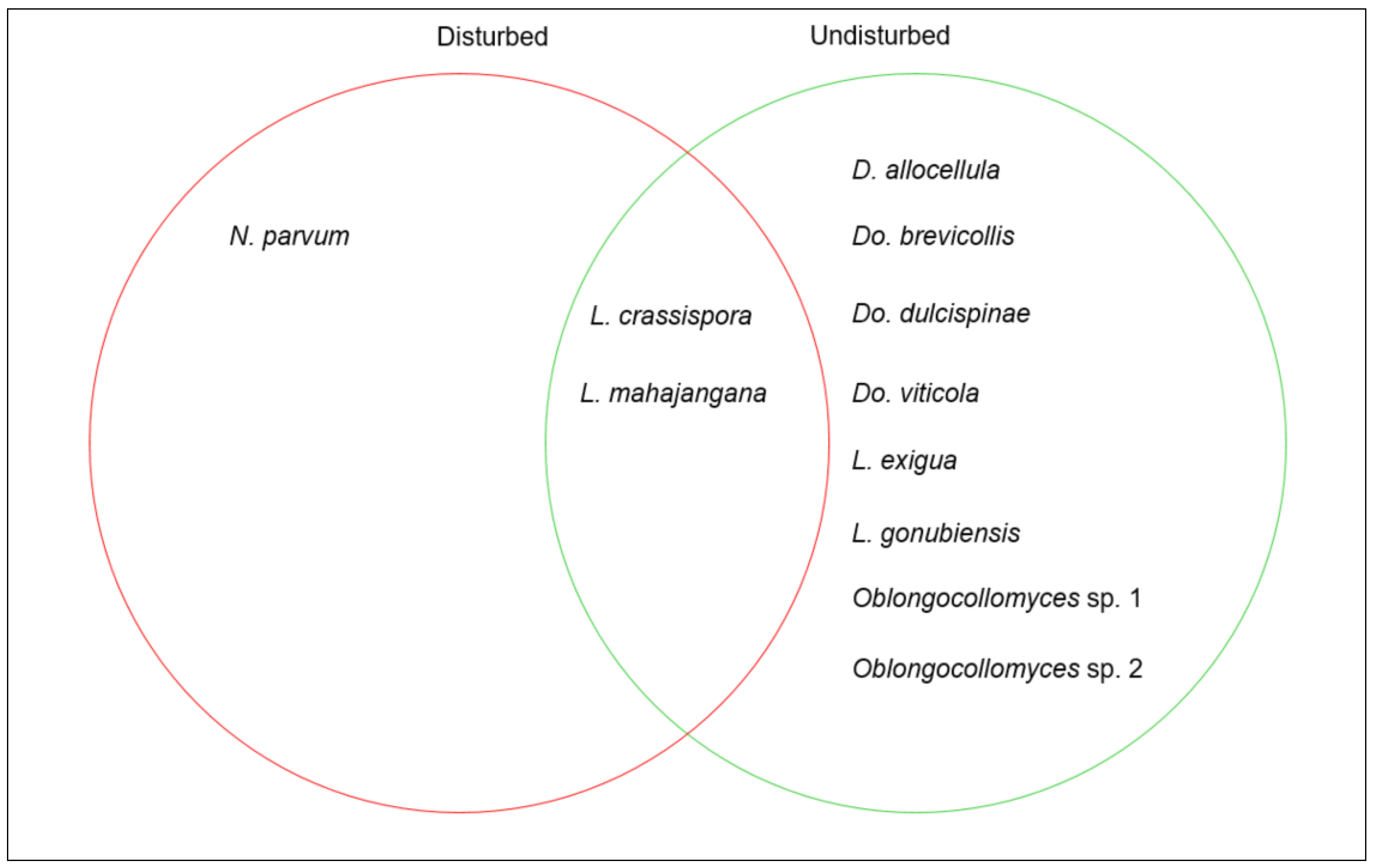
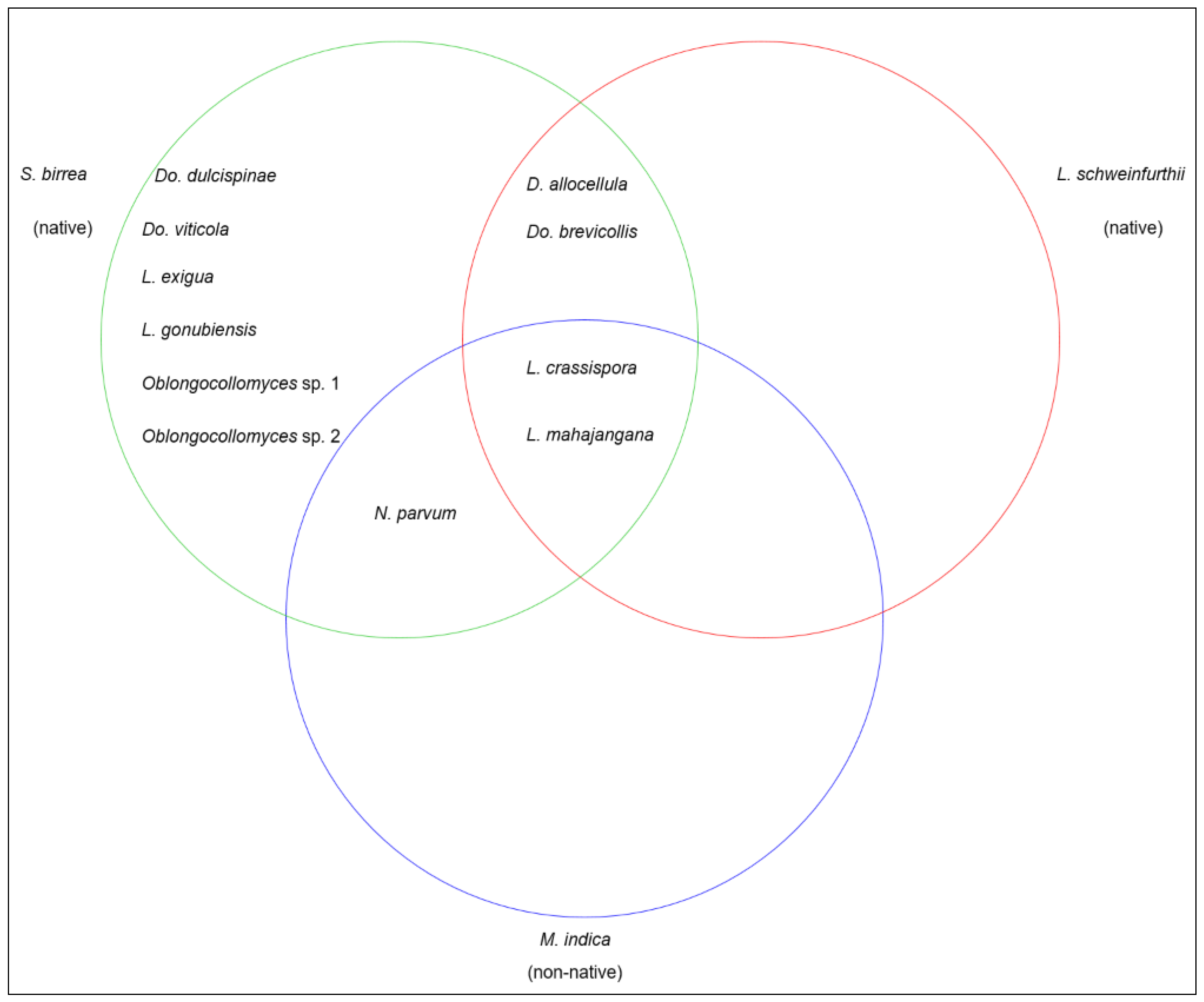
3.4. Species Diversity Distribution
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Van Niekerk, J.M.; Crous, P.W.; Groenewald, J.Z.; Fourie, P.H.; Halleen, F. DNA phylogeny, morphology and pathogenicity of Botryosphaeria species on grapevines. Mycologia 2004, 96, 781–798. [Google Scholar] [CrossRef]
- Damm, U.; Crous, P.W.; Fourie, P.H. Botryosphaeriaceae as potential pathogens of Prunus in South Africa, with descriptions of Diplodia africana and Lasiodiplodia plurivora sp. nov. Mycologia 2007, 99, 664–680. [Google Scholar] [CrossRef]
- Slippers, B.; Smith, W.A.; Crous, P.W.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Taxonomy, phylogeny and identification of Botryosphaeriaceae associated with pome and stone fruit trees in South Africa and other regions of the world. Plant Pathol. 2007, 56, 128–139. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.; Wingfield, M.J.; Crous, P.W.; Coutinho, T.A. Sphaeropsis sapinea and Botryosphaeria dothidea endophytic in Pinus species and Eucalyptus species in South Africa. S. Afr. J. Bot. 1996, 62, 86–88. [Google Scholar] [CrossRef] [Green Version]
- Slippers, B.; Burgess, T.; Pavlic, D.; Ahumada, R.; Maleme, H.; Mohali, S.; Rodas, C.; Wingfield, M.J. A diverse assemblage of Botryosphaeriaceae infect Eucalyptus in native and non-native environments. South. For. 2009, 71, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Pavlic, D.; Slippers, B.; Coutinho, T.A.; Wingfield, M.J. Botryosphaeriaceae occurring on native Syzygium cordatum in South Africa and their potential threat to Eucalyptus. Plant Pathol. 2007, 56, 624–636. [Google Scholar] [CrossRef] [Green Version]
- Van der Linde, J.A.; Six, D.L.; Wingfield, M.J.; Roux, J. Lasiodiplodia species associated with dying Euphorbia ingens in South Africa. South. For. 2011, 73, 165–173. [Google Scholar] [CrossRef]
- Jami, F.; Slippers, B.; Wingfield, M.J.; Gryzenhout, M. Botryosphaeriaceae species overlap on four unrelated, native South African hosts. Fungal Biol. 2014, 118, 168–179. [Google Scholar] [CrossRef] [Green Version]
- Mehl, J.W.M.; Slippers, B.; Roux, J.; Wingfield, M.J. Cankers and other diseases caused by the Botryosphaeriaceae. In Infectious Forest Diseases; Gonthier, P., Nicolotti, G., Eds.; CAB International: Boston, MN, USA, 2013; pp. 298–317. [Google Scholar]
- Burgess, T.I.; Tan, Y.P.; Garnas, J.; Edwards, J.; Scarlett, K.A.; Lucas, A.; Shuttleworth, L.A.; Daniel, R.; Dann, E.K.; Parkinson, L.E.; et al. Current status of the Botryosphaeriaceae in Australia. Australas. Plant Pathol. 2018, 48, 35–44. [Google Scholar] [CrossRef]
- Pillay, K.; Slippers, B.; Wingfield, M.J.; Gryzenhout, M. Diversity and distribution of co-infecting Botryosphaeriaceae from Eucalyptus grandis and Syzygium cordatum in South Africa. S. Afr. J. Bot. 2013, 84, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Mehl, J.W.M.; Slippers, B.; Roux, J.; Wingfield, M.J. Overlap of latent pathogens in the Botryosphaeriaceae on a native and agricultural host. Fungal Biol. 2017, 121, 405–419. [Google Scholar] [CrossRef] [Green Version]
- Slippers, B.; Fourie, G.; Crous, P.W.; Coutinho, T.A.; Wingfield, B.D.; Carnegie, A.J.; Wingfield, M.J. Speciation and distribution of Botryosphaeria species on native and introduced Eucalyptus trees in Australia and South Africa. Stud. Mycol. 2004, 50, 343–358. [Google Scholar]
- Jami, F.; Slippers, B.; Wingfield, M.J.; Gryzenhout, M. Greater Botryosphaeriaceae diversity in healthy than associated diseased Acacia karroo tree tissues. Australas. Plant Pathol. 2013, 42, 421–430. [Google Scholar] [CrossRef]
- Bihon, W.; Burgess, T.I.; Slippers, B.; Wingfield, M.J.; Wingfield, B.D. Distribution of Diplodia pinea and its genotypic diversity within asymptomatic Pinus patula trees. Australas. Plant Pathol. 2011, 40, 540–548. [Google Scholar] [CrossRef]
- Sakalidis, M.L.; Slippers, B.; Wingfield, B.D.; Hardy, G.E.S.J.; Burgess, T.I. The challenge of understanding the origin, pathways and extent of fungal invasions: Global populations of the Neofusicoccum parvum/N. ribis species complex. Divers. Dist. 2013, 19, 873–883. [Google Scholar] [CrossRef]
- Marsberg, A.; Kemler, M.; Jami, F.; Nagel, J.H.; Postma-Smidt, A.; Naidoo, S.; Wingfield, M.J.; Crous, P.W.; Spatafora, J.W.; Hesse, C.N. Botryosphaeria dothidea: A latent pathogen of global importance to woody plant health. Mol. Plant Pathol. 2017, 18, 477–488. [Google Scholar] [CrossRef]
- Mehl, J.; Wingfield, M.J.; Roux, J.; Slippers, B. Invasive everywhere? Phylogeographic analysis of the globally distributed tree pathogen Lasiodiplodia theobromae. Forests 2017, 8, 145. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.Y.; Xie, Y.; Zhang, W.; Wang, Y.; Liu, J.K.; Hyde, K.D.; Seem, R.C.; Zhang, G.Z.; Wang, Z.Y.; Yao, S.W.; et al. Species of Botryosphaeriaceae involved in grapevine dieback in China. Fungal Divers. 2013, 61, 221–236. [Google Scholar] [CrossRef]
- Jami, F.; Wingfield, M.J.; Gryzenhout, M.; Slippers, B. Diversity of tree infecting Botryosphaeriales on native and non-native trees in South Africa and Namibia. Australas. Plant Pathol. 2017, 46, 529–545. [Google Scholar] [CrossRef] [Green Version]
- Trakunyingcharoen, T.; Cheewangkoon, R.; To-anun, C.; Crous, P.W.; Van Niekerk, J.M.; Lombard, L. Botryosphaeriaceae associated with diseases of mango (Mangifera indica). Australas. Plant Pathol. 2014, 43, 425–438. [Google Scholar] [CrossRef]
- Christenhusz, M.J.M.; Byng, J.W. The number of known plants species in the world and its annual increase. Phytotaxa 2016, 261, 201–221. [Google Scholar] [CrossRef] [Green Version]
- Kudi, A.C.; Umoh, J.U.; Eduvie, L.O.; Gefu, J. Screening of some Nigerian medicinal plants for antibacterial activity. J. Ethnopharmacol. 1999, 67, 225–228. [Google Scholar] [CrossRef]
- Shackleton, S.E.; Shackleton, C.M.; Cunningham, A.B.; Lombard, C.; Sullivan, C.A.; Netshiluvhi, T.R. Knowledge on Sclerocarya birrea subsp. caffra with emphasis on its importance as a non-timber forest product in South and southern Africa: A summary. Part 1: Taxonomy, ecology and role in rural livelihoods. South. Afr. For. J. 2002, 194, 27–41. [Google Scholar]
- Rayner, R.W. A mycological colour chart. Brit. Mycol Soc. 1970, 1, 1–34. [Google Scholar]
- Möller, E.M.; Bahnweg, G.; Sandermann, H.; Geiger, H.H. A simple and efficient protocol for isolation of high molecular weight DNA from filamentous fungi, fruit bodies, and infected plant tissues. Nucleic Acids Res. 1992, 20, 6115–6116. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Cruywagen, E.M.; Slippers, B.; Roux, J.; Wingfield, M.J. Phylogenetic Species Recognition and hybridisation in Lasiodiplodia: A case study on species from baobabs. Fungal Biol. 2017, 121, 420–436. [Google Scholar] [CrossRef] [Green Version]
- Sakalidis, M.L.; Hardy, G.E.S.J.; Burgess, T.i. Use of the Genealogical Sorting Index (GSI) to delineate species boundaries in the Neofusicoccum parvum–Neofusicoccum ribis species complex. Mol. Phylogenet. Evol. 2011, 60, 333–344. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Briefings. Bioinf. 2017, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posada, D. JModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Gálvez, E.; Guerrero, P.; Barradas, C.; Crous, P.W.; Alves, A. Phylogeny and pathogenicity of Lasiodiplodia species associated with dieback of mango in Peru. Fungal Biol. 2017, 121, 452–465. [Google Scholar] [CrossRef]
- Zhang, W.; Groenewald, J.Z.; Lombard, L.; Schumaker, R.K.; Phillips, A.J.L.; Crous, P.W. Evaluating species of Botryosphaeriales. Persoonia 2021, 46, 63–115. [Google Scholar] [CrossRef]
- Opik, M.; Moora, M.; Liira, J.; Zobel, M. Composition of root-colonising arbuscular mycorrhizal fungal communities in different ecosystems around the globe. J. Ecol. 2006, 94, 778–790. [Google Scholar] [CrossRef]
- Pavlic, D.; Wingfield, M.J.; Boissin, E.; Slippers, B. The distribution of genetic diversity in the Neofusicoccum parvum/N. ribis complex suggests structure correlated with level of disturbance. Fungal Ecol. 2015, 13, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.E.; Gelembiuk, G.W. Evolutionary origins of invasive populations. Evol. Appl. 2008, 1, 427–448. [Google Scholar] [CrossRef]
- Mehl, J.W.M.; Slippers, B.; Roux, J.; Wingfield, M.J. Botryosphaeriaceae associated with Pterocarpus angolensis (kiaat) in South Africa. Mycologia 2011, 103, 534–553. [Google Scholar] [CrossRef] [Green Version]
- Linaldeddu, B.T.; Deidda, A.; Scanu, B.; Franceschini, A.; Serra, S.; Berraf-Tebbal, A.; Boutiti, M.Z.; Jamâa, M.B.; Phillips, A.J. Diversity of Botryosphaeriaceae species associated with grapevine and other woody hosts in Italy, Algeria and Tunisia, with descriptions of Lasiodiplodia exigua and Lasiodiplodia Mediterranea sp. nov. Fungal Divers. 2015, 71, 201–214. [Google Scholar] [CrossRef]
- Akgül, D.S.; Savaş, N.G.; Özarslandan, M. First report of wood canker caused by Lasiodiplodia exigua and Neoscytalidium novaehollandiae on grapevine in Turkey. Plant Dis. 2019, 103, 1036. [Google Scholar] [CrossRef]
- Montoya, E.A.; Paolinelli, M.; Rolshausen, P.E.; Valenzuela-Solano, C.; Martinez, R. Characterization of Lasiodiplodia species associated with grapevines in Mexico. Phytopathol. Mediterr. 2021, 60, 237–251. [Google Scholar] [CrossRef]
- Jami, F.; Slippers, B.; Wingfield, M.J.; Gryzenhout, M. Five new species of the Botryosphaeriaceae from Acacia karroo in South Africa. Cryptogam. Mycol. 2012, 33, 245–266. [Google Scholar] [CrossRef]
- Luque, J.; Martos, S.; Phillips, A.J.L. Botryosphaeria viticola sp. nov. on grapevines: A new species with a Dothiorella anamorph. Mycologia 2005, 97, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Marques, M.W.; Lima, N.B.; De Morais, A.M.; Barbosa, M.A.G.; Souza, B.O.; Michereff, S.J.; Phillips, A.J.L.; Câmara, M.P.S. Species of Lasiodiplodia associated with mango in Brazil. Fungal Divers. 2013, 61, 181–193. [Google Scholar] [CrossRef]
- Ismail, A.M.; Cirvilleri, G.; Polizzi, G.; Crous, P.W.; Groenewald, J.Z.; Lombard, L. Lasiodiplodia species associated with dieback disease of mango (Mangifera indica) in Egypt. Australas. Plant Pathol. 2012, 41, 649–660. [Google Scholar] [CrossRef]
- Ni, H.F.; Yang, H.R.; Chen, R.S.; Hung, T.H.; Liou, R.F. A nested multiplex PCR for species-specific identification and detection of Botryosphaeriaceae species on mango. Eur. J. Plant Pathol. 2012, 133, 819–828. [Google Scholar] [CrossRef]
- Yang, T.; Groenewald, J.Z.; Cheewangkoon, R.; Jami, F.; Abdollahzadeh, J.; Lombard, L.; Crous, P.W. Families, genera and species of Botryosphaeriales. Fungal Biol. 2016, 121, 322–346. [Google Scholar] [CrossRef]
- Iturritxa, E.; Slippers, B.; Mesanza, N.; Wingfield, M.J. First report of Neofusicoccum parvum causing canker and die-back of Eucalyptus in Spain. Australas. Plant Dis. 2011, 6, 57–59. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate No. | Identity | Host | Location | GenBank | |||
|---|---|---|---|---|---|---|---|
| ITS | tef1-α | β-tub | rpb2 | ||||
| CMW54349 | Diplodia allocellula | Lannea schweinfurthii | Nwanedi | OL441853 | OL441909 | OL441965 | OM585579 |
| CMW54351 | D. allocellula | Sclerocarya birrea | Kruger | OL441855 | OL441911 | OL441967 | OM585581 |
| CMW54353 | D. allocellula | Lannea schweinfurthii | Kruger | OL441857 | OL441913 | OL441969 | OM585583 |
| CMW57629 | Dothiorella brevicollis | Lannea schweinfurthii | Nwanedi | OL441858 | OL441914 | OL441970 | - |
| CMW57630 | Do. brevicollis | Sclerocarya birrea | Mapungubwe | OL441859 | OL441915 | OL441971 | - |
| CMW57462 | Do. dulcispinae | Sclerocarya birrea | Kruger | OL441861 | OL441917 | OL441973 | - |
| CMW57466 | Do. dulcispinae | Sclerocarya birrea | Mapungubwe | OL441862 | OL441918 | OL441974 | - |
| CMW57463 | Do. viticola | Sclerocarya birrea | Mapungubwe | OL441863 | OL441919 | OL441975 | - |
| CMW57464 | Do. viticola | Sclerocarya birrea | Mapungubwe | OL441864 | OL441920 | OL441976 | - |
| CMW54318 | Lasiodiplodia crassispora | Lannea schweinfurthii | Kruger | OL441866 | OL441922 | OL441978 | OL442021 |
| CMW54319 | L. crassispora | Sclerocarya birrea | Tshikundamalema | OL441867 | OL441923 | OL441979 | OL442022 |
| CMW54320 | L. crassispora | Sclerocarya birrea | Mapungubwe | OL441868 | OL441924 | OL441980 | OL442023 |
| CMW54321 | L. crassispora | Mangifera indica | Tshipise | OL441869 | OL441925 | OL441981 | OL442024 |
| CMW57579 | L. crassispora | Sclerocarya birrea | Kruger | OL441870 | OL441926 | OL441982 | OL442025 |
| CMW54314 | L. gonubiensis | Sclerocarya birrea | Kruger | OL441873 | OL441929 | OL441985 | OL442028 |
| CMW54315 | L. gonubiensis | Sclerocarya birrea | Kruger | OL441874 | OL441930 | OL441986 | OL442029 |
| CMW54312 | L. exigua | Sclerocarya birrea | Mapungubwe | OL441875 | OL441931 | OL441987 | OL442030 |
| CMW54313 | L. exigua | Sclerocarya birrea | Mapungubwe | OL441876 | OL441932 | OL441988 | OL442031 |
| CMW54326 | L. mahajangana | Mangifera indica | Tshikundamalema | OL441877 | OL441933 | OL441989 | OL442032 |
| CMW54329 | L. mahajangana | Lannea schweinfurthii | Nwanedi | OL441878 | OL441934 | OL441990 | OL442033 |
| CMW54331 | L. mahajangana | Sclerocarya birrea | Mapungubwe | OL441880 | OL441936 | OL441992 | OL442035 |
| CMW54334 | L. mahajangana | Sclerocarya birrea | Kruger | OL441881 | OL441937 | OL441993 | OL442036 |
| CMW54357 | Neofusicoccum parvum | Sclerocarya birrea | Tshipise | OL441892 | OL441948 | OL442004 | OL442047 |
| CMW54359 | N. parvum | Mangifera indica | Tshipise | OL441893 | OL441949 | OL442005 | OL442048 |
| CMW54360 | N. parvum | Mangifera indica | Tshikundamalema | OL441894 | OL441950 | OL442006 | OL442049 |
| CMW57467 | Oblongocollomyces sp. 1 | Sclerocarya birrea | Mapungubwe | OL441905 | OL441961 | OL442017 | OL442060 |
| CMW57572 | Oblongocollomyces sp. 1 | Sclerocarya birrea | Mapungubwe | OL441906 | OL441962 | OL442018 | OL442061 |
| CMW57465 | Oblongocollomyces sp. 2 | Sclerocarya birrea | Mapungubwe | OL441907 | OL441963 | OL442019 | OL442062 |
| CMW57573 | Oblongocollomyces sp. 2 | Sclerocarya birrea | Mapungubwe | OL441908 | OL441964 | OL442020 | OL442063 |
| Health Status | S. birrea | M. indica | L. schweinfurthii | Total |
|---|---|---|---|---|
| 0 = healthy | 6 | 0 | 0 | 6 |
| 1 = <25% dieback branches | 117 | 17 | 81 | 215 |
| 2 = 25–50% dieback branches | 52 | 55 | 34 | 141 |
| 3 = >50% dieback branches | 12 | 12 | 9 | 33 |
| 4 = appearing to be dying or nearly dead | 4 | 0 | 5 | 9 |
| Total | 191 | 84 | 129 | 404 |
| Healthy | <25% Dieback Branches | 25–50% Dieback Branches | >50% Dieback Branches | Appearing to Be Dying or Nearly Dead | Total | |
|---|---|---|---|---|---|---|
| Tshikundamalema | 0 | 51 | 42 | 10 | 0 | 103 |
| Tshipise | 0 | 24 | 24 | 2 | 0 | 50 |
| Nwanedi | 0 | 28 | 2 | 0 | 0 | 30 |
| Mapungubwe | 6 | 11 | 26 | 7 | 4 | 54 |
| Kruger | 0 | 101 | 47 | 14 | 5 | 167 |
| Species | Country | Strain Number | GenBank | |||
|---|---|---|---|---|---|---|
| ITS | tef1-α | rpb2 | β-tub | |||
| Diplodia allocellula | South Africa | CBS 130408 CMW 36468 | JQ239397 | JQ239384 | None | JQ239378 |
| D. allocellula | South Africa | CMW 36469 | NR_111701 | JQ239385 | None | JQ239379 |
| D. eriobotryicola | Spain | CBS 140851 | NR_152462 | KT240193 | None | MG015806 |
| D. sapinea | Netherlands | CBS 393.84 | DQ458895 | DQ458880 | None | DQ458863 |
| D. sapinea | South Africa | CBS 109726 | KX464094 | KX464568 | KX463956 | KX464800 |
| D. sapinea | Australia | CBS 189.37 | KX464099 | KX464573 | KX463957 | KX464808 |
| Dothiorella brevicollis | South Africa | CMW 36464 | JQ239404 | JQ239391 | None | JQ239372 |
| Do. brevicollis | South Africa | CBS 130411 CMW 36463 | NR_111703 | JQ239390 | None | JQ239371 |
| Do. dulcispinae | South Africa | CMW 36461 | JQ239401 | JQ239388 | None | JQ239374 |
| Do. dulcispinae | Namibia | CMW 36460 | NR_111702 | JQ239387 | None | JQ239373 |
| Do. longicollis | Australia | CBS 122068 CMW 26166 | NR_136999 | EU144069 | KX463972 | KF766246 |
| Do. longicollis | Australia | CMW 26165 | EU144053 | EU144068 | None | None |
| Do. oblonga | South Africa | CBS 121765 CMW 25407 | EU101301 | EU101345 | None | KX464862 |
| Do. oblonga | South Africa | CBS 121766 CMW 25408 | NR_137689 | EU101346 | None | KX464863 |
| Do. plurivora | Iran | CBS 124724 IRAN 1557C | KC898225 | KC898208 | None | KX464874 |
| Do. plurivora | USA, California | CBS 120999 | KX464125 | KX464617 | None | KX464870 |
| Do. pretoriensis | South Africa | CBS 130404 CMW 36480 | JQ239405 | JQ239392 | None | JQ239376 |
| Do. pretoriensis | South Africa | CMW 36481 | JQ239406 | JQ239393 | None | JQ239377 |
| Do. viticola | South Africa | CMW 37928 | JX283730 | JX283741 | None | JX283717 |
| Do. viticola | South Africa | CMW 37933 | JX283735 | JX283743 | None | JX283719 |
| Do. viticola | Spain | CBS 117009 | MH863011 | AY905559 | DQ677985 | EU673104 |
| Do. viticola | South Africa | STE-U5048 | AY343373 | AY343336 | EF204479 | None |
| L. americana | USA, Arizona | CERC 1962 CFCC 50066 | KP217060 | KP217068 | None | KP217076 |
| L. americana | USA, Arizona | CERC 1961 CFCC 50065 | KP217059 | KP217067 | None | KP217075 |
| Lasiodiplodia chonburiensis | Thailand | MFLUCC 16-0376 | MH275066 | MH412773 | None | MH412742 |
| L. citricola | Iran | IRNKB3 | MN634040 | MN633994 | None | None |
| L. citricola | Iran | CBS 124707 IRAN 1522C | GU945354 | GU945340 | KP872455 | KU887505 |
| L. citricola | Iran | CBS 124706 IRAN 1521C | GU945353 | GU945339 | KP872456 | KU887504 |
| L. crassispora | Venezuela | CMW 13488 | DQ103552 | DQ103559 | KP872458 | KU887507 |
| L. crassispora | Australia | CBS 118741 WAC 12533 | NR_111194 | EU673303 | KP872457 | KU887506 |
| L. exigua | Tunisia | CBS 137785 BL104 | KJ638317 | KJ638336 | KU696355 | KU887509 |
| L. exigua | Tunisia | BL184 | KJ638318 | KJ638337 | None | None |
| L. gonubiensis | South Africa | CBS 115812 CMW 14077 | DQ458892 | DQ103566 | KP872464 | KU887512 |
| L. gonubiensis | South Africa | CBS 116355 CMW 14078 | AY639594 | DQ103567 | KP872465 | KU887513 |
| L. lignicola | Thailand | MFLUCC 11-0435 | JX646797 | KP872375 | KP872470 | JX646845 |
| L. lignicola | India | SUF161 | MT081525 | None | None | None |
| L. magnoliae | China | MFLUCC 18-0948 | MK499387 | MK568537 | None | MK521587 |
| L. mahajangana | Madagascar | CMW 27801 | FJ900595 | FJ900641 | KP872471 | FJ900630 |
| L. mahajangana | Madagascar | CMW 27818 | FJ900596 | FJ900642 | KU696366 | FJ900631 |
| L. margaritacea | Australia | CBS 122519 CMW 26162 | NR_136998 | EU144065 | KP872473 | KX464903 |
| L. pandanicola | Thailand | MFLUCC 16-0265 | MH275068 | MH412774 | None | None |
| L. pandanicola | China | GBLZ16BO-008 | MN540679 | None | None | MN539183 |
| L. pseudotheobromae | Costa Rica | CBS 116459 CMW 40939 | EF622077 | EF622057 | KU696376 | EU673111 |
| L. pseudotheobromae | Zaire | CBS 374.54 | KX464139 | KX464633 | None | KX464906 |
| L. pseudotheobromae | China | BJFU ZYP151106-14 | KX499902 | KX499940 | KX499977 | None |
| L. pyriformis | Namibia | CBS 121771 CMW 25415 | EU101308 | EU101353 | KU696379 | KU887528 |
| L. pyriformis | Namibia | CMW 25416 | EU101309 | EU101354 | None | None |
| L. pyriformis | Namibia | CBS 121770 CMW 25414 | NR_136993 | EU101352 | KP872483 | KU887527 |
| L. theobromae | Papua New Guinea | CBS 164.96 CMW 50942 | AY640255 | AY640258 | KU696383 | KU887532 |
| L. theobromae | unknown | CBS 111530 | EF622074 | EF622054 | KU696382 | None |
| L. vaccinii | China, Beijing | CGMCC 3.19022 | MH330318 | MH330327 | MH330321 | MH330324 |
| L. vaccinii | China, Beijing | CGMCC3.19256 | MK157139 | MK157166 | MK157148 | MK157157 |
| Neofusicoccum parvum | South Africa | CMW 41213 | KP860849 | KP860693 | KU587896 | KP860771 |
| N. parvum | New Zealand | CBS 138823 CMW 9081 | NR_119487 | AY236888 | EU821963 | AY236917 |
| N. ribis | USA, New York | CMW 7772 | AY236935 | AY236877 | EU863170 | AY236906 |
| N. ribis | USA, New York | CMW 7773 | AY236936 | AY236878 | EU863169 | AY236907 |
| N. umdonicola | South Africa | CMW 14058 | EU821904 | EU821874 | EU821934 | EU821844 |
| N. umdonicola | South Africa | CBS 123646 CMW 14060 | EU821905 | EU821875 | EU821935 | KF766145 |
| Oblongocollomyces variabilis | Namibia/RSA | CMW 36482 | JX283726 | JX283738 | None | JX283714 |
| O. variabilis | Namibia | CBS 121774 CMW 25419 | NR_136994 | EU101357 | KX464053 | JX283715 |
| Sphaeropsis porosa | South Africa | CBS 110496 CPC 5132 | NR_119492 | AY343340 | KX464076 | EU673130 |
| Melanops tulasnei | Germany | CBS 116805 | FJ824769 | None | None | FJ824780 |
| M. tulasnei | Germany | CBS 116806 | FJ824770 | FJ824775 | KX463998 | FJ824781 |
| Location | Host | Identity | Number of Isolates |
|---|---|---|---|
| Tshikundamalema | Sclerocarya birrea | Lasiodiplodia crassispora | 10 |
| Lasiodiplodia mahajangana | 7 | ||
| Neofusicoccum parvum | 3 | ||
| Tshikundamalema | Mangifera indica | Lasiodiplodia crassispora | 28 |
| Lasiodiplodia mahajangana | 26 | ||
| Neofusicoccum parvum | 4 | ||
| Tshipise | Sclerocarya birrea | Lasiodiplodia mahajangana | 25 |
| Neofusicoccum parvum | 4 | ||
| Tshipise | Mangifera indica | Lasiodiplodia crassispora | 3 |
| Lasiodiplodia mahajangana | 5 | ||
| Neofusicoccum parvum | 34 | ||
| Kruger | Lannea schweinfurthii | Diplodia allocellula | 25 |
| Lasiodiplodia crassispora | 27 | ||
| Lasiodiplodia mahajangana | 6 | ||
| Kruger | Sclerocarya birrea | Diplodia allocellula | 29 |
| Dothiorella dulcispinae | 6 | ||
| Lasiodiplodia crassispora | 9 | ||
| Lasiodiplodia gonubiensis | 2 | ||
| Lasiodiplodia mahajangana | 7 | ||
| Nwanedi | Lannea schweinfurthii | Dothiorella brevicollis | 34 |
| Lasiodiplodia mahajangana | 20 | ||
| Mapungubwe | Sclerocarya birrea | Dothiorella brevicollis | 20 |
| Dothiorella dulcispinae | 2 | ||
| Dothiorella viticola | 6 | ||
| Lasiodiplodia crassispora | 15 | ||
| Lasiodiplodia exigua | 3 | ||
| Lasiodiplodia mahajangana | 40 | ||
| Oblongocollomyces sp. 1 | 2 | ||
| Oblongocollomyces sp. 2 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramabulana, E.; Kunjeku, E.; Slippers, B.; Coetzee, M.P.A. Diversity of Endophytes in the Botryosphaeriaceae Differs on Anacardiaceae in Disturbed and Undisturbed Ecosystems in South Africa. Forests 2022, 13, 341. https://doi.org/10.3390/f13020341
Ramabulana E, Kunjeku E, Slippers B, Coetzee MPA. Diversity of Endophytes in the Botryosphaeriaceae Differs on Anacardiaceae in Disturbed and Undisturbed Ecosystems in South Africa. Forests. 2022; 13(2):341. https://doi.org/10.3390/f13020341
Chicago/Turabian StyleRamabulana, Elelwani, Ednah Kunjeku, Bernard Slippers, and Martin P. A. Coetzee. 2022. "Diversity of Endophytes in the Botryosphaeriaceae Differs on Anacardiaceae in Disturbed and Undisturbed Ecosystems in South Africa" Forests 13, no. 2: 341. https://doi.org/10.3390/f13020341
APA StyleRamabulana, E., Kunjeku, E., Slippers, B., & Coetzee, M. P. A. (2022). Diversity of Endophytes in the Botryosphaeriaceae Differs on Anacardiaceae in Disturbed and Undisturbed Ecosystems in South Africa. Forests, 13(2), 341. https://doi.org/10.3390/f13020341