1. Introduction
The sustainability and biodiversity of forests depend on the quality of stock from forest nurseries. In Poland, foresters plant approximately half a billion seedlings every year based on the production of 800 million seedlings per year. Forecasts by the National Forest Cover Increasing Program assume an increasing the forest cover level from a current 30% to 33% by 2050 [
1]. This will require the afforestation of millions of hectares and the availability of huge quantities of viable seed for sowing in nurseries. Currently, forest nursery seed is sent to seed stations, where quality is determined based on batches of 400 germinated seeds. However, this method is labor- and time-consuming, and seed must be sent well in advance. What is lacking is a quick and efficient method of assessing the seed suitability for sowing. Automated devices such as electronic noses may be useful for quicker assessments on a large scale. These devices have already been used to assess food quality. Human noses can be very sensitive to odors [
2], and these abilities have developed during evolution to decrease the risk of consuming something that might be harmful. Odors can be emitted before fungal hyphae are visible on a plant surface and becomes visible to the naked eye. A seed may be non-viable due to damage or infection, but this is often not noticeable for seeds stored in dry conditions. Superficial visual inspection would not reveal the lack of viability which becomes apparent after seeds are sown and seedlings fail to emerge. We would like to develop a quick and efficient method to detect whether a seed sample is healthy and suitable for sowing or spoiled and should be further examined or discarded.
Since the introduction of the concept of an electronic nose [
3,
4,
5], which is a device consisting of an array of nonspecific gas sensors, equipped with machine learning pattern recognition algorithms, various applications of this rapid and non-invasive diagnostic tool have been proposed. Several reports review potential applications, challenges, and possible improvements of electronic noses in focusing on forestry and agriculture [
6,
7,
8,
9,
10,
11]. Applications of electronic nose for detection and identification of fungal species were reviewed by Mota et al. [
12]. Volatile organic compounds (VOC) emitted by various seeds infected by fungi have been analyzed with the help of electronic noses, including fungal contamination of cereal grain samples [
13,
14], rice [
15], and rapeseed [
16,
17].
The purpose of this research was to examine seeds of silver fir (
Abies alba) for viability via detection of degradative organisms via their VOC’s using an electronic nose. Silver fir is one of the main forest-forming species in Poland and is typical of the mountainous regions of central and southern Europe [
18], but it is threatened by climate change and increasing pest pressures such as bark beetles.
2. Silver Fir in the Białowieża Forest
There are only two known countries in Europe where silver fir grows naturally in low-lying areas: France (Normandy) and Poland (Yata, Topór, Mienia, Rudka, and Tisovik reserves) [
19,
20,
21,
22]. Forests growing in the lowlands dominate in Poland at 84.8%, while the area of mountainous forests is 8.5% [
23].
For almost 200 years, there has been a decline in wood resources and the area occupied by silver fir in European forests, known as the phenomenon of “the decline of silver fir in its natural range” [
24]. According to some studies, the period of stagnation in some regions of Europe (Serbia) reaches 330 years [
25]. It is assumed that this phenomenon was caused by a whole complex of biotic and abiotic factors, including the influence of humans [
18,
22,
24]. However, the actual cause remains unclear to this day. Since 1981–1989, there has been a positive trend in the status of silver fir stands both in Poland and throughout Europe, which has improved significantly [
24,
26].
In the Belarusian part of the Białowieża Forest (which means wider forest) is the Tisovik tract (until 1939, called Cisówka), which is of˙ interest because of two crucial circumstances: (1) this is the northernmost “island” of autochthonous silver fir growing in the lowlands (120 km from the nearest natural stand in the Jata Reserve) [
27,
28,
29]; and (2) the only currently surviving relict population of
A. alba is present in the Tisovik tract, which is a part of the Białowieża Forest, one of the largest forest areas in Central and Eastern Europe [
30,
31]. For various reasons the silver fir population has never been large. From 1823 to World War II the number of silver fir trees fluctuated between 100 and 300 [
32]. Currently only about 20 mature trees remain (unpublished data).
In 1922, Wiśniewski [
33] found another very small group of silver fir trees in the Hubar tract, of the Białoweieża Forest. According to the researcher, it was undoubtedly of natural origin. As we learned from the oral communication of colleagues from the National Park “Belavežskaja Pušča” (Belarus), this fragment has not survived.
According to Środoń [
34], who based, based his information on Wiśniewski [
33], the oldest silver fir tree, blown down by the wind in Tisovik in 1924, had a trunk circumference of 120 cm and a height of 33.5 m, corresponding to an age of at least 250 years. This means it became established in the second half of the 17th century. This tree was a descendant of a silver fir which began to produce seeds at about 70 years of age [
35]. Consequently, the beginning of the female parent’s life of this 250-year-old tree was at the end of the 16th to the beginning of the 17th century. Thus, based on these estimates, the age of the silver fir population in Tisovik is over 400 years, but is likely much longer. As noted by Paczoski [
28], the appearance of silver fir in the Forest dates back to the climatic optimum of the Holocene.
Such long preservation in the Forest of this unique stand, first described in 1829 by Górski [
27], and then studied in detail by Paczoski [
28], allows an assumption that particular genetic traits allowed for survival. However, it has often been suggested that this is a consequence of a favorable location, i.e., a small sandy island covered with forest on the watershed in the middle of impassable swamps and watercourses, which provided high air humidity and frequent indirect precipitation (dew, frost) [
32], as well as a high level of groundwater. However, for more than half a century now, those swamps have undergone senescence and infill, and the level of the groundwater has dropped significantly. Tisovik, currently does not have favorable hydrological conditions for silver fir [
36] and is at the extreme northern end of a range which allows silver fir to regenerate naturally [
32].
This unique relict population may have highly suitable genetic properties [
37] and is a promising source of seed and vegetative material. Thus, it can be used in practical forestry outside the mountainous part of the range of the studied species, and not only in the Białowieza Forest region, but in other parts of Poland first of all in the natural forest region Mazury-Podlaskie (II).
This situation becomes especially important and relevant against the background of the current mass extinction of Norway spruce (
Picea abies), especially in the northeastern part of its range [
38,
39]. The modern range of silver fir in Poland began in the sub-Boreal period of the Holocene and ended about 2000 years ago in the sub-Atlantic [
34,
40]. However, in the postglacial or in the so-called Little Ice Age (1300–1850), which came after the climatic optimum of the Holocene (about 800–1300 BP), Norway spruce (
Picea abies), a typical boreal species, displaced the silver fir, growing in admixture with the main species [
28].
Now, under conditions of global warming, only silver fir, which is similar to
P. abies in terms of ecological and biological properties, can be a possible alternative to this species. We have already mentioned the favorable growth conditions for the silver fir in Tisovik, and this area was the source of seeds for our study [
41], as well as from two other artificial silver fir stands of unknown geographical origin in the Polish part of the Białowieża Forest, but of an older age [
42].
It is known that one of the main stresses faced by
A. alba is low winter temperatures [
18,
35]. According to the predictive model of Vitasse et al. [
43], under the conditions of modern climate, silver fir in Europe is favored, as predictions are for warming characterized by a sharp increase in winter temperatures with a constant amount of precipitation [
36,
44]. Dyderski et al. [
45] also came to a similar conclusion, and they classified
A. alba as a “winner” while modeling the ranges of major European tree species by 2061–2080 and the level of threat they may face under different scenarios of climate change.
However, at present, there is still not enough silvicultural experience of silver fir in northern Poland [
46], although, in the middle of the 20th century, it grew on small plots, at least in 22 forestry enterprises in the western and middle part of the Mazury-Podlaskie forest region of northeastern Poland [
19].
A serious problem with efficient use of the germplasm of silver fir from Tisovik is that a low percentage of silver fir seeds can germinate. According to Korczyk [
29,
32], this ranges from 12.8% to 17.5%, which are rates similar to the 12.6% reported by Gonczarenko et al. [
47] for the Tisovik population [
47]. These authors also noted that this can vary from tree to tree (2.6% to 40.0%).
Another negative factor significantly affects the quality of silver fir seeds from Tisovik. According to Korczyk [
29,
32], the percentage of damage to mature seeds by the larvae of
Megastigmus suspectus Bor.,
Resseliella picea Seitn., and
Barbara herrichina Obr. is very significant, reaching the range of 62–77%. It is not unreasonable to assume that these pests have a complex of associates, represented by pathogenic fungi, capable of infecting seeds and other parts of trees.
A similar situation, for example, takes place with Siberian fir (
A. sibirica Ledb.), which is related to
A. alba. In recent years, it has been dying from the combined action of the ussuri polygraph (
Polygraphus proximus Blandf.) and associated ophiostomatoid fungi [
48,
49]. According to Russian researchers, this biotic tandem represents a new threat to fir forests in Siberia and throughout Europe [
48].
That is why it is so important to have a tool such as the electronic nose to assess seeds of silver fir. This will make it possible to determine the degree of their infection with pathogenic organisms quickly and take measures in advance to anticipate the influence of this negative factor, which can significantly reduce the germination rate of seeds. This will facilitate sufficient healthy planting material for a unique population of relict silver fir. The purpose of this research was to examine silver fir seeds for viability using an electronic nose and analyze the emitted VOC’s composition by the GC-MS method.
3. Materials and Methods
3.1. Seed Collection and Preparation
Cones were harvested on 20 September 2021 from one plantation of half-sib silver fir
Abies alba (Hajnówka forest district (Wilczy Jar sub-District, compartment 416ad) with geographical coordinates of E 23°39′17″, N 52°42′33″ (
Figure 1). These trees were established in 1996 from 4-year-old seedlings derived from seeds collected in 1992 in the Tisovik reserve (Belarus) of the Białowieża Forest [
50]. There were 11 mother trees giving rise to half-sib families of undetermined paternity.
Seed production in plantations was observed for the first time in 2019, and in the next two years (2020, 2021) they were collected for sowing in autumn in the nursery of the Hajnówka forest enterprise, located in Białowieża Primeval. Cones were collected (20 September 2021), (
Figure 2) and seeds were isolated from˙ cones (30 October 2021) (
Figure 3). The seeds were stored at room temperature until experiments began (2 November 2021). The experiments were carried out using seeds of half-sib family No. 17. After collection, the cones were stored in a dry, well-ventilated room at a temperature of 20–25 °C, where they naturally dried. Artificial regulation of the photoperiod was not used. Under these dry conditions, it is less likely that the seeds have become contaminated by air spora which were able to establish.
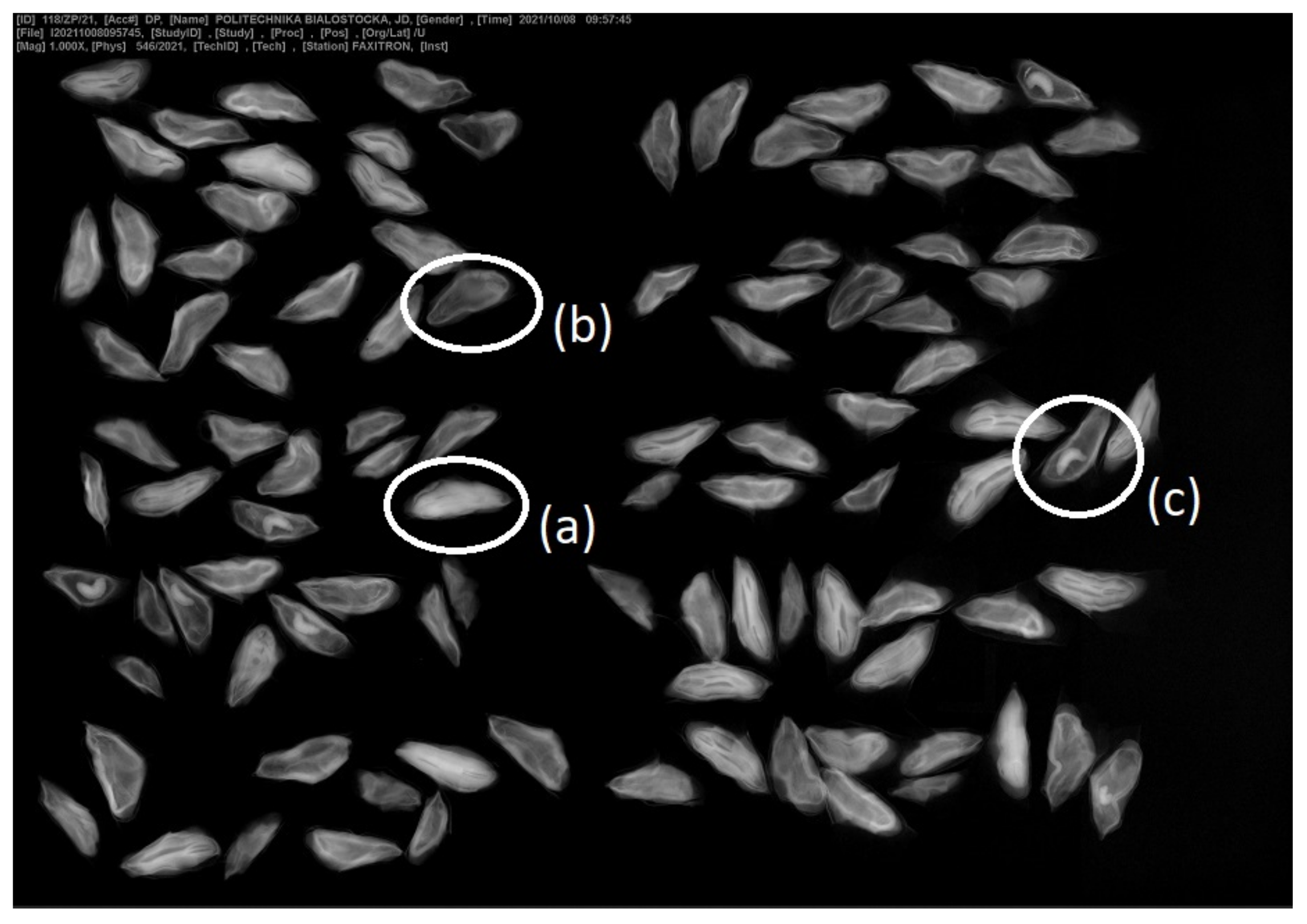
For the analysis of seed germination viability, a sample was taken from the fraction of clean seeds. Approximately 49 kg of the mixture of seeds with hulls dried at room temperature 20–25 °C were sent to the Kostrzyca Forest Gene Bank. From this mixture, 7.61 kg of pure seeds were obtained. X-ray analysis has been performed in Kostrzyca (
Figure 4 to verify the quality of seeds and the presence of insects. A sample of 400 cleaned seeds, consisting of 4 replicates of 100 seeds each was tested [
51] for germination dynamics and seed quality.
Investigated seeds were surface sterilized by immersion in 75% propanol for thirty seconds and then rinsed with sterile distilled water for one minute. Six hundred seeds were chosen, and lots of up to 30 seeds were placed in 9-cm diameter Petri dishes on moist sterile paper towels. The plates were lidded but not tightly sealed. The papers towels were periodically moistened with sterile distilled water. Twenty Petri dishes were prepared in this way.
3.2. Fungal Selection and Identification
After incubation of the 20 Petri dishes with seeds, nine plates were selected for further investigation based on homogenous appearance, with similar fungal colonies appearing in the sets of plates. In addition to these plates, several other plates were selected for identification by subculturing and DNA sequencing. Genomic DNA was extracted from hyphae of three green and three white colonies. Then, genomic DNA was extracted using the NucleoSpin Plant II kit (Macherey-Nagel, Düren, Germany) according to the manufacturer’s instructions.
The region of the fungal internal transcribed spacer (ITS) was amplified with primers ITS1 (5
-TCCGTAGGTGAACCTGCGG-3
), and ITS4 (5
-TCCTCCGCTTATTGATATGC-3
) [
52]. PCR was performed using the TaqNova- RED kit (BLIRT, Gdańsk, Poland). The 20
L PCR mix consisted of 10 µL 2X TaqNova- RED mix, 2
L 5
M of each primer, 2
L DNA extracts, and 2
L H
O. Cycling was performed using a Veriti 96-well thermal cycler (ThermoFisher Scientific, Waltham, MA, USA) as follows: an initial denaturation step at 95 °C for 3 min, followed by 30 cycles (95 °C for 30 s, 55 °C for 30 s, and elongation at 72 °C for 30 s), and a final extension step at 75 °C for 5 min. Excess dNTPs and unincorporated primers were removed from the PCR product using CleanPCR (BLIRT, Gdańsk, Poland). DNA was eluted in 40
L H
O. The amplified products were sequenced using the Sanger sequencing method in our laboratory.
Sequencing PCR reactions were performed using 1
L BigDye Terminator v. 3.1 Ready Reaction Mix (ThermoFisher Scientific), 2
L BigDye Sequencing Buffer (ThermoFisher Scientific), 1
L (5
M) ITS1 or ITS4 primer, and H
O to bring the total volume to 10
L. The thermal profile for the sequencing reactions consisted of initial denaturation step of 96 °C for 1 min, followed by 25 cycles of 96 °C for 10 s, 50 °C for 5 s, and 60 °C for 105 s. The rDNA region was sequenced using an ABI 3500 xL genetic analyzer (ThermoFisher Scientific). For the sequences obtained a 98% alignment threshold over at least 440 base pairs was applied for species identification. Assignment of the obtained sequences to species was performed using the BOLDSYSTEMS identification engine [
53,
54].
3.3. Measurement of Volatile Organic Compounds
Volatile organic compounds (VOC) emitted from seeds were analyzed by headspace solid-phase microextraction coupled with gas chromatography and mass spectrometry (HS-SPME/GC-MS) following a previously used method [
55,
56] briefly described below.
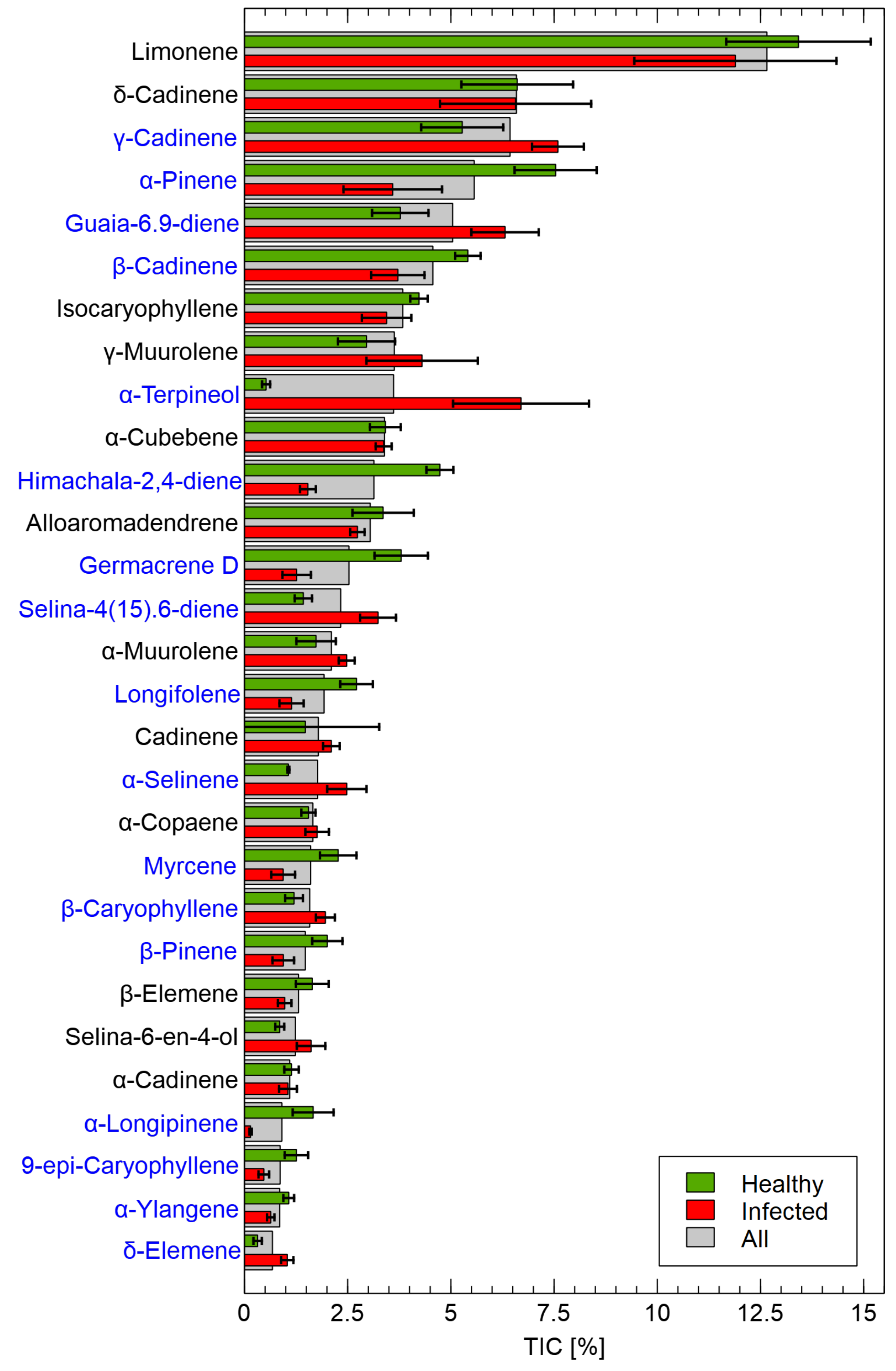
In preliminary studies of VOCs emitted from asymptomatic silver fir seeds (control), divinylbenzene/carboxen/polydimethylsiloxane (DVB/CAR/PDMS), CAR/PDMS, and PDMS sorption fibres (Supelco, Bellefonte, PA, USA) were compared and DVB/CAR/PDMS fibre was selected for further research based on the highest effectiveness of the extraction–desorption cycle. Seeds for chemical analysis were selected at random, targeting those seeds with visible fungal growth. Seeds (1 ± 0.05 g, ~8 seeds) were placed in 60 mL glass vial with cap and septum (Büchi 049535) and heated at 40 °C for 60 min. SPME fiber was then added to the vial with a divinylbenzene/carboxene/polydimethylsiloxan stationary phase (Supelco, Bellefonte, PA, USA). The fiber was exposed to the headspace gas phase at 40 °C for 30 min. Immediately after exposure, the SPME fibre was inserted into an injection port of the GC-MS instrument for 10 min. GC–MS analyses were performed using an Agilent 7890A gas chromatograph with an Agilent 5975C mass spectrometer (Agilent Technologies Inc., Santa Clara, CA, USA). The injector was operated at a temperature of 250 °C in splitless mode. Chromatographic separation was performed on a capillary column HP -5MS (30 m × 0.25 mm × 0.25 m) at a helium flow rate of 1 mL/min. The initial temperature of the column was 35 °C and increased to 250 °C at a rate of 5 °C/min. The ion source and quadrupole temperatures were 230 °C and 150 °C, respectively. The electron impact mass spectra were obtained at ionization energy of 70 eV. Detection was performed in full scan mode for a range of 29–600 atomic mass units.
The peaks from the chromatogram were integrated, and the percentage of the components in the total ion current (TIC) was calculated. The mass spectral data and calculated retention indices were used to identify the components. Mass spectrometric identification was performed using the NIST (2020) and Wiley (2020) mass spectral libraries and the collections of Adams (2007) and Tkachev (2008) as well as a private, unpublished library of mass spectra, which had been created using standard chemical compounds. The retention indices of the analytes were determined considering the retention times of the n-alkanes. In a separate run, the mixture of C5C40 n-alkanes (1 L) was injected onto the chromatographic column and separated under the conditions previously described for GC–MS analyses of volatiles. Linear temperature-programmed experimental retention indices (RIexp) were calculated using the following expression RI = t t t t, where n is the number of carbon atoms in the alkane, t is the retention time of the analyte, t is the retention time of the n-alkane eluting immediately before the analyte, and t is the retention time of the n-alkane eluting directly after the analyte. The experimental retention indices (RI) were compared with those in the retention indices database (RI ) mentioned above.
3.4. Electronic Nose Measurements
3.4.1. Electronic Nose Device
The electronic nose used in these measurements was the PEN3 device (Airsense Analytics GmbH, Schwerin, Germany) [
57]. It is a commercially available electronic nose, widely used in scientific laboratories, based on 10 metal oxide sensors that work only at high temperatures (about 350 °C and 500 °C). The sensors are sensitive to a wide range of gases, as listed in
Table A1 in
Appendix A. The unit has a very efficient air pump system with two inlets: one for the odor sampling and another for ambient air. The second is filtered through an activated carbon filter and is used as a reference signal level or to dilute the samples. The PEN3 was connected to a computer and controlled by Airsense WinMuster 1.6.2 software. It was also responsible for the acquisition of measurement data.
The measurement data of the PEN3 electronic nose were values or the conductivity of the sensors as a function of time when the sensors responded to the change in the chemical composition of the gas to which they were exposed. At the beginning of each measurement, the sensor matrix was purged with clean air, and then the air was pulled into the device, with a chamber flow rate of 7.7 mL/min. The values collected were unitless, as these were the magnitudes normalised by the baseline response of the sensor to clean air conditions measured just before the sensor was exposed to sampled air.
3.4.2. Samples Measurements
The electronic nose was turned on at least one hour before the measurement to ensure properly heated sensors. To ensure that no residue of old measurement samples remained in the sensor chamber, PEN3 automatically purges the sensors with filtered air for 180 s before starting the measurement. The zero point of the sensors was then measured, and during the next 120 s, the instrument recorded the response of the sensors to the constant flow of headspace gas from the measured sample. The sensor signals recorded by the software were G/G0. The conductance of the sensor during the measurement was divided by the conductance in ambient air.
The fir seed samples were kept in Petri dishes in which the infected seeds had been incubated (
Figure 5). The number of seeds varied between individual samples, but in all cases the fungal hyphae covered approximately half or more of the seed surfaces in each 9 cm diameter Petri dish. We attempted standardize the samples by the amount of hyphal growth rather than by the number of seeds as we assumed that odor emission was related to hyphal growth. Variation in the amount of biological material in samples was an additional source of noise giving higher variability to the sensor signals collected by the electronic nose. This was related to the variability of the intensity of gases emitted by samples, but, to a lesser extent, to the chemical composition of the gases.
Immediately before measurements, the Petri dish to be measured was half-opened and the PEN3 electronic nose tube was placed near the center at 1 cm above a seed. The air above the seeds which should have contained volatile organic components emitted by the seeds and fungi, was pulled through the tube into the electronic nose device. In the interval between the 24-h measurements, the Petri dishes were kept closed and sealed with parafilm.
One or two measurements of each sample were taken every day. With two measurements per day, one series of measurements was performed in the morning and the second in the late afternoon to allow build up of VOC from the seeds. The order of samples in each series of measurement was randomized.
3.5. Analysis of Electronic Nose Data
In our experiments, 1200 units of data were collected in a single sample measurement since this corresponded to the number of readings of the sensor conductance within 2 min with a 1 s interval, multiplied by the number of sensors in the electronic nose device. The results of measurements collected during the experiment, discussed in
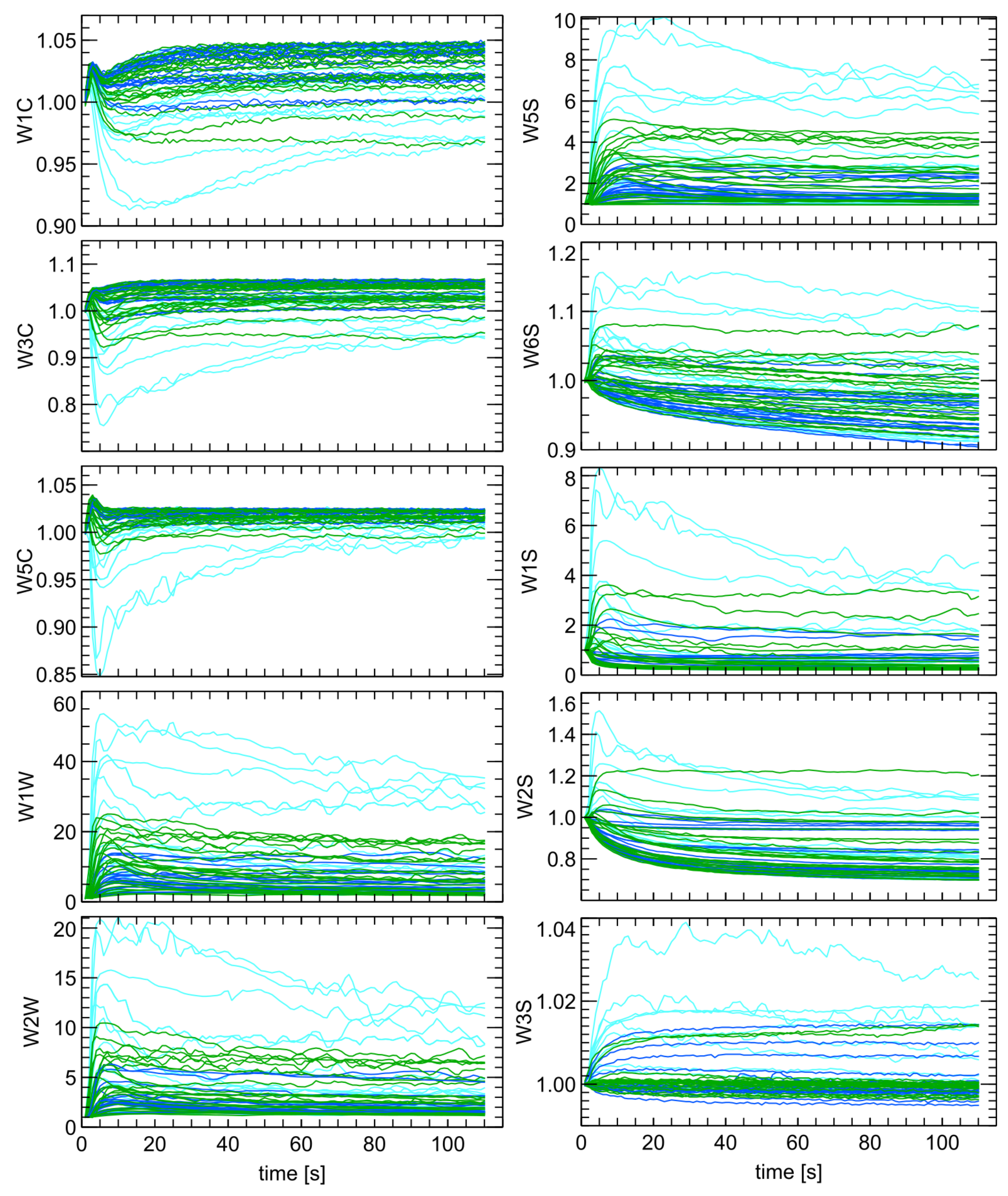
Section 4.6.1, indicated the presence of measurement noise. We then preprocessed the data using the exponential smoothing method.
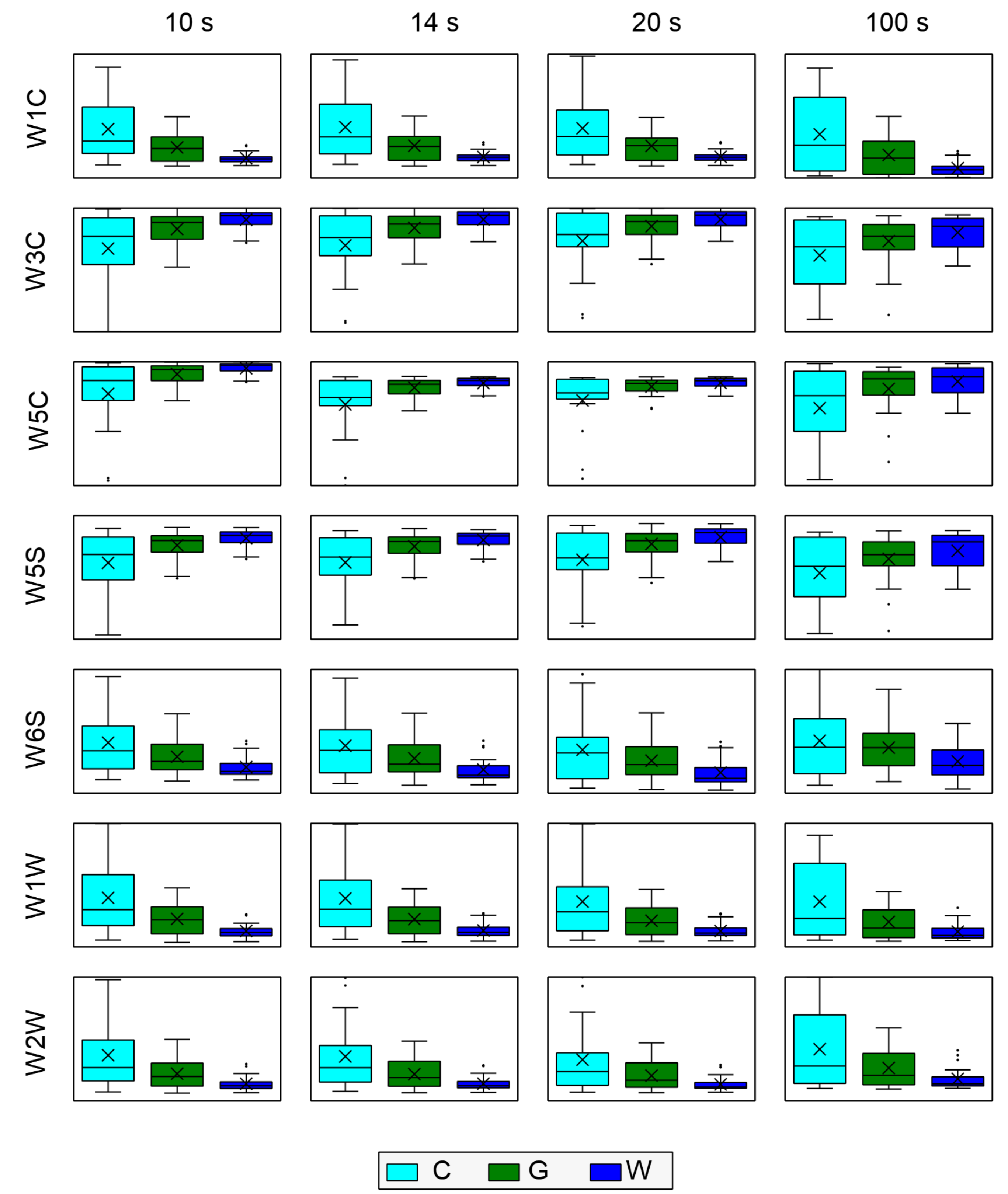
In our research, we used a subset of points from the smoothed curves for further analysis. First, we selected the response values between the 4th and 20th s in 2 s intervals. We also selected the values at the end of data collection at the 80th and 100th s. Thus, we obtained 10 features for each sensor. With this approach, we captured the main features of the overall sensor response due to the presence of the measured odor while significantly reducing the dimensionality of the problem. These features were then used as input for machine learning classification modeling.
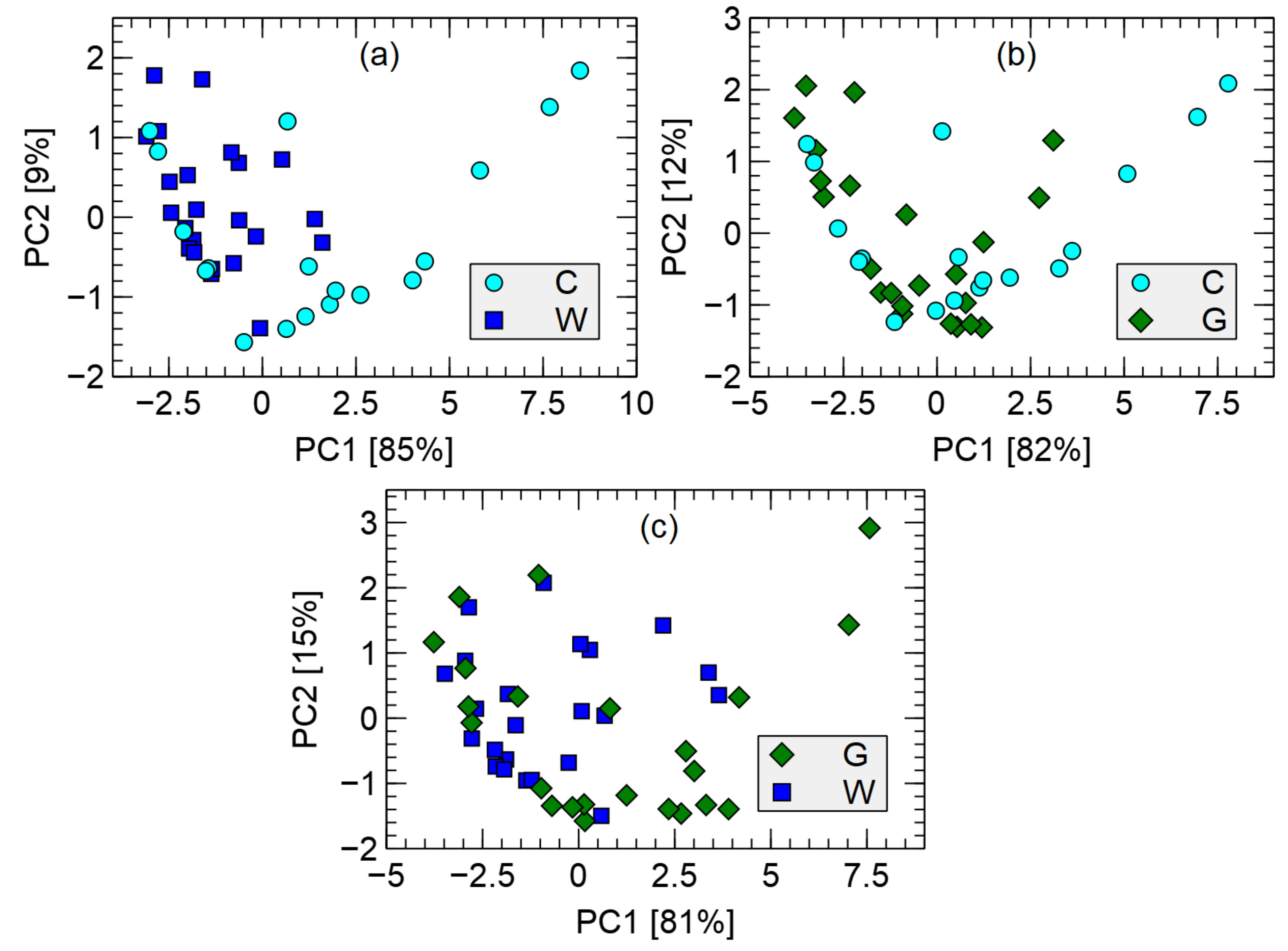
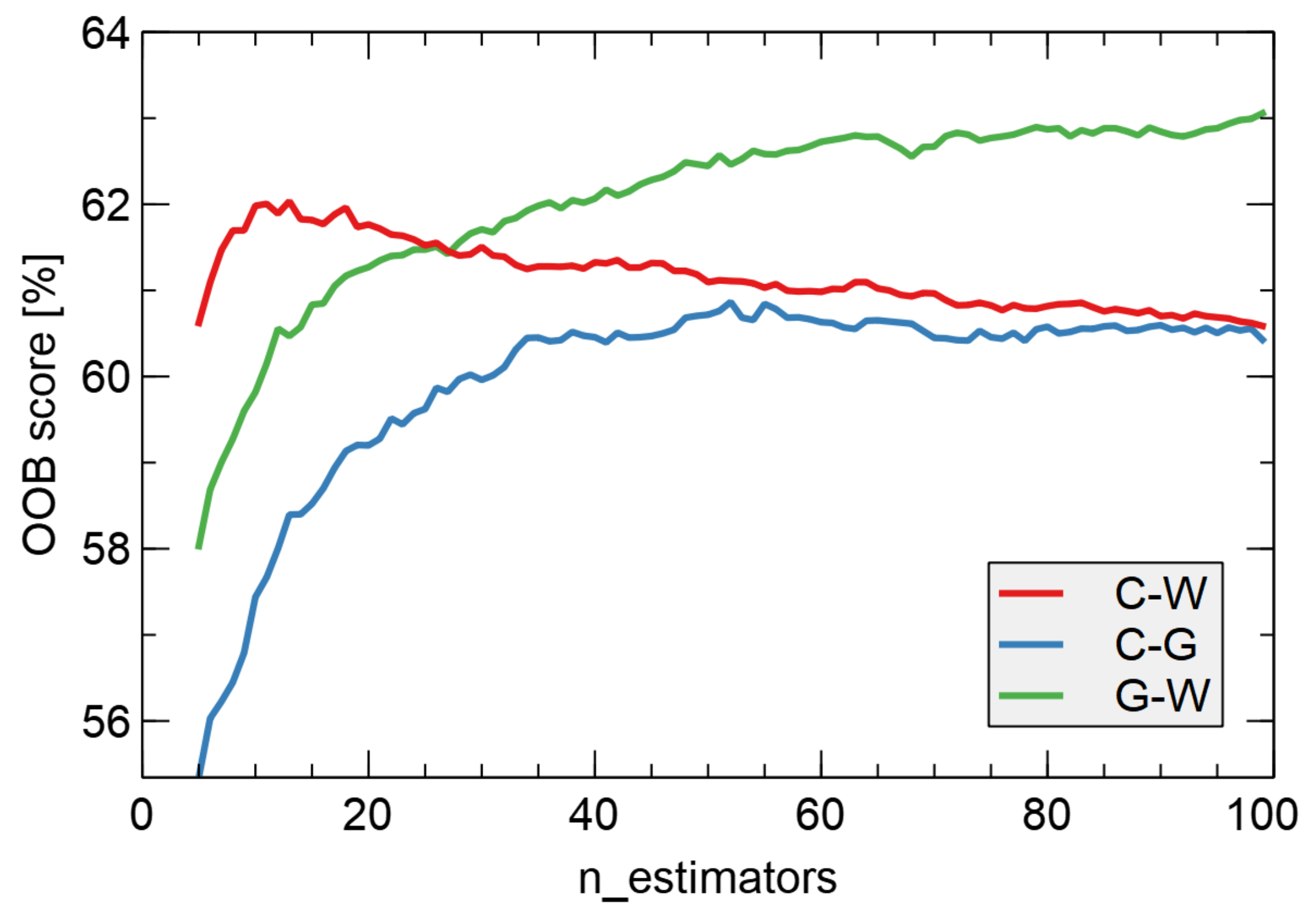
Electronic nose measurements aim to use the collected data to build classification models capable of discriminating between the samples under study. This objective would make it possible to use such devices to detect the presence of odors in the environment and, in this way, evaluate the possibility of the presence of pathogens. For this task, we used a random forest model of machine learning [
58].
Random forest is a popular machine learning algorithm used for classification tasks. It fits a number of decision tree classifiers on various sub-samples of the dataset and/or sub-samples of modeling features and uses averaging of the results. That approach allows to improve the predictive accuracy and control over-fitting.
3.6. Visualization of Distribution of Sensor Response Data
One of the random forest model outputs is a ranking of the most important features used for classification. In our analysis, we used these variables to visualize the distribution of the measured data in different ways. We compared the measured sensor response at a few characteristic moments of time elapsed from the beginning of sensor exposure to the measured gas. For such comparisons of distribution of data we used box-plot diagrams.
In addition, we transformed the modeling features using the principal component analysis (PCA) method. The PCA is one of the most commonly statistical techniques used for visualizing this type of data and allows us to gain some intuitive insights into the relationship between the patterns in the distribution of the data points. The input data for the PCA analysis methods are the sensor response magnitudes obtained at different time moments and possibly from different sensors. These values should not be directly compared when the data were collected from different sensors. The modeling features should be scaled to obtain the same variance for the PCA transformation. We used the top variables selected by the random forest model for the PCA transformation.
3.7. Data Processing and Analysis
All analyses of the data collected by the electronic nose measurements presented in this report were performed with Python 3.8 language codes, using statistical analysis methods from the scikit-learn module [
59]. Processing and statistical analysis of data collected by the GC-MS measurements were performed using SAS 9.4 (SAS Institute, Cary, NC, USA) software using the SAS Enterprise Guide user interface and SAS/Stat procedures. PROC TTEST was used to verify statistical significance of differences in concentration of chemical components between uninfected and infected samples. Satterthwaite statistics, not assuming equal variance between groups, was used for calculation of
p-value [
60].


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}











