
Wood Formation under Changing Environment: Omics Approaches to Elucidate the Mechanisms Driving the Early-to-Latewood Transition in Conifers




Abstract
:1. Introduction
1.1. Xylogenesis, Ring Width, Xylem Traits, and Global Warming: What Is the Matter?
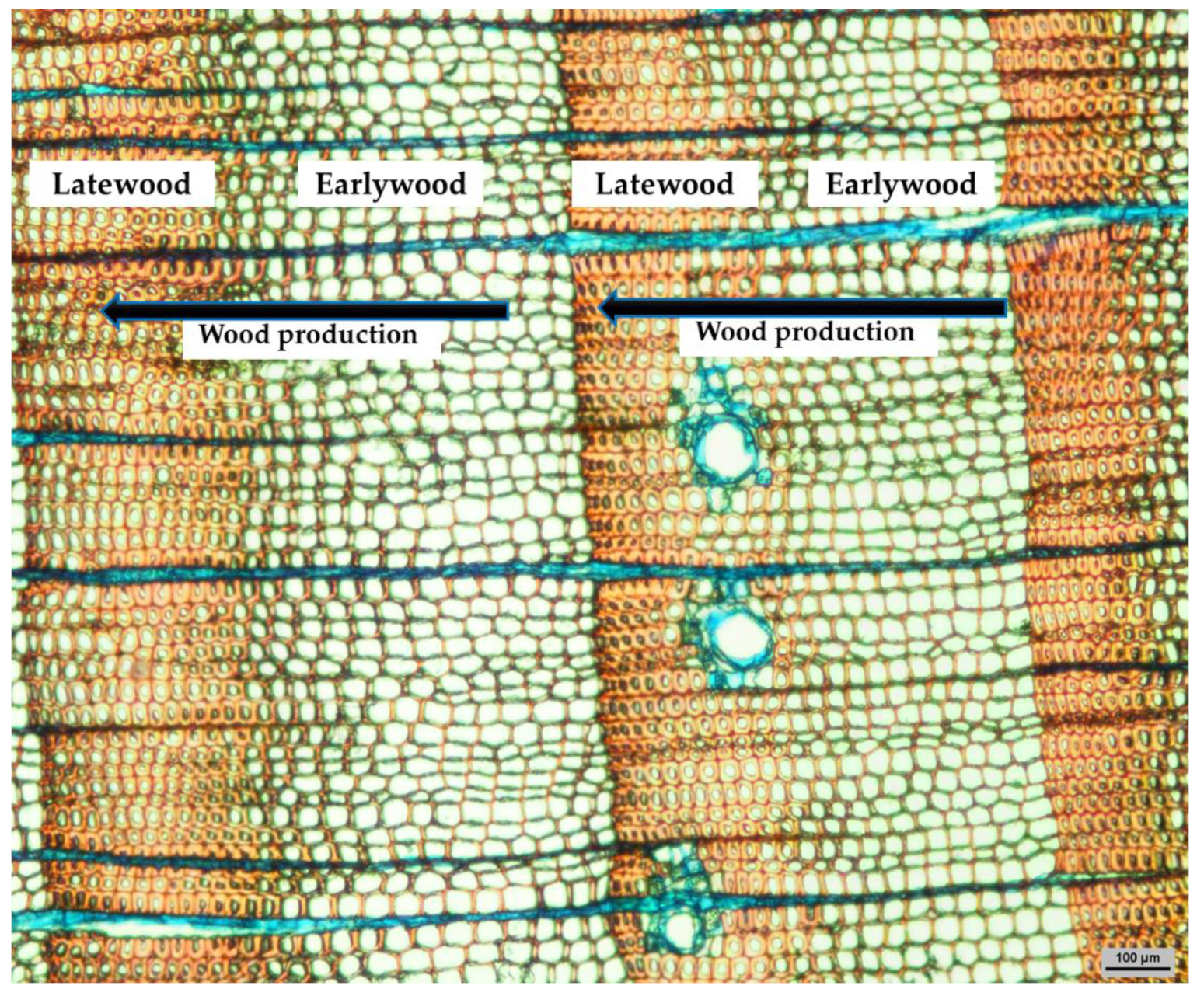
1.2. Earlywood vs. Latewood Traits in Conifers: The Control of the Environmental Cues over Phenophases?
1.3. How Could Genomics Help to Disentangle the Physiological Processes Related to the Early-to-Latewood Transition?
2. Genomic Resources for Conifer Wood Formation Studies
2.1. Available Conifer Genomes
2.2. Transcriptomic Resources and Functional Genomic Studies
2.3. Proteomic Resources
3. Disentangling the Molecular Mechanisms Underlying the Early-to-Latewood Transition in the Omics Era
4. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- IPCC. Summary for policy makers. In Global Warming of 1.5 C; Masson-Delmotte, V., Zhai, P., Pörtner, H.O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Moufouma-Okia, W., Péan, C., Pidcock, R., et al., Eds.; World Meteorological Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Vincent, L.; Wang, X.; Milewska, E.; Wan, H.; Yang, F.; Swail, V. A second generation of homogenized Canadian monthly surface air temperature for climate trend analysis. J. Geophys. Res. Atmos. 2012, 117, D18110. [Google Scholar] [CrossRef]
- Wang, X.; Feng, Y.; Vincent, L. Observed changes in one-in-20 year extremes of Canadian surface air temperatures. Atmos. Ocean 2014, 52, 222–231. [Google Scholar] [CrossRef]
- Tabari, H. Climate change impact on flood and extreme precipitation increases with water availability. Sci. Rep. 2020, 10, 13768. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-G.; Ma, Q.; Rossi, S.; Biondi, F.; Deslauriers, A.; Fonti, P.; Liang, E.; Mäkinen, H.; Oberhuber, W.; Rathgeber, C.B.K.; et al. Photoperiod and temperature as dominant environmental drivers triggering secondary growth resumption in Northern Hemisphere conifers. Proc. Natl. Acad. Sci. USA 2020, 117, 20645–20652. [Google Scholar] [CrossRef]
- Allen, C.; Macalady, A.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H.; Biber, P.; Shutze, G.; Kemmerer, J.; Uhl, E. Wood density reduced while wood volume accelerated in Central European forests since 1870. Forest Ecol. Manag. 2018, 429, 589–616. [Google Scholar] [CrossRef]
- Gričar, J.; Čufar, K.; Eler, K.; Gryc, V.; Vavrcík, H.; De Luis, M.; Prislan, P. Transition dates from earlywood to latewood and early phloem to late phloem in Norway Spruce. Forests 2021, 12, 331. [Google Scholar] [CrossRef]
- Zobel, B.J.; Van Buitenen, J.P. Wood Variation, Its Causes and Control; Springer: Berlin/Heidelberg, Germany, 1989; p. 363. [Google Scholar]
- Zobel, B.J.; Jett, J.B. Genetics of Wood Production; Springer: Berlin/Heidelberg, Germany, 1995. [Google Scholar]
- Carrer, M.; Castagneri, D.; Prendin, A.; Petit, G.; Von Arx, G. Retrospective analysis of wood anatomical traits reveals a recent extension in tree cambial activity in two high-elevation conifers. Front. Plant Sci. 2017, 8, 737. [Google Scholar] [CrossRef] [Green Version]
- Björklund, J.; Seftigen, K.; Schweingruber, F.; Fonti, P.; Von Arx, G.; Bryukhanova, M.V.; Cuny, H.E.; Carrer, M.; Castagneri, D.; Frank, D.C. Cell size and wall dimensions drive distinct variability of earlywood and latewood density in Northern Hemisphere conifers. New Phytol. 2017, 216, 728–740. [Google Scholar] [CrossRef] [Green Version]
- Rosell, J.A.; Olson, M.E.; Anfodillo, T. Scaling of xylem vessel diameter with plant size: Causes, predictions, and outstanding questions. Curr. For. Rep. 2017, 3, 46–59. [Google Scholar] [CrossRef]
- Li, S.; Li, X.; Link, R.; Li, R.; Deng, L.; Schuldt, B.; Jiang, X.; Zhao, R.; Zheng, J.; Li, S.; et al. Influence of cambial age and axial height on the spatial patterns of xylem traits in Catalpa bungei, a ring-porous tree species native to China. Forests 2019, 10, 662. [Google Scholar] [CrossRef] [Green Version]
- Spicer, R.; Gartner, B. The effects of cambial age and position within the stem on specific conductivity in Douglas-fir (Pseudotsuga menziesii) sapwood. Trees 2001, 15, 222–229. [Google Scholar] [CrossRef]
- Rahman, M.; Begum, S.; Nakaba, S.; Yamagishi, Y.; Kudo, K.; Nabeshima, E.; Nugroho, W.; Oribe, Y.; Funada, R. Relationship between the earlywood-to-latewood transition and changes in levels of stored starch around the cambium in locally heated stems of the evergreen conifer Chamaecyparis pisifera. Trees Struct. Funct. 2016, 30, 1619–1631. [Google Scholar] [CrossRef]
- Gričar, J.; Zupancic, M.; Čufar, K.; Koch, G.; Schmitt, U.; Oven, P. Effect of local heating and cooling on cambial activity and cell differentiation in the stem of Norway spruce (Picea abies). Ann. Bot. 2006, 97, 943–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gričar, J.; Zupancic, M.; Čufar, K.; Oven, P. Regular cambial activity and xylem and phloem formation in locally heated and cooled stem portions of Norway spruce. Wood Sci. Technol. 2007, 41, 463–475. [Google Scholar] [CrossRef]
- Begum, S.; Nakaba, S.; Yamagishi, Y.; Yamane, K.; Islam, A.; Oribe, Y.; Ko, J.; Jin, H.; Funada, R. A rapid decrease in temperature induces latewood formation in artificially reactivated cambium of conifer stems. Ann. Bot. 2012, 110, 875–885. [Google Scholar] [CrossRef] [Green Version]
- Giovannelli, A.; Mattana, S.; Emiliani, G.; Anichini, M.; Traversi, M.L.; Pavone, F.S.; Cicchi, R. Localized stem heating from the rest to growth phase induces latewood-like cell formation and slower stem radial growth in Norway spruce saplings. Tree Physiol. 2021, tpab116. [Google Scholar] [CrossRef]
- Delpierre, N.; Lireux, S.; Hartig, F.; Camarero, J.J.; Cheaib, A.; Čufar, K.; Cuny, H.; Deslauriers, A.; Fonti, P.; Gričar, J.; et al. Chilling and forcing temperatures interact to predict the onset of wood formation in northern hemisphere conifers. Glob. Chang. Biol. 2019, 25, 1089–1105. [Google Scholar] [CrossRef]
- Ford, K.; Harrington, C.; Bansal, S.; Gould, P.; St Clair, J. Will changes in phenology track climate change? A study of growth initiation timing in coast Douglas-fir. Glob. Chang. Biol. 2016, 22, 3712–3723. [Google Scholar] [CrossRef]
- Lebourgeois, F. Climatic signals in earlywood and total ring width of Corsican pine from western France. Ann. For. Sci. 2000, 57, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Melcher, P.; Holbrook, N.M.; Burns, M.J.; Zwieniecki, A.M.; Cobb, A.R.; Brodribb, T.J.; Choat, B.; Sack, L. Measurements of stem xylem hydraulic conductivity in the laboratory and field. Methods Ecol. Evol. 2012, 3, 685–694. [Google Scholar] [CrossRef]
- Sperry, J.S.; Hacke, U.G.; Pittermann, J. Size and function in conifer tracheids and angiosperm vessels. Am. J. Bot. 2006, 93, 1490–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domec, J.C.; Gartner, B.L. Age and position-related changes in hydraulic vs. mechanical dysfunction of xylem: Inferring the design criteria for Douglas-fir wood structure. Tree Physiol. 2002, 22, 91–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacke, U.G.; Jansen, S. Embolism resistance of three boreal conifer species varies with pit structure. New Phytol. 2009, 182, 675–686. [Google Scholar] [CrossRef]
- Fukatsu, E.; Nakada, R. The timing of latewood formation determines the genetic variation of wood density in Larix kaempferi. Trees 2018, 32, 1233–1245. [Google Scholar] [CrossRef]
- Plomion, C.; Leprovost, G.; Stokes, A. Wood formation in trees. Plant Physiol. 2001, 127, 1513–1523. [Google Scholar] [CrossRef]
- Rathgeber, C.B.K.; Cuny, H.E.; Fonti, P. Biological basis of tree-ring formation: A crash course. Front. Plant Sci. 2016, 7, 734. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Anfodillo, T.; Čufar, K.; Cuny, H.E.; Deslauriers, A.; Fonti, P.; Frank, D.; Gričar, J.; Gruber, A.; Huang, J.G.; et al. Pattern of xylem phenology in conifers of cold ecosystems at the northern hemisphere. Glob. Chang. Biol. 2016, 22, 3804–3813. [Google Scholar] [CrossRef] [Green Version]
- Bertaud, F.; Holmbom, B. Chemical composition of earlywood and latewood in Norway spruce heartwood, sapwood and transition zone wood. Wood Sci. Technol. 2004, 38, 245–256. [Google Scholar] [CrossRef]
- Antonova, G.F.; Varaksina, T.N.; Zheleznichenko, T.V.; Stasova, V.V. Lignin deposition during earlywood and latewood formation in Scots pine stems. Wood Sci. Technol. 2014, 48, 919–936. [Google Scholar] [CrossRef]
- Andersson, S.; Serimaa, R.; Torkkeli, M.; Paakkari, T.; Saranpaa, P.; Pesonen, E. Microfibril angle of Norway spruce [Picea abies (L.) Karst.] compression wood: Comparison of measuring techniques. J. Wood Sci. 2000, 46, 343–349. [Google Scholar] [CrossRef]
- Cuny, H.E.; Rathgeber, C.B. Xylogenesis: Coniferous trees of temperate forests are listening to the climate tale during the growing season but only remember the last words! Plant Physiol. 2016, 171, 306–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buttò, V.; Deslauriers, A.; Rossi, S.; Rozenberg, P.; Shishov, V.; Morin, H. The role of plant hormones in tree-ring formation. Trees 2020, 34, 315–335. [Google Scholar] [CrossRef]
- Hartmann, F.P.; Rathgeber, C.B.; Badel, E.; Fournier, M.; Moulia, B. Modelling the spatial crosstalk between two biochemical signals explains wood formation dynamics and tree-ring structure. J. Exp. Bot. 2021, 72, 1727–1737. [Google Scholar] [CrossRef]
- Uggla, C.; Magel, E.; Moritz, T.; Sundberg, B. Function and dynamics of auxin and carbohydrates during earlywood/latewood transitions in Scots pine. Plant Physiol. 2001, 125, 2029–2039. [Google Scholar] [CrossRef] [Green Version]
- Bhalerao, R.P.; Fischer, U. Auxin gradients across wood—instructive or incidental? Physiol. Plant. 2014, 151, 43–51. [Google Scholar] [CrossRef]
- Funada, R.; Kubo, T.; Tabuchi, M.; Sugiyama, T.; Fushitani, M. Seasonal variations in endogenous indole-3-acetic acid and abscisic acid in the cambial region of Pinus densiflora Sieb. et Zucc. stems in relation to earlywood-latewood transition and cessation of tracheid production. Holzforschung 2001, 55, 128–134. [Google Scholar] [CrossRef]
- Gray, W.M.; Östin, A.; Sandberg, G.; Romano, C.P.; Estelle, M. High temperature promotes auxin-mediated hypocotyl elongation in Arabidopsis. Proc. Natl. Acad. Sci. USA 1998, 95, 7197–7202. [Google Scholar] [CrossRef] [Green Version]
- Mellerowicz, E.J.; Baucher, M.; Sundberg, B.; Bojeran, W. Unraveling cell wall formation in the woody dicot stem. Plant Mol. Biol. 2001, 47, 239–274. [Google Scholar] [CrossRef]
- Fajstavr, M.; Paschová, Z.; Giagli, K.; Vavrčík, H.; Gryc, V.; Urban, J. Auxin (IAA) and soluble carbohydrate seasonal dynamics monitored during xylogenesis and phloemogenesis in Scots pine. iForest 2018, 11, 553. [Google Scholar] [CrossRef]
- Traversari, S.; Emiliani, G.; Traversi, M.L.; Anichini, M.; Giovannelli, A. Pattern of carbohydrate changes in maturing xylem and phloem during growth to dormancy transition phase in Picea abies (L.) Karst. Dendrobiology 2018, 80, 12–23. [Google Scholar] [CrossRef]
- Sundberg, B.; Ericsson, A.; Little, C.H.A.; Nasholm, T.; Gref, R. The relationship between crown size and ring width in Pinus sylvestris L. stems: Dependence on indole-3-acetic acid, carbohydrates and nitrogen in the cambial region. Tree Physiol. 1993, 12, 347–362. [Google Scholar] [CrossRef] [PubMed]
- Delmer, D.P.; Haigler, C.H. The regulation of metabolic flux to cellulose, a major sink for carbon in plants. Metab. Eng. 2002, 4, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Pascual, M.B.; El-Azaz, J.; De La Torre, F.N.; Cañas, R.A.; Avila, C.; Cánovas, F.M. Biosynthesis and metabolic fate of phenylalanine in conifers. Front. Plant Sci. 2016, 7, 1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grattapaglia, D.; Silva-Junior, O.B.; Resende, R.T.; Cappa, E.P.; Muller, B.S.F.; Tan, B.; Isik, F.; Ratcliffe, B.; El-Kassaby, Y.A. Quantitative genetics and genomics converge to accelerate forest tree breeding. Front. Plant Sci. 2018, 9, 1693. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Dungey, H.S. Expected benefit of genomic selection over forward selection in conifer breeding and deployment. PLoS ONE 2018, 13, e0208232. [Google Scholar] [CrossRef] [Green Version]
- Soro, A.; Lenz, P.; Hassegawa, M.; Roussel, J.R.; Bousquet, J.; Achim, A. Genetic influence on components of wood density variation in white spruce. Int. J. For. Res. 2021, 95, 153–165. [Google Scholar] [CrossRef]
- Baison, J.; Zhou, L.; Forsberg, N.; Mörling, T.; Grahn, T.; Olsson, L.; Karlsson, B.; Wu, H.X.; Mellerowicz, E.J.; Lundqvist, S.; et al. Genetic control of tracheid properties in Norway spruce wood. Sci. Rep. 2020, 10, 18089. [Google Scholar] [CrossRef]
- Du, Q.; Lu, W.; Quan, M.; Xiao, L.; Song, F.; Li, P.; Zhou, D.; Xie, J.; Wang, L.; Zhang, D. Genome-Wide Association Studies to improve wood properties: Challenges and prospects. Front. Plant Sci. 2018, 9, 1912. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.Q.; Zan, Y.; Milesi, P.; Zhou, L.; Chen, J.; Li, L.; Cui, B.; Niu, S.; Westin, J.; Karlsson, B.; et al. Leveraging breeding programs and genomic data in Norway spruce (Picea abies L. Karst) for GWAS analysis. Genome Biol. 2021, 22, 179. [Google Scholar] [CrossRef]
- Cheng, S.; Melkonian, M.; Smith, S.A.; Brockington, S.; Archibald, J.M.; Delaux, P.M.; Li, F.W.; Melkonian, B.; Mavrodiev, E.V.; Sun, W.; et al. 10KP: A phylodiverse genome sequencing plan. GigaScience 2018, 7, giy013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewin, H.A.; Robinson, G.E.; Kress, W.J.; Baker, W.J.; Coddington, J.; Crandall, K.A.; Durbin, R.; Edwards, S.V.; Forest, F.; Gilbert, M.T.P.; et al. Earth BioGenome Project: Sequencing life for the future of life. Proc. Natl. Acad. Sci. USA 2018, 115, 4325–4333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De La Torre, A.R.; Birol, I.; Bousquet, J.; Ingvarsson, P.K.; Jansson, S.; Jones, S.J.; Keeling, C.I.; MacKay, J.; Nilsson, O.; Ritland, K.; et al. Insights into conifer giga-genomes. Plant Physiol. 2014, 166, 724–732. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Shang, L.; Zhu, Q.H.; Fan, L.; Guo, L. Twenty years of plant genome sequencing: Achievements and challenges. Trends Plant Sci. 2021, 27, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Nystedt, B.; Street, N.R.; Wetterbom, A.; Zuccolo, A.; Lin, Y.C.; Scofield, D.G.; Douglas, G.; Vezii, F.; Delhome, N.; Giacomello, S.; et al. The Norway spruce genome sequence and conifer genome evolution. Nature 2013, 497, 579–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Berkum, N.L.; Lieberman-Aiden, E.; Williams, L.; Imakaev, M.; Gnirke, A.; Mirny, L.A.; Dekker, J.; Lander, E.S. Hi-C: A method to study the three-dimensional architecture of genomes. J. Vis. Exp. 2010, 6, 1869. [Google Scholar] [CrossRef] [Green Version]
- Barseghyan, H.; Tang, W.; Wang, R.T.; Almalvez, M.; Segura, E.; Bramble, M.S.; Lipson, A.; Douine, E.D.; Lee, H.; Délot, E.C.; et al. Next-generation mapping: A novel approach for detection of pathogenic structural variants with a potential utility in clinical diagnosis. Genome Med. 2017, 9, 90. [Google Scholar] [CrossRef] [Green Version]
- Kaul, S.; Koo, H.L.; Jenkins, J.; Rizzo, M.; Rooney, T.; Tallon, L.J.; Feldblyum, T.; Nierman, W.; Benito, M.; Lin, X.; et al. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar]
- Tuskan, G.A.; Difazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar]
- Ahmar, S.; Ballesta, P.; Ali, M.; Mora-Poblete, F. Achievements and challenges of genomics-assisted breeding in forest trees: From marker-assisted selection to genome editing. Int. J. Mol. Sci. 2021, 22, 10583. [Google Scholar] [CrossRef]
- Doudna, J.A.; Charpentier, E. Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef] [PubMed]
- Poovaiah, C.; Phillips, L.; Geddes, B.; Reeves, C.; Sorieul, M.; Thorlby, G. Genome editing with CRISPR/Cas9 in Pinus radiata (D. Don). BMC Plant Biol. 2021, 21, 363. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, D.A.; Feranchuk, S.I.; Sharov, V.V.; Cybin, A.N.; Makolov, S.V.; Putintseva, Y.A.; Oreshkova, N.V.; Krutovsky, K.V. Stepwise large genome assembly approach: A case of Siberian larch (Larix sibirica Ledeb). BMC Bioinform. 2019, 20, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birol, I.; Raymond, A.; Jackman, S.D.; Pleasance, S.; Coope, R.; Taylor, G.A.; Yuen, M.M.; Keeling, C.I.; Brand, D.; Vandervalk, B.P.; et al. Assembling the 20 Gb white spruce (Picea glauca) genome from whole-genome shotgun sequencing data. Bioinformatics 2013, 29, 1492–1497. [Google Scholar] [CrossRef]
- Warren, R.L.; Keeling, C.I.; Yuen, M.M.; Raymond, A.; Taylor, G.A.; Vandervalk, B.P.; Mohamadi, H.; Paulino, D.; Chiu, R.; Jackman, S.D.; et al. Improved white spruce (Picea glauca) genome assemblies and annotation of large gene families of conifer terpenoid and phenolic defense metabolism. Plant J. 2015, 83, 189–212. [Google Scholar] [CrossRef]
- Stevens, K.A.; Wegrzyn, J.L.; Zimin, A.; Puiu, D.; Crepeau, M.; Cardeno, C.; Paul, R.; Gonzalez-Ibeas, D.; Koriabine, M.; E Holtz-Morris, A.; et al. Sequence of the Sugar Pine Megagenome. Genetics 2016, 204, 1613–1626. [Google Scholar] [CrossRef]
- Zimin, A.; Stevens, K.A.; Crepeau, M.W.; Holtz-Morris, A.; Koriabine, M.; Marçais, G.; Puiu, D.; Roberts, M.; Wegrzyn, J.L.; De Jong, P.J.; et al. Sequencing and assembly of the 22-gb loblolly pine genome. Genetics 2014, 196, 875–890. [Google Scholar] [CrossRef] [Green Version]
- Zimin, A.V.; Stevens, K.A.; Crepeau, M.W.; Puiu, D.; Wegrzyn, J.L.; Yorke, J.A.; Langley, C.H.; Neale, D.B.; Salzberg, S.L. An improved assembly of the loblolly pine mega-genome using long-read single-molecule sequencing. GigaScience 2017, 6, giw016. [Google Scholar]
- Neale, D.B.; McGuire, P.E.; Wheeler, N.C.; Stevens, K.A.; Crepeau, M.W.; Cardeno, C.; Zimin, A.V.; Puiu, D.; Pertea, G.M.; Sezen, U.U.; et al. The Douglas-Fir genome sequence reveals specialization of the photosynthetic apparatus in Pinaceae. G3 Genes Genomes Genet. 2017, 7, 3157–3167. [Google Scholar] [CrossRef] [Green Version]
- Neale, D.B.; Zimin, A.V.; Zaman, S.; Scott, A.D.; Shrestha, B.; Workman, R.E.; Puiu, D.; Allen, B.J.; Moore, Z.J.; Sekhwal, M.K.; et al. Assembled and annotated 26.5 Gbp coast redwood genome: A resource for estimating evolutionary adaptive potential and investigating hexaploid origin. G3 Genes Genomes Genet. 2022, 12, kab380. [Google Scholar] [CrossRef]
- Scott, A.D.; Zimin, A.V.; Puiu, D.; Workman, R.; Britton, M.; Zaman, S.; Caballero, M.; Read, A.C.; Bogdanove, A.J.; Burns, E.; et al. A reference genome sequence for Giant Sequoia. G3 Genes|Genomes|Genetics 2020, 10, 3907–3919. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Gou, J.; Liao, Q.; Li, Y.; Zhou, Q.; Bi, G.; Li, C.; Du, R.; Wang, X.; Sun, T.; et al. The Taxus genome provides insights into paclitaxel biosynthesis. Nat. Plants 2021, 7, 1026–1036. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Fu, F.; Yang, L.; Niu, Y.; Tian, Z.; He, X.; Yang, X.; Chen, J.; Sun, W.; Wan, T.; et al. Taxus yunnanensis genome offers insights into gymnosperm phylogeny and taxol production. Commun. Biol. 2021, 4, 1203. [Google Scholar] [CrossRef] [PubMed]
- Wan, T.; Liu, Z.M.; Li, L.F.; Leitch, A.R.; Leitch, I.J.; Lohaus, R.; Liu, Z.-J.; Xin, H.-P.; Gong, Y.-B.; Liu, Y.; et al. A genome for gnetophytes and early evolution of seed plants. Nat. Plants 2018, 4, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Wan, T.; Liu, Z.; Leitch, I.J.; Xin, H.; Maggs-Kölling, G.; Gong, Y.; Li, Z.; Marais, E.; Liao, Y.; Dai, C.; et al. The Welwitschia genome reveals a unique biology underpinning extreme longevity in deserts. Nat. Commun. 2021, 12, 4247. [Google Scholar] [CrossRef]
- Niu, S.; Li, J.; Bo, W.; Yang, W.; Zuccolo, A.; Giacomello, S.; Chen, X.; Han, F.; Yang, J.; Song, Y.; et al. The Chinese pine genome and methylome unveil key features of conifer evolution. Cell 2022, 185, 204–217. [Google Scholar] [CrossRef]
- Cheng, J.; Wang, X.; Liu, X.; Zhu, X.; Li, Z.; Chu, H.; Wang, Q.; Lou, Q.; Cai, B.; Yang, Y.; et al. Chromosome-level genome of Himalayan yew provides insights into the origin and evolution of the paclitaxel biosynthetic pathway. Mol. Plant. 2021, 14, 1199–1209. [Google Scholar] [CrossRef]
- Mosca, E.; Cruz, F.; Gómez-Garrido, J.; Bianco, L.; Rellstab, C.; Brodbeck, S.; Csilléry, K.; Fady, B.; Fladung, M.; Fussi, B.; et al. A reference genome sequence for the European Silver Fir (Abies alba Mill.): A community-generated genomic resource. G3 Genes|Genomes|Genetics 2019, 9, 2039–2049. [Google Scholar] [CrossRef] [Green Version]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Heitkam, T.; Schulte, L.; Weber, B.; Liedtke, S.; Breitenbach, S.; Kögler, A.; Morgenstern, K.; Brückner, M.; Tröber, U.; Wolf, H.; et al. Comparative repeat profiling of two closely related conifers (Larix decidua and Larix kaempferi) reveals high genome similarity with only few fast-evolving satellite DNAs. Front. Genet. 2021, 12, 683668. [Google Scholar] [CrossRef]
- Lin, D.; Coombe, L.; Jackman, S.; Gagalova, K.; Warren, R.; Hammond, S.; McDonald, H.; Kirk, H.; Pandoh, P.; Zhao, Y.; et al. Complete chloroplast genome sequence of an Engelmann spruce (Picea engelmannii) genotype from western Canada. Microbiol. Resour. Announc. 2019, 8, e00382–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, T.; Coombe, L.; Lin, D.; Warren, R.; Kirk, H.; Pandoh, P.; Zhao, Y.; Moore, R.; Mungall, A.; Ritland, C.; et al. Complete chloroplast genome sequence of a black spruce (Picea mariana) from Eastern Canada. Microbiol. Resour. Announc. 2020, 9, e00877–20. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, H.H.; Harms, L.; Epp, L.S.; Mewes, N.; Bernhardt, N.; Kruse, S.; Stoof-Leichsenring, K.R.; Pestryakova, L.A.; Wieczorek, M.; Trense, D.; et al. Chloroplast and mitochondrial genetic variation of larches at the Siberian tundra-taiga ecotone revealed by de novo assembly. PLoS ONE 2019, 14, e0216966. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Grewe, F.; Fan, W.; Young, G.J.; Knoop, V.; Palmer, J.D.; Mower, J.P. Ginkgo and Welwitschia mitogenomes reveal extreme contrasts in gymnosperm mitochondrial evolution. Mol. Biol. Evol. 2016, 33, 1448–1460. [Google Scholar] [CrossRef] [Green Version]
- Jackman, S.D.; Coombe, L.; Warren, R.L.; Kirk, H.; Trinh, E.; MacLeod, T.; Pleasance, S.; Pandoh, P.; Zhao, Y.; Coope, R.J.; et al. Complete mitochondrial genome of a gymnosperm, Sitka spruce (Picea sitchensis), indicates a complex physical structure. Genome Biol. Evol. 2020, 12, 1174–1179. [Google Scholar] [CrossRef]
- Kan, S.L.; Shen, T.T.; Gong, P.; Ran, J.H.; Wang, X.Q. The complete mitochondrial genome of Taxus cuspidate (Taxaceae): Eight protein-coding genes have transferred to the nuclear genome. BMC Evol. Biol. 2020, 20, 10. [Google Scholar] [CrossRef] [Green Version]
- Kan, S.L.; Shen, T.T.; Ran, J.H.; Wang, X.Q. Both Conifer II and Gnetales are characterized by a high frequency of ancient mitochondrial gene transfer to the nuclear genome. BMC Biol. 2021, 19, 146. [Google Scholar] [CrossRef]
- Falk, T.; Herndon, N.; Grau, E.; Buehler, S.; Richter, P.; Zaman, S.; Baker, E.M.; Ramnath, R.; Ficklin, S.; Staton, M.; et al. Growing and cultivating the forest genomics database, TreeGenes. Database 2018, 2018, bay084. [Google Scholar] [CrossRef]
- Kirst, M.; Johnson, A.F.; Baucom, C.; Ulrich, E.; Hubbard, K.; Staggs, R.; Paule, C.; Retzel, E.; Whetten, R.; Sederoff, R. Apparent homology of expressed genes from wood-forming tissues of loblolly pine (Pinus taeda L.) with Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2003, 100, 7383–7388. [Google Scholar] [CrossRef] [Green Version]
- Paiva, J.A.P.; Garcés, M.; Alves, A.; Garnier-Géré, P.; Rodrigues, J.C.; Lalanne, C.; Porcon, S.; Le Provost, G.; Da Silva Perez, D.; Brach, J.; et al. Molecular and phenotypic profiling from the base to the crown in maritime pine wood-forming tissue. New Phytol. 2008, 78, 283–301. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wu, H.X.; Dillon, S.K.; Southerton, S.G. Generation and analysis of expressed sequence tags from six developing xylem libraries in Pinus radiata D. Don. BMC Genom. 2009, 10, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, I.K.; Wegrzyn, J.L.; Rajora, O.P. Generation, functional annotation and comparative analysis of black spruce (Picea mariana) ESTs: An important conifer genomic resource. BMC Genom. 2013, 14, 702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Fung-Leung, W.P.; Bittner, A.; Ngo, K.; Liu, X. Comparison of RNA-Seq and microarray in transcriptome profiling of activated T cells. PLoS ONE 2014, 9, e78644. [Google Scholar] [CrossRef] [PubMed]
- Chano, V.; Collada, C.; Soto, A. Transcriptomic analysis of wound xylem formation in Pinus canariensis. BMC Plant Biol. 2017, 17, 234. [Google Scholar] [CrossRef] [Green Version]
- Pavan, S.; Delvento, C.; Ricciardi, L.; Lotti, C.; Ciani, E.; D’Agostino, N. Recommendations for choosing the genotyping method and best practices for quality control in crop Genome-Wide Association Studies. Front. Genet. 2020, 11, 447. [Google Scholar] [CrossRef]
- Bernhardsson, C.; Zan, Y.; Chen, Z.; Ingvarsson, P.K.; Wu, H.X. Development of a highly efficient 50K single nucleotide polymorphism genotyping array for the large and complex genome of Norway spruce (Picea abies L. Karst) by whole genome resequencing and its transferability to other spruce species. Mol. Ecol. Resour. 2021, 21, 880–896. [Google Scholar] [CrossRef]
- Howe, G.T.; Jayawickrama, K.; Kolpak, S.E.; Kling, J.; Trappe, M.; Hipkins, V.; Ye, T.; Guida, S.; Cronn, R.; Cushman, S.A.; et al. An axiom SNP genotyping array for Douglas-fir. BMC Genom. 2020, 21, 9. [Google Scholar] [CrossRef] [Green Version]
- Perry, A.; Wachowiak, W.; Downing, A.; Talbot, R.; Cavers, S. Development of a single nucleotide polymorphism array for population genomic studies in four European pine species. Mol. Ecol. Resour. 2020, 20, 1697–1705. [Google Scholar] [CrossRef]
- Plomion, C.; Bartholomé, J.; Lesur, I.; Boury, C.; Rodríguez-Quilón, I.; Lagraulet, H.; Ehrenmann, F.; Bouffier, L.; Gion, J.M.; Grivet, D.; et al. High-density SNP assay development for genetic analysis in maritime pine (Pinus pinaster). Mol. Ecol. Resour. 2016, 16, 574–587. [Google Scholar] [CrossRef]
- Pavy, N.; Gagnon, F.; Deschênes, A.; Boyle, B.; Beaulieu, J.; Bousquet, J. Development of highly reliable in silico SNP resource and genotyping assay from exome capture and sequencing: An example from black spruce (Picea mariana). Mol. Ecol. Resour. 2016, 16, 588–598. [Google Scholar] [CrossRef]
- Pavy, N.; Gagnon, F.; Rigault, P.; Blais, S.; Deschênes, A.; Boyle, B.; Pelgas, B.; Deslauriers, M.; Clément, S.; Lavigne, P.; et al. Development of high-density SNP genotyping arrays for white spruce (Picea glauca) and transferability to subtropical and nordic congeners. Mol. Ecol. Resour. 2013, 13, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Lamara, M.; Raherison, E.; Lenz, P.; Beaulieu, J.; Bousquet, J.; MacKay, J. Genetic architecture of wood properties based on association analysis and co-expression networks in white spruce. New Phytol. 2016, 210, 240–255. [Google Scholar] [CrossRef] [Green Version]
- Kastally, C.; Niskanen, A.K.; Perry, A.; Kujala, S.T.; Avia, K.; Cervantes, S.; Haapanen, M.; Kesälahti, R.; Kumpula, T.A.; Mattila, T.M.; et al. Taming the massive genome of Scots pine with PiSy50k, a new genotyping array for conifer research. Plant J. 2021, 109, 1337–1350. [Google Scholar] [CrossRef] [PubMed]
- Moreno, P.; Fexova, S.; George, N.; Manning, J.R.; Miao, Z.; Mohammed, S.; Muñoz-Pomer, A.; Fullgrabe, A.; Bi, Y.; Bush, N.; et al. Expression Atlas update: Gene and protein expression in multiple species. Nucleic Acids Res. 2022, 50, D129–D140. [Google Scholar] [CrossRef] [PubMed]
- Jokipii-Lukkari, S.; Sundell, D.; Nilsson, O.; Hvidsten, T.R.; Street, N.R.; Tuominen, H. NorWood: A gene expression resource for evo-devo studies of conifer wood development. New Phytol. 2017, 216, 482–494. [Google Scholar] [CrossRef] [Green Version]
- One Thousand Plant Transcriptomes Initiative. One thousand plant transcriptomes and the phylogenomics of green plants. Nature 2019, 574, 679–685. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, E.J.; Matasci, N.; Ayyampalayam, S.; Wu, S.; Sun, J.; Yu, J.; Jimenez Vieira, F.R.; Bowler, C.; Dorrell, R.G.; Gitzendanner, M.A.; et al. Access to RNA-sequencing data from 1173 plant species: The 1000 plant transcriptomes initiative (1KP). GigaScience 2019, 8, giz126. [Google Scholar] [CrossRef]
- Plomion, C.; Pionneau, C.; Brach, J.; Costa, P.; Bailleres, H. Compression wood- responsive proteins in developing xylem of maritime pine (Pinus pinaster Ait.). Plant Physiol. 2000, 123, 959–970. [Google Scholar] [CrossRef] [Green Version]
- Gion, J.M.; Lalanne, C.; Le Provost, G.; Ferry-Dumazet, H.; Paiva, J.; Chaumeil, P.; Frigerio, J.M.; Brach, J.; Barré, A.; De Daruvar, A.; et al. The proteome of maritime pine wood forming tissue. Proteomics 2005, 5, 3731–3751. [Google Scholar] [CrossRef]
- Paiva, J.A.P.; Garnier-Geéreé, P.H.; Rodrigues, J.C.; Alves, A.; Santos, S.; Graça, J.; Le Provost, G.; Chaumeil, P.; da Silva-Perez, D.; Bosc, A.; et al. Plasticity of maritime pine (Pinus pinaster) wood-forming tissues during a growing season. New Phytol. 2008, 179, 1180–1194. [Google Scholar] [CrossRef] [Green Version]
- Galindo González, L.M.; El Kayal, W.; Ju, C.J.; Allen, C.C.; King-Jones, S.; Cooke, J.E. Integrated transcriptomic and proteomic profiling of white spruce stems during the transition from active growth to dormancy. Plant Cell Environ. 2012, 35, 682–701. [Google Scholar] [CrossRef] [PubMed]
- Mast, S.; Peng, L.; Jordan, T.W.; Flint, H.; Phillips, L.; Donaldson, L.; Strabala, T.J.; Wagner, A. Proteomic analysis of membrane preparations from developing Pinus radiata compression wood. Tree Physiol. 2010, 30, 1456–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, H.T.; Weinberg, C.S.; Macdonald, L.J.; Adams, P.D.; Petzold, C.J.; Strabala, T.J.; Wagner, A.; Heazlewood, J.L. Golgi enrichment and proteomic analysis of developing Pinus radiata xylem by free-flow electrophoresis. PLoS ONE 2013, 8, e84669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, A.R.; Ukrainetz, N.K.; Kang, K.Y.; Mansfield, S.D. Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trials reveals strong environmental and weak genetic variation. New Phytol. 2007, 174, 762–773. [Google Scholar] [CrossRef]
- Eckert, A.J.; Wegrzyn, J.L.; Cumbie, W.P.; Goldfarb, B.; Huber, D.A.; Tolstikov, V.; Fiehn, O.; Neale, D.B. Association genetics of the loblolly pine (Pinus taeda, Pinaceae) metabolome. New Phytol. 2012, 193, 890–902. [Google Scholar] [CrossRef]
- Hall, R.D.; D’Auria, J.C.; Silva Ferreira, A.C.; Gibon, Y.; Kruszka, D.; Mishra, P.; Van De Zedde, R. High-throughput plant phenotyping: A role for metabolomics? Trends Plant Sci. 2022, 2, S1360-1385(22)00030-9. [Google Scholar] [CrossRef]
- Hawkins, C.; Ginzburg, D.; Zhao, K.; Dwyer, W.; Xue, B.; Xu, A.; Rice, S.; Cole, B.; Paley, S.; Karp, P.; et al. Plant Metabolic Network 15: A resource of genome-wide metabolism databases for 126 plants and algae. J. Integr. Plant Biol. 2021, 63, 1888–1905. [Google Scholar] [CrossRef]
- Raherison, E.S.; Giguère, I.; Caron, S.; Lamara, M.; MacKay, J.J. Modular organization of the white spruce (Picea glauca) transcriptome reveals functional organization and evolutionary signatures. New Phytol. 2015, 207, 172–187. [Google Scholar] [CrossRef] [Green Version]
- Cartenì, F.; Deslauriers, A.; Rossi, S.; Morin, H.; De Micco, V.; Mazzoleni, S.; Giannino, F. The physiological mechanisms behind the earlywood-to-latewood transition: A process-based modeling approach. Front. Plant Sci. 2018, 9, 1053. [Google Scholar] [CrossRef]
- Egertsdotter, U.; Van Zyl, L.M.; MacKay, J.; Peter, G.; Kirst, M.; Clark, C.; Whetten, R.; Sederoff, R. Gene expression during formation of earlywood and latewood in loblolly pine: Expression profiles of 350 genes. Plant Biol. 2004, 6, 654–663. [Google Scholar] [CrossRef]
- Yang, S.H.; Loopstra, C.A. Seasonal variation in gene expression for loblolly pines (Pinus taeda) from different geographical regions. Tree Physiol. 2005, 25, 1063–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Wu, H.X.; Southerton, S.G. Identification of putative candidate genes for juvenile wood density in Pinus radiata. Tree Physiol. 2012, 32, 1046–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Wu, H.X.; Southerton, S.G. Transcriptome profiling of wood maturation in Pinus radiata identifies differentially expressed genes with implications in juvenile and mature wood variation. Gene 2011, 487, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, H.X.; Southerton, S.G. Seasonal reorganization of the xylem transcriptome at different tree ages reveals novel insights into wood formation in Pinus radiata. New Phytol. 2010, 187, 764–776. [Google Scholar] [CrossRef]
- Emiliani, G.; Traversi, M.L.; Anichini, M.; Giachi, G.; Giovannelli, A. Transcript accumulation dynamics of phenylpropanoid pathway genes in the maturing xylem and phloem of Picea abies during latewood formation. J. Integr. Plant Biol. 2011, 53, 783–799. [Google Scholar] [CrossRef]
- He, S.; Xie, Y.; Sun, X.; Zhang, S. Comparative transcriptome analyses reveal candidate genes regulating wood quality in Japanese larch (Larix kaempferi). J. For. Res. 2020, 31, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Jokipii-Lukkari, S.; Delhomme, N.; Schiffthaler, B.; Mannapperuma, C.; Prestele, J.; Nilsson, O.; Street, N.R.; Tuominen, H. Transcriptional roadmap to seasonal variation in wood formation of Norway spruce. Plant Physiol. 2018, 176, 2851–2870. [Google Scholar] [CrossRef] [Green Version]
- Mishima, K.; Fujiwara, T.; Iki, T.; Kuroda, K.; Yamashita, K.; Tamura, M.; Fujisawa, Y.; Watanabe, A. Transcriptome sequencing and profiling of expressed genes in cambial zone and differentiating xylem of Japanese cedar (Cryptomeria japonica). BMC Genom. 2014, 15, 5920. [Google Scholar] [CrossRef] [Green Version]
- Baison, J.; Vidalis, A.; Zhou, L.; Chen, Z.Q.; Li, Z.; Sillanpää, M.J.; Bernhardsson, C.; Scofield, D.; Forsberg, N.; Grahn, T.; et al. Genome-wide association study identified novel candidate loci affecting wood formation in Norway spruce. Plant J. 2019, 100, 83–100. [Google Scholar] [CrossRef]
- González-Martínez, S.C.; Wheeler, N.C.; Ersoz, E.; Nelson, C.D.; Neale, D.B. Association genetics in Pinus taeda LI Wood property traits. Genetics 2007, 175, 399–409. [Google Scholar] [CrossRef] [Green Version]
- D’Odorico, P.; Besik, A.; Wong, C.Y.S.; Isabel, N.; Ensminger, I. High-throughput drone-based remote sensing reliably tracks phenology in thousands of conifer seedlings. New Phytol. 2020, 226, 1667–1681. [Google Scholar] [CrossRef] [PubMed]
- Santini, F.; Kefauver, S.C.; Araus, J.L.; Resco de Dios, V.; Martín García, S.; Grivet, D.; Voltas, J. Bridging the genotype-phenotype gap for a Mediterranean pine by semi-automatic crown identification and multispectral imagery. New Phytol. 2021, 229, 245–258. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; Miguel, C.; Chaves, I.; António, C. Mass spectrometry-based forest tree metabolomics. Mass Spectrom. Rev. 2021, 40, 126–157. [Google Scholar] [CrossRef] [PubMed]
- Nanasato, Y.; Mikami, M.; Futamura, N.; Endo, M.; Nishiguchi, M.; Ohmiya, Y.; Konagaya, K.; Taniguchi, T. CRISPR/Cas9-mediated targeted mutagenesis in Japanese cedar (Cryptomeria japonica D. Don). Sci. Rep. 2021, 11, 16186. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Species | Assembly Status | Year of First Publication/Release | Reference(s) | Genbank Assembly Database | Genbank Accession Number |
|---|---|---|---|---|---|
| Larix kaempferi (Japanese larch) | Contig | 2020 | - | ASM1317126v2 | GCA_013171265.2 |
| Larix sibirica (Siberian larch) | Scaffold | 2019 | [66] | LarixSibirica0.1 | GCA_004151065.1 |
| Picea abies (Norway spruce) | Scaffold | 2013 | [58] | Pabies01 | GCA_900067695.1 |
| Picea engelmannii (Engelmann’s spruce) | Scaffold | 2020 | - | Se404-851_v1 | GCA_009831015.1 |
| Picea glauca (white spruce) | Contig | 2013 | [67,68] | PG29_v5 | GCA_000411955.6 |
| Picea sitchensis (Sitka spruce) | Contig | 2020 | - | SNQJ01 | GCA_010110895.1 |
| Pinus lambertiana (sugar pine) | Scaffold | 2016 | [69] | Sugar pine JHU assembly | GCA_001447015.2 |
| Pinus taeda (loblolly pine) | Scaffold | 2014 | [70,71] | Ptaeda2.0 | GCA_000404065.3 |
| Pseudotsuga menziesii (Douglas-fir) | Scaffold | 2017 | [72] | DougFir1.0 | GCA_001517045.1 |
| Sequoia sempervirens (Coast redwood) | Scaffold | 2022 | [73] | SESE.2.2 | GCA_007258455.2 |
| Sequoiadendron giganteum (Giant sequoia) | Chromosome | 2020 | [74] | SEGI.2.0 | GCA_007115665.2 |
| Taxus chinensis (Chinese yew) | Chromosome | 2021 | [75] | Ta-2021 | GCA_019776745.2 |
| Taxus wallichiana var. yunnanensis (Himalayan yew) | Chromosome | 2021 | [76] | ASM1834077v1 | GCA_018340775.1 |
| Database Name | Content | Link |
|---|---|---|
| TreeGenes database | Genomic assemblies and raw sequences and annotation for:
Comparative genomic tools | https://treegenesdb.org [91] |
| Norway spruce genome project—Congenie | Genomic assemblies and raw sequences and annotation for Picea abies Blast searches Comparative genomic tools | https://congenie.org/ |
| Spruce-Up Project and SMarTForests | Genomic assemblies and raw sequences for Picea glauca | https://spruce-up.ca/ https://www.smartforests.ca |
| Phytozome | Genomic assemblies and raw data for Thuja plicata Plant comparative genomics portal | https://phytozome-next.jgi.doe.gov |
| China National GeneBank Database | Genomic assemblies and raw sequences and annotation for:
| https://db.cngb.org |
| PLAZA | Comparative genomic data for several conifers | https://bioinformatics.psb.ugent.be/plaza/ |
| European nucleotide archive | Genomic assemblies and raw sequences and annotation for Larix decidua and L. kaempferi | https://www.ebi.ac.uk/ena |
| Earth Biogenome projects | About 300 coniferous ongoing genomes with links to raw data and sequencing projects | https://www.earthbiogenome.org |
| 10KP: 10,000 Plant Genomes Project | About 80 coniferous ongoing genomes with links to raw data and sequencing projects | https://db.cngb.org/10kp/ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Traversari, S.; Giovannelli, A.; Emiliani, G. Wood Formation under Changing Environment: Omics Approaches to Elucidate the Mechanisms Driving the Early-to-Latewood Transition in Conifers. Forests 2022, 13, 608. https://doi.org/10.3390/f13040608
Traversari S, Giovannelli A, Emiliani G. Wood Formation under Changing Environment: Omics Approaches to Elucidate the Mechanisms Driving the Early-to-Latewood Transition in Conifers. Forests. 2022; 13(4):608. https://doi.org/10.3390/f13040608
Chicago/Turabian StyleTraversari, Silvia, Alessio Giovannelli, and Giovanni Emiliani. 2022. "Wood Formation under Changing Environment: Omics Approaches to Elucidate the Mechanisms Driving the Early-to-Latewood Transition in Conifers" Forests 13, no. 4: 608. https://doi.org/10.3390/f13040608
APA StyleTraversari, S., Giovannelli, A., & Emiliani, G. (2022). Wood Formation under Changing Environment: Omics Approaches to Elucidate the Mechanisms Driving the Early-to-Latewood Transition in Conifers. Forests, 13(4), 608. https://doi.org/10.3390/f13040608