
A Structural Assessment of Sycamore Maple Bark Disintegration by Nectria cinnabarina

 ,
,  ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant and Fungal Materials
2.2. Light Microscopy
2.3. Fluorescence and Confocal Laser Scanning Microscopy
2.4. Scanning Electron Microscopy
2.5. X-ray Micro-Computed Tomography
2.6. Statistical Analysis
3. Results
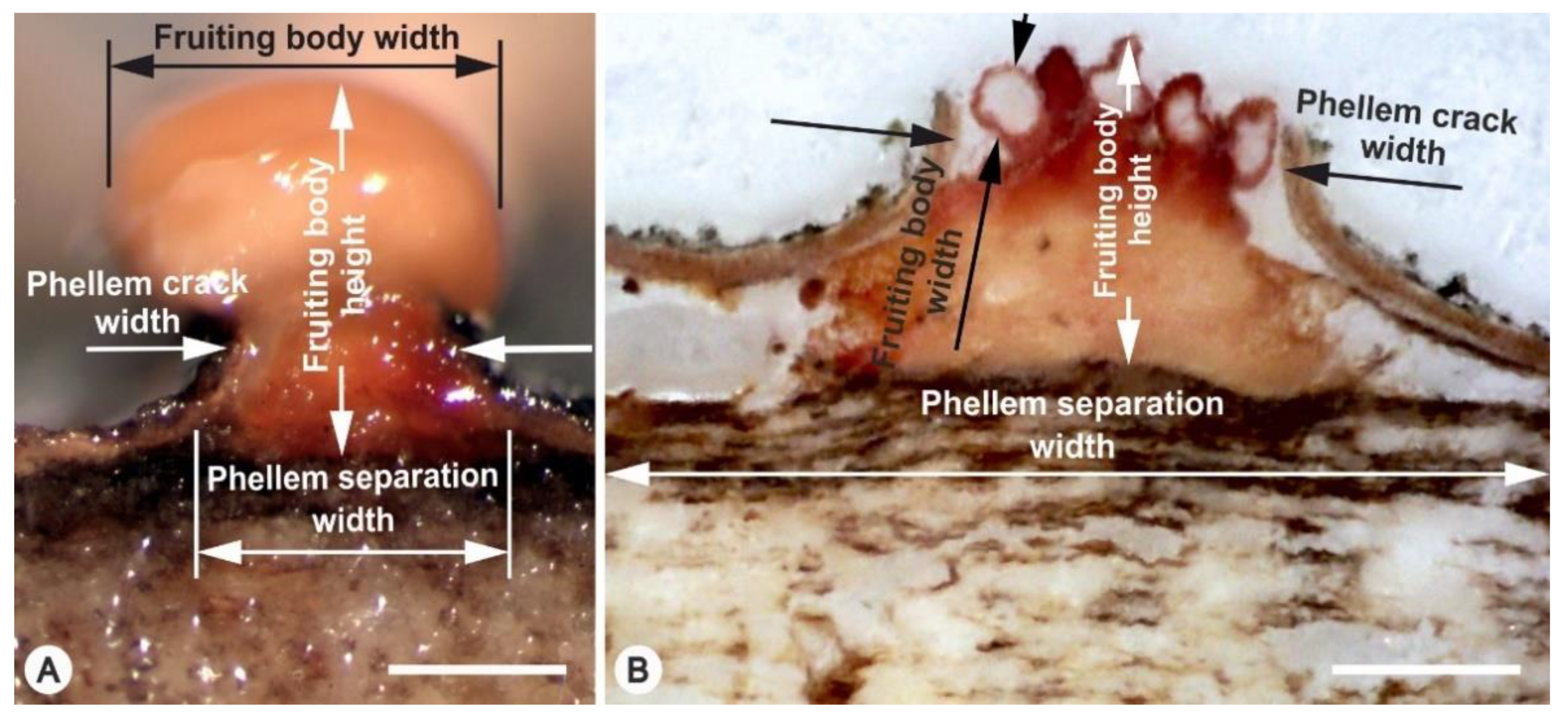
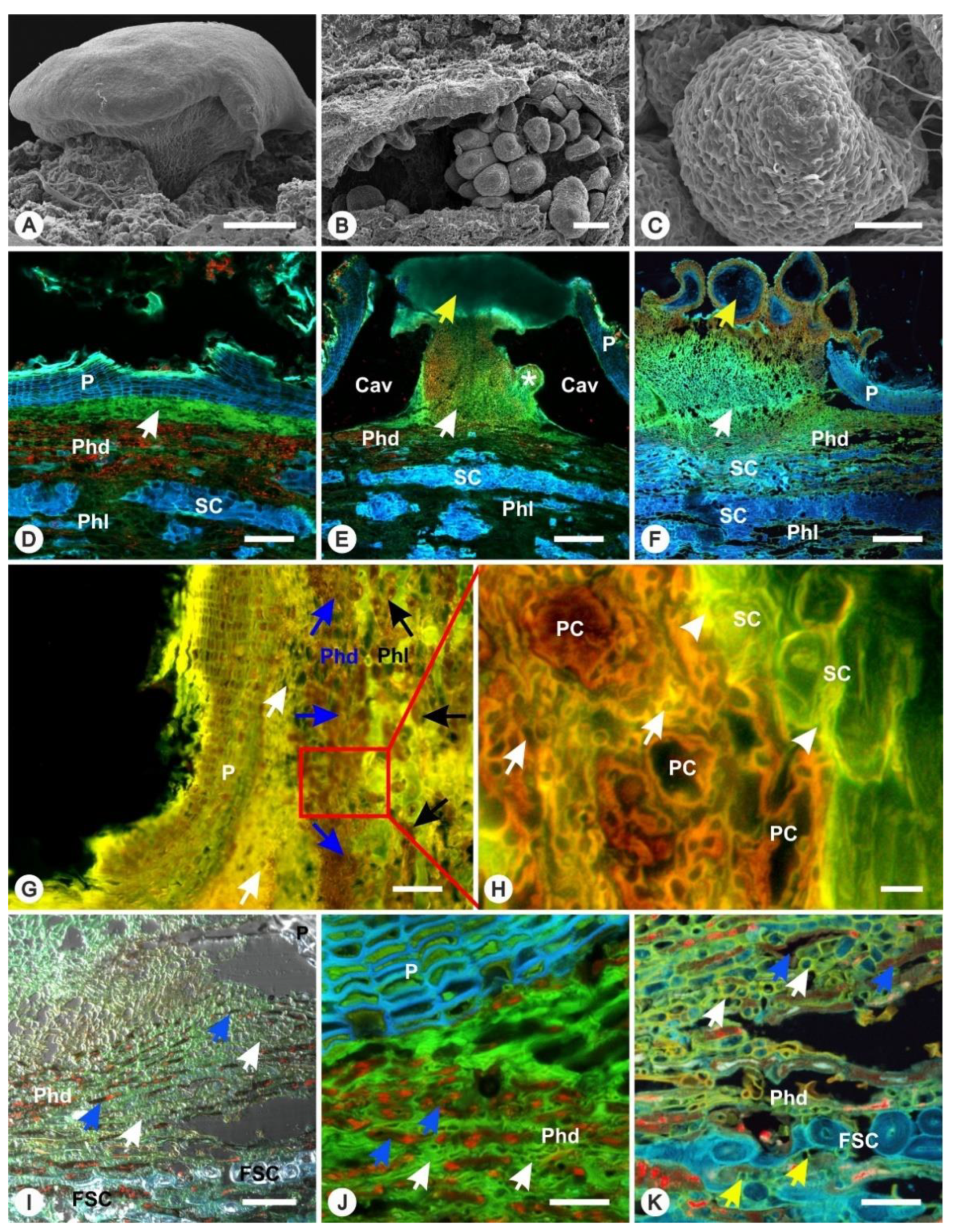
3.1. Formation of FBs
3.2. Spread of Infection Inside the Periderm
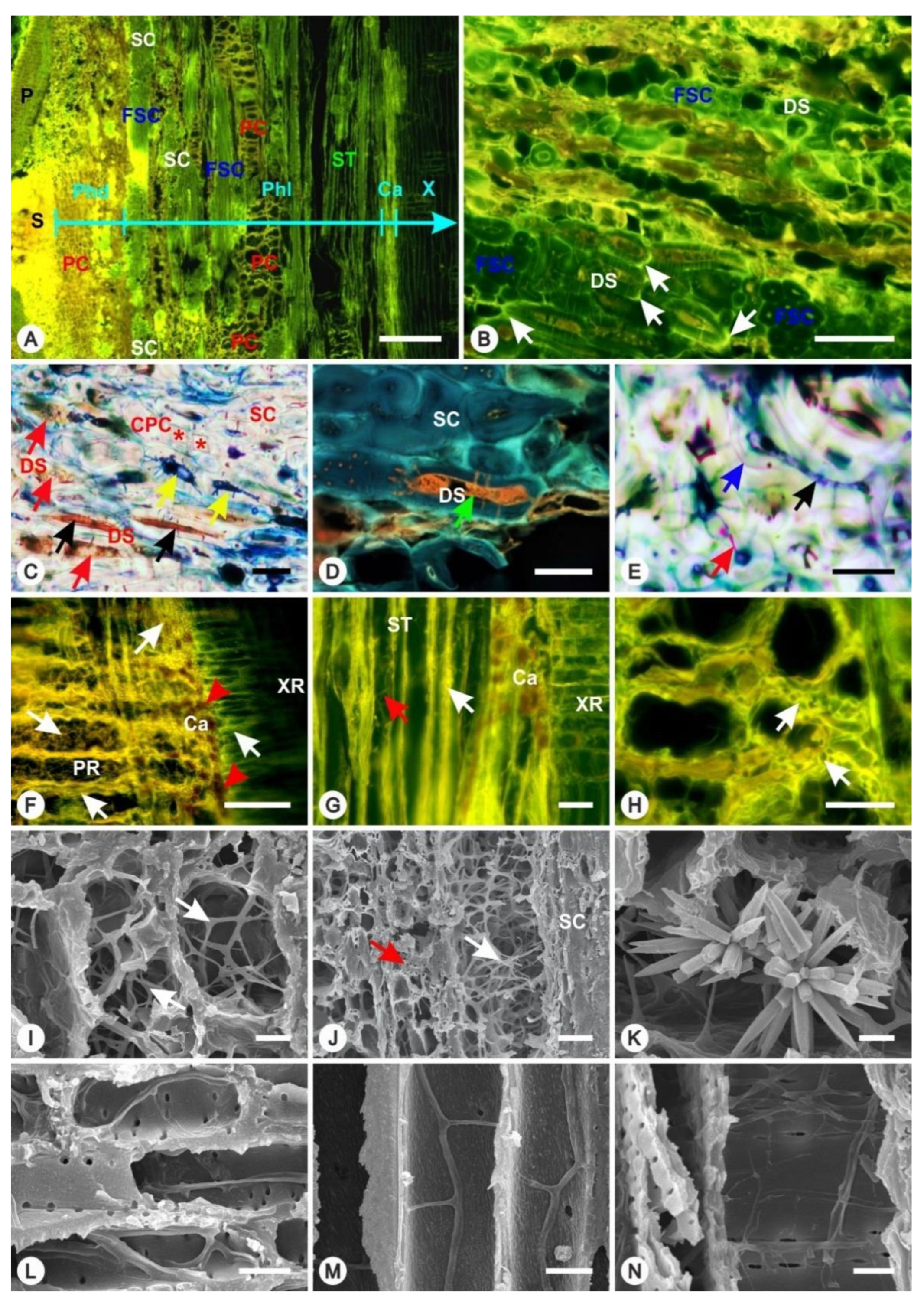
3.3. Disintegration of the Periderm Layer
3.4. Spread of Infection Inside Phloem Tissues
3.5. Spread of Infection Inside Xylem Tissues
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hirooka, Y.; Rossman, A.Y.; Chaverri, P. A morphological and phylogenetic revision of the Nectria cinnabarina species complex. Stud. Mycol. 2011, 68, 35–56. [Google Scholar] [CrossRef]
- Mihál, I. Occurrence of fungi of the genus Nectria s.l. (Ascomycota, Hypocreales, Bionectriaceae, Nectriaceae) in Slovakia. Folia Oecol. 2011, 38, 80–88. [Google Scholar]
- Rossman, A.Y.; Seifert, K.A.; Samuels, G.J.; Minnis, A.M.; Schroers, H.J.; Lombard, L.; Crous, P.W.; Põldmaa, K.; Cannon, P.F.; Summerbell, R.C.; et al. Genera in Bionectriaceae, Hypocreaceae, and Nectriaceae (Hypocreales) proposed for acceptance or rejection. IMA Fungus 2013, 4, 41–51. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Ainsworth & Bisby’s Dictionary of Fungi, 10th ed.; CAB International: Wallingford, UK, 2008; 771p. [Google Scholar]
- Mitchell, C.P.; Ford-Robertson, J.B.; Hinckley, T.; Sennerby-Forsse, L. Ecophysiology of Short Rotation Forest Crops; Kluwer Academic Publishers: Dordrecht, the Netherlands, 1993; 308p. [Google Scholar]
- Jones, R.K.; Benson, D.M. Diseases of Woody Ornamentals and Trees in Nurseries; American Phytopathological Society: St. Paul, MN, USA, 2001; 482p. [Google Scholar]
- Uri, J. The parasitism of Nectria cinnabarina (Tode) Fr. Tijdschr. Planteziekt. 1948, 54, 29–73. [Google Scholar]
- Sinclair, W.A.; Lyon, H.H. Diseases of Trees and Shrubs, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 2005; 660p. [Google Scholar]
- Mayr, H. Über den Parasitismus von Nectria cinnabarina; Springer: Berlin, Germany, 1882. [Google Scholar]
- Račko, V.; Mihál, I.; Mišíková, O. Beech bark disease in Slovakia related to fungi of the genus Nectria s.l. and the anatomy of necrotised bark and wood: A brief review. Folia Oecol. 2020, 47, 16–22. [Google Scholar] [CrossRef]
- Jorgensen, H.A. Studies on Nectria cinnabarina–hosts and variations. Årsskr. K. Vet. Landbohøjsk. 1952, 35, 57–120. [Google Scholar]
- Bedker, P.J.; Blanchette, R.A. Development of cankers caused by Nectria cinnabarina on honey locusts after root pruning. Plant Dis. 1983, 67, 1010–1013. [Google Scholar] [CrossRef]
- Bedker, P.J.; Blanchette, R.A.; French, D.W. Nectria cinnabarina: The cause of a canker disease of honey locust in Minnesota. Plant Dis. 1982, 66, 1067–1070. [Google Scholar] [CrossRef]
- Rossman, A.Y.; Samuels, G.J.; Rogerson, C.T.; Lowen, R. Genera of Bionectriaceae, Hypocreaceae and Nectriaceae (Hypocreales, Ascomycetes). Stud. Mycol. 1999, 42, 1–248. [Google Scholar]
- Seifert, K.A. A Monograph of Stilbella and Some Allied Hyphomycetes; Centraalbureau voor Schimmelcultures: Baarn, the Netherlands, 1985; 235p. [Google Scholar]
- Grigaliūnaitė, B.; Burokienė, D.; Matelis, A.; Stackevičienė, E. Cancer diseases on birch (Betula L.) in Vilnius city green plantations. Acta. Biol. Univ. Daugavp. 2010, 2, 133–136. [Google Scholar]
- Van Gelderen, D.M.; De Jong, P.C.; Oterdoom, H.J. Maples of the World; Timber Press: Portland, OR, USA, 1994; 478p. [Google Scholar]
- Hiemstra, J.A.; Harris, D.C. A Compendium of Verticillium Wilt in Tree Species; Ponsen & Looijen: Wageningen, the Netherlands, 1998; 80p. [Google Scholar]
- Milenković, I.; Nowakowska, J.A.; Oszako, T.; Mladenović, K.; Lučić, A.; Rakonjac, L.; Karadžić, D. Morphological and molecular identification of Phytophthora species from maple trees in Serbia. Genetika 2014, 42, 353–368. [Google Scholar] [CrossRef] [Green Version]
- Tkaczyk, M.; Sikora, K.; Kunca, A. First report on the occurrence of Phytophthora obscura on Acer pseudoplatanus in Slovakia. For. Pathol. 2021, 51, e12686. [Google Scholar] [CrossRef]
- Kelnarová, I.; Černý, K.; Zahradník, D.; Koukol, O. Widespread latent infection of Cryptostroma corticale in asymptomatic Acer pseudoplatanus as a risk for urban plantations. For. Pathol. 2017, 47, e12344. [Google Scholar] [CrossRef]
- Yang, D.-Q. Isolation of wood-inhabiting fungi from Canadian hardwood logs. Can. J. Microbiol. 2005, 51, 1–6. [Google Scholar] [CrossRef]
- Hirooka, Y.; Rossman, A.Y.; Samuels, G.J.; Lechat, C.; Chaverri, P. A monograph of Allantonectria, Nectria, and Pleonectria (Nectriaceae, Hypocreales, Ascomycota) and their pycnidial, sporodochial, and synnematous anamorphs. Stud. Mycol. 2012, 71, 1–210. [Google Scholar] [CrossRef] [Green Version]
- Breitenbach, J.; Kränzlin, F. Fungi of Switzerland. Vol. 1. Ascomycetes; Verlag Mykologia: Lucerne, Switzerland, 1986; 412p. [Google Scholar]
- Račko, V.; Mišíková, O.; Štefková, J.; Čunderlík, I. A fast method to prepare microslides of wood in advanced stages of decay. IAWA J. 2018, 39, 234–243. [Google Scholar] [CrossRef]
- O’Brien, T.P.; Feder, N.; McCully, M.E. Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma 1964, 59, 368–373. [Google Scholar] [CrossRef]
- Yeung, E.C.; Chan, C.K.W. The Glycol Methacrylate Embedding Resins—Technovit 7100 and 8100. In Plant Microtechniques and Protocols; Yeung, E., Stasolla, C., Sumner, M., Huang, B., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 67–82. [Google Scholar]
- Karadžić, D.; Stanivuković, Z.; Milanović, S.; Sikora, K.; Radulović, Z.; Račko, V.; Kardošová, M.; Ďurkovič, J.; Milenković, I. Development of Neonectria punicea pathogenic symptoms in juvenile Fraxinus excelsior trees. Front. Plant Sci. 2020, 11, 592260. [Google Scholar] [CrossRef]
- Line, J. The parasitism of Nectria cinnabarina (coral spot), with special reference to its action on red currant. Trans. Br. Mycol. Soc. 1922, 8, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Ghasemkhani, M.; Holefors, A.; Marttila, S.; Dalman, K.; Lehrkinder, A.; Rur, M.; Rees-George, J.; Nybom, H.; Everett, K.; Scheper, R.W.A.; et al. Real-time PCR for detection and quantification, and histological characterization of Neonectria ditissima in apple trees. Trees 2016, 30, 1111–1125. [Google Scholar] [CrossRef]
- Xiwang, K.; Huang, L.; Han, Q.; Gao, X.; Kang, Z. Histological and cytological investigations of the infection and colonization of apple bark by Valsa mali var. mali. Australas. Plant Pathol. 2012, 42, 85–93. [Google Scholar]
- Blanchette, R.A.; Biggs, A.R. Defense Mechanisms of Woody Plants Against Fungi; Springer: Berlin/Heidelberg, Germany, 1992; 458p. [Google Scholar]
- Crowdy, S.H. Observations on apple canker III. The anatomy of the stem canker. Ann. Appl. Biol. 1949, 36, 483–495. [Google Scholar] [CrossRef]
- Biggs, A.; Davis, D.; Merrill, W. Histology of cankers on Populus caused by Cytospora chrysosperma. Can. J. Bot. 1983, 61, 563–574. [Google Scholar] [CrossRef]
- Sakamoto, Y.; Yamada, Y.; Sano, Y.; Tamai, Y.; Funada, R. Pathological anatomy of Nectria canker on Fraxinus mandshurica var. japonica. IAWA J. 2004, 25, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Zalasky, H. Penetration and initial establishment of Nectria galligena in aspen and peachleaf willow. Can. J. Bot. 1968, 46, 57–60. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Anamorph | Teleomorph | p-Value |
|---|---|---|---|
| FB density (min and max number per cluster) | 1–3 | 9–23 | |
| FB height (μm) | 824 ± 110 | 606 ± 134 | <0.0001 *** |
| FB width (μm) | 854 ± 215 | 166 ± 43 | <0.0001 *** |
| Phellem separation width (μm) | 690 ± 179 | 1012 ± 258 | <0.0001 *** |
| Phellem crack width (μm) | 418 ± 153 | 688 ± 263 | <0.0001 *** |
| Phellem crack length (μm) | 560 ± 266 | 1263 ± 1307 | 0.0029 ** |
| Phellem crack area (mm2) | 0.218 ± 0.134 | 0.455 ± 0.488 | 0.0086 ** |
| Trait | Anamorph | Teleomorph | p-Value |
|---|---|---|---|
| Phelloderm parenchyma cells | 1.5 ± 0.5 a | 4.1 ± 0.2 a | <0.0001 *** |
| Phloem parenchyma cells | 1.1 ± 0.5 b | 2.0 ± 0.9 b | <0.0001 *** |
| Xylem parenchyma cells | 1.6 ± 0.3 a | 1.6 ± 0.4 c | 0.9976 NS |
| Xylem vessels | 0.6 ± 0.5 c | 0.6 ± 0.5 e | 0.8725 NS |
| Xylem fibres | 1.3 ± 0.4 b | 1.3 ± 0.4 d | 0.7586 NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Račko, V.; Kováč, J.; Mišíková, O.; Mihál, I.; Milenković, I.; Ďurkovič, J. A Structural Assessment of Sycamore Maple Bark Disintegration by Nectria cinnabarina. Forests 2022, 13, 452. https://doi.org/10.3390/f13030452
Račko V, Kováč J, Mišíková O, Mihál I, Milenković I, Ďurkovič J. A Structural Assessment of Sycamore Maple Bark Disintegration by Nectria cinnabarina. Forests. 2022; 13(3):452. https://doi.org/10.3390/f13030452
Chicago/Turabian StyleRačko, Vladimír, Ján Kováč, Oľga Mišíková, Ivan Mihál, Ivan Milenković, and Jaroslav Ďurkovič. 2022. "A Structural Assessment of Sycamore Maple Bark Disintegration by Nectria cinnabarina" Forests 13, no. 3: 452. https://doi.org/10.3390/f13030452
APA StyleRačko, V., Kováč, J., Mišíková, O., Mihál, I., Milenković, I., & Ďurkovič, J. (2022). A Structural Assessment of Sycamore Maple Bark Disintegration by Nectria cinnabarina. Forests, 13(3), 452. https://doi.org/10.3390/f13030452