
Seed Dispersal Models for Natural Regeneration: A Review and Prospects

 , , , , ,
, , , , ,
Abstract
:1. Introduction
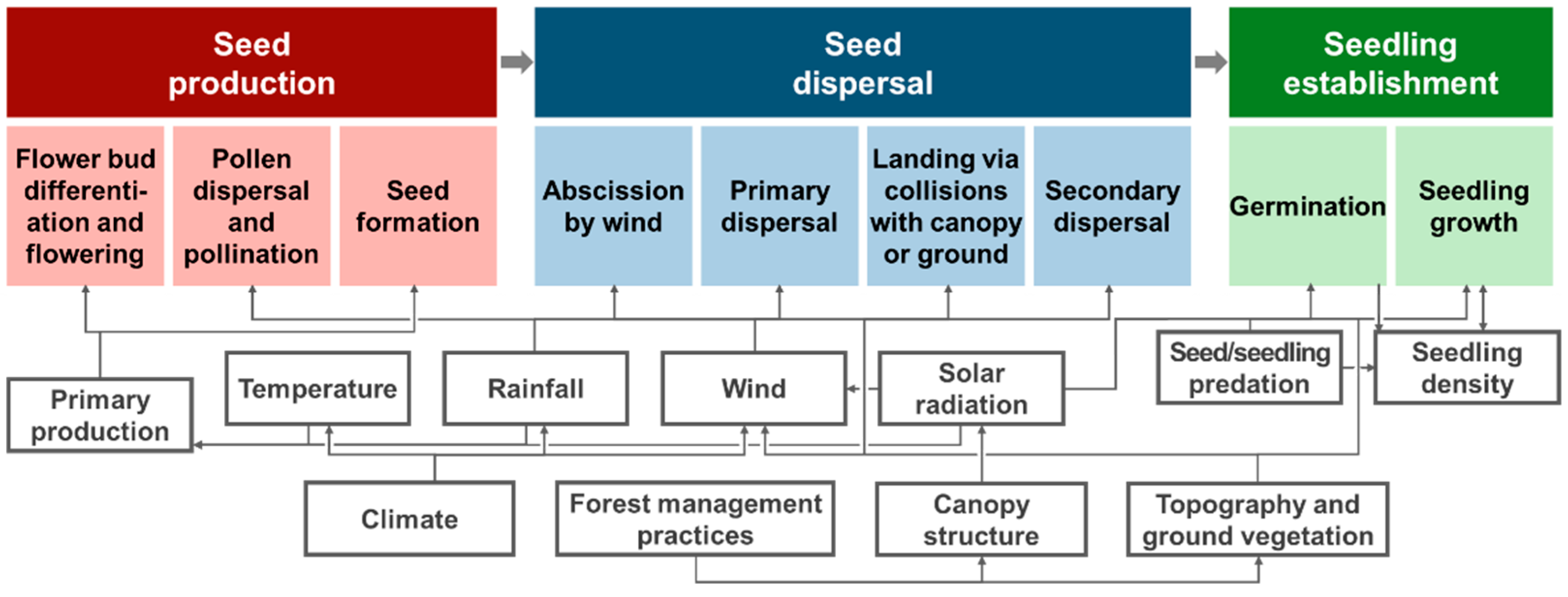
2. Natural Regeneration and Seed Dispersal
3. Key Mechanisms of Seed Dispersal Models
3.1. Simple Empirical and Mechanistic Dispersal Models
3.2. Eulerian and Lagrangian Modeling Approach
3.3. Long-Distance Dispersal
3.4. Challenges in Seed Dispersal Prediction for Natural Regeneration Using the WINDISPER
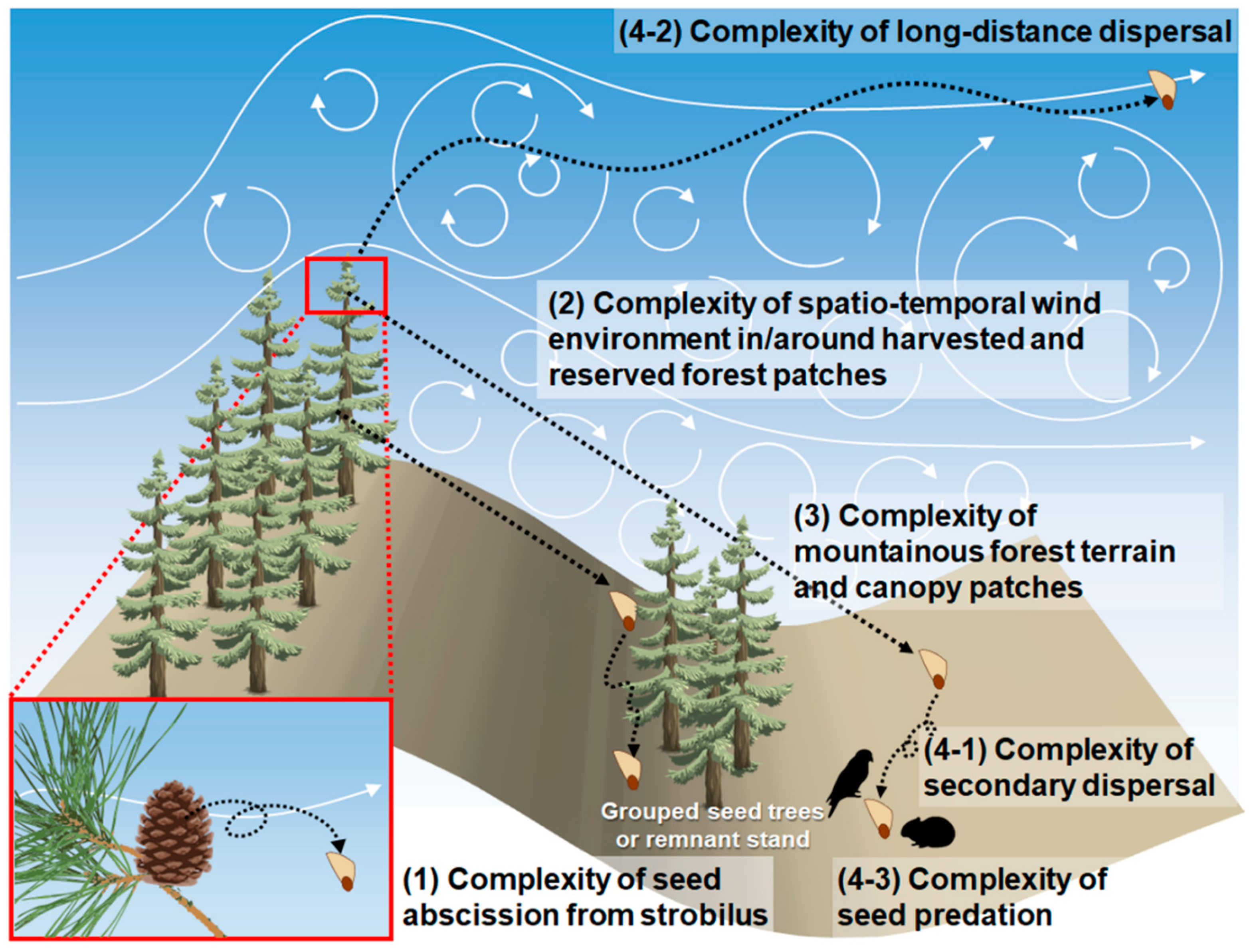
4. Prospects for Seed Dispersal Models Applicable to Natural Regeneration
4.1. Seed Abscission Mechanisms by Wind
4.2. Primary Dispersal by Spatio-Temporally Complex Wind Environment
4.3. Termination of Seed Flight via Collisions with Canopy or Ground
4.4. Secondary Dispersal, LDD, and Consumption by Herbivores
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
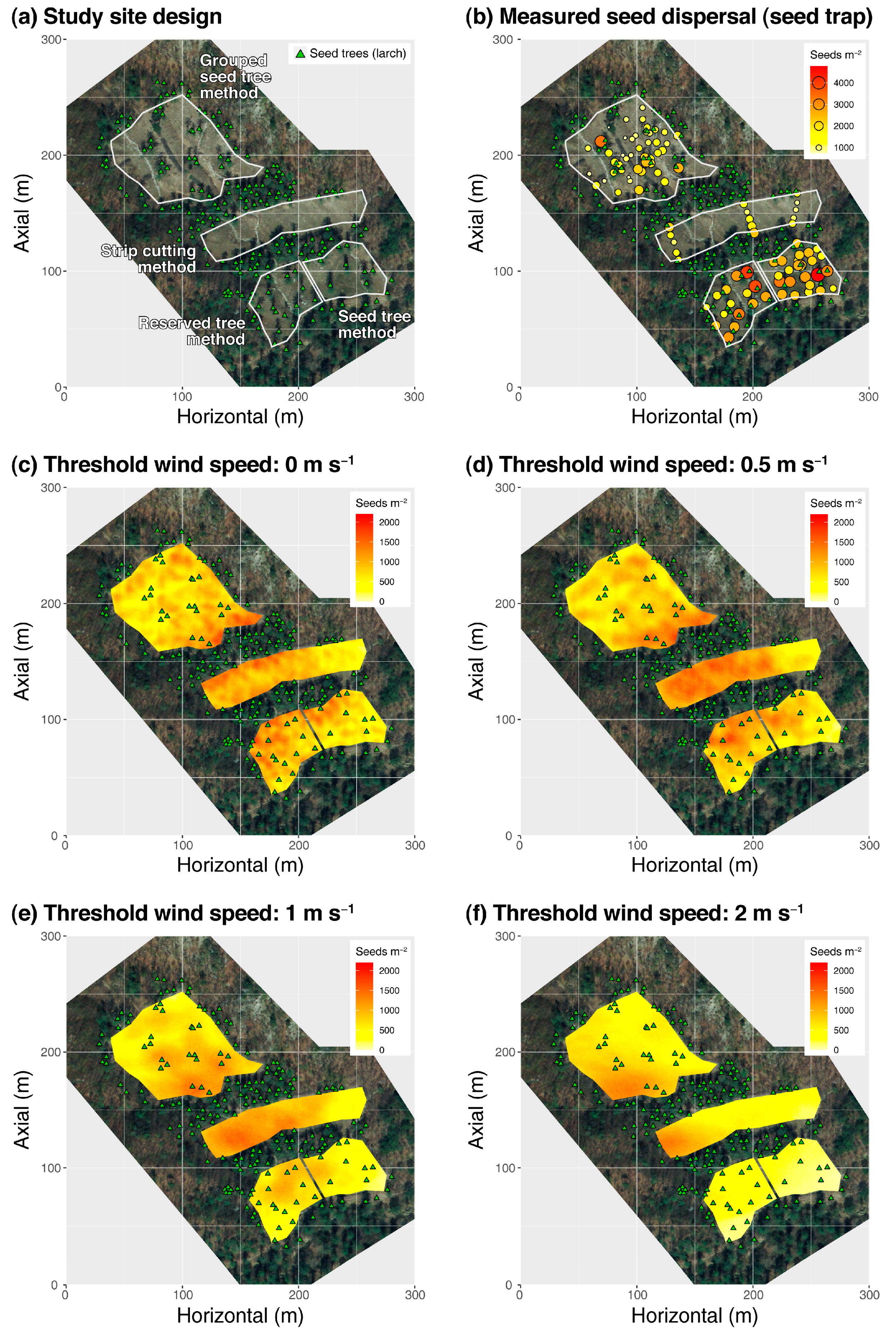
Appendix A. Case of Seed Dispersal Simulation by the WINDISPER
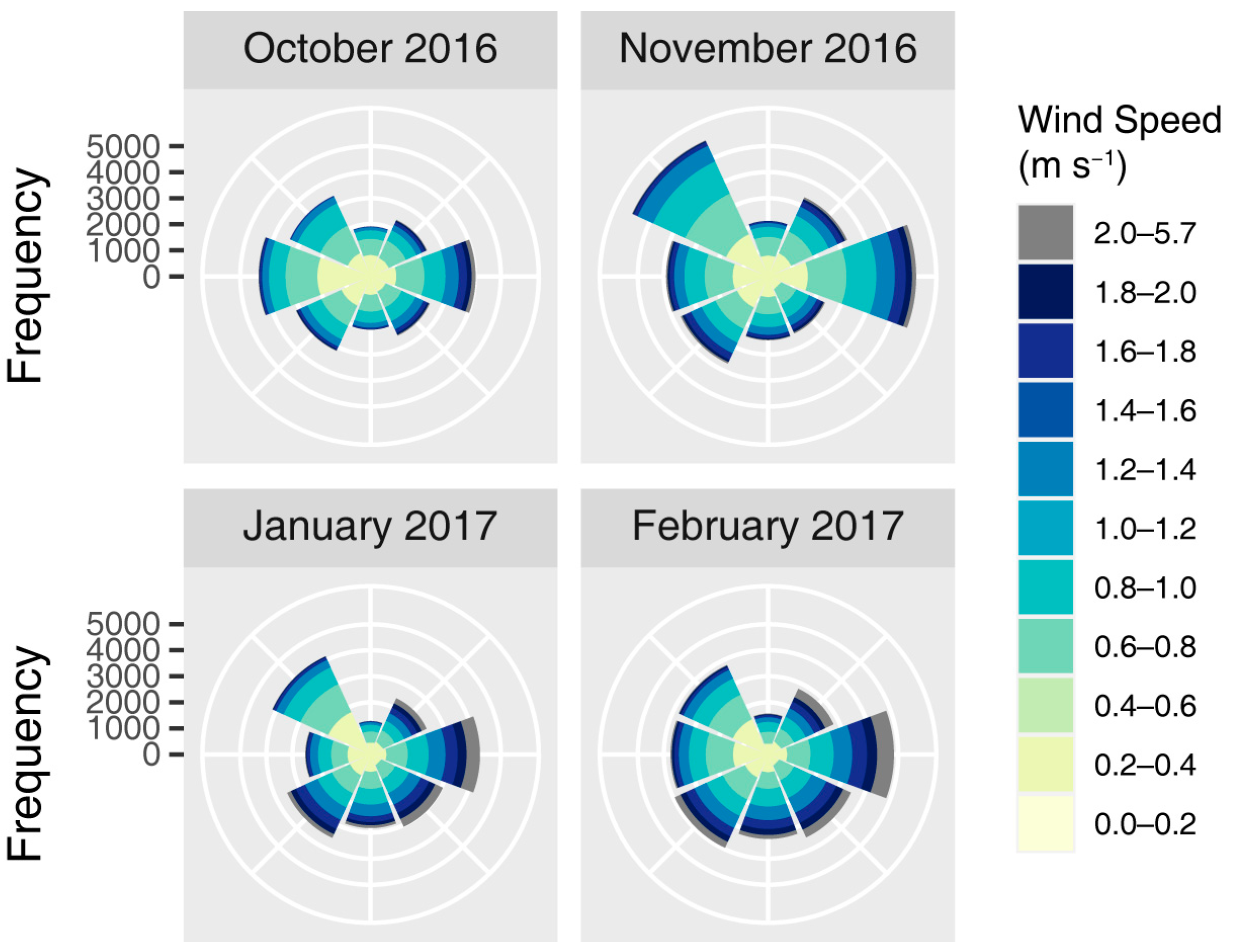
Appendix A.1. Study Site
Appendix A.2. Seed Dispersal Monitoring


Appendix A.3. Modeling Procedures

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wind Direction (°) | 0 m s−1 | 0.5 m s−1 | 1 m s−1 | 2 m s−1 | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | % | Mean | % | Mean | % | Mean | % | |
| 0 | 0.60 | 6.6 | 0.86 | 5.8 | 1.48 | 3.0 | 2.51 | 0.7 |
| 45 | 0.78 | 10.6 | 1.07 | 11.1 | 1.71 | 12.6 | 2.69 | 16.7 |
| 90 | 0.86 | 18.7 | 1.13 | 20.8 | 1.74 | 27.1 | 2.68 | 40.6 |
| 135 | 0.88 | 10.5 | 1.12 | 12.0 | 1.69 | 15.6 | 2.64 | 18.9 |
| 180 | 0.72 | 9.6 | 1.00 | 9.7 | 1.57 | 9.5 | 2.55 | 6.5 |
| 225 | 0.72 | 13.2 | 1.04 | 12.8 | 1.61 | 14.5 | 2.60 | 10.2 |
| 270 | 0.61 | 13.8 | 0.93 | 11.8 | 1.53 | 8.5 | 2.53 | 4.6 |
| 315 | 0.64 | 16.9 | 0.89 | 15.9 | 1.45 | 9.1 | 2.44 | 1.7 |
| Total * | 0.72 | 64.6 | 1.02 | 45.1 | 1.64 | 15.4 | 2.65 | 2.1 |
Appendix A.4. Results of Seed Dispersal Simulation Using the WINDISPER
| 0 m s−1 | 0.5 m s−1 | 1 m s−1 | 2 m s−1 | |||||
|---|---|---|---|---|---|---|---|---|
| R2 | p-Value | R2 | p-Value | R2 | p-Value | R2 | p-Value | |
| Seed tree method | – | 0.793 | – | 0.515 | – | 0.615 | – | 0.981 |
| Grouped seed tree | 0.091 | 0.044 | 0.067 | 0.086 | 0.077 | 0.064 | – | 0.109 |
| Reserved seed tree | – | 0.246 | – | 0.436 | – | 0.567 | 0.357 | 0.019 |
| Strip cutting | – | 0.478 | – | 0.147 | 0.321 | 0.028 | – | 0.525 |
| Global | 0.035 | 0.059 | – | 0.102 | – | 0.636 | 0.038 | 0.050 |
References
- Boydak, M. Silvicultural characteristics and natural regeneration of Pinus brutia Ten.—A review. Plant Ecol. 2004, 171, 153–163. [Google Scholar] [CrossRef]
- Crouzeilles, R.; Beyer, H.L.; Monteiro, L.M.; Feltran-Barbieri, R.; Pessôa, A.C.; Barros, F.S.; Lindenmayer, D.B.; Lino, E.D.; Grelle, C.E.; Chazdon, R.L. Achieving cost-effective landscape-scale forest restoration through targeted natural regeneration. Conserv. Lett. 2020, 13, e12709. [Google Scholar] [CrossRef]
- Crouzeilles, R.; Ferreira, M.S.; Chazdon, R.L.; Lindenmayer, D.B.; Sansevero, J.B.; Monteiro, L.; Iribarrem, A.; Latawiec, A.E.; Strassburg, B.B. Ecological restoration success is higher for natural regeneration than for active restoration in tropical forests. Sci. Adv. 2017, 3, e1701345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gärtner, S.M.; Lieffers, V.J.; Macdonald, S.E. Ecology and management of natural regeneration of white spruce in the boreal forest. Environ. Rev. 2011, 19, 461–478. [Google Scholar] [CrossRef]
- Kohler, M.; Pyttel, P.; Kuehne, C.; Modrow, T.; Bauhus, J. On the knowns and unknowns of natural regeneration of silviculturally managed sessile oak (Quercus petraea (Matt.) Liebl.) forests—A literature review. Ann. For. Sci. 2020, 77, 101. [Google Scholar] [CrossRef]
- Goerlich, D.L.; Nyland, R.D. Natural regeneration of eastern hemlock: A review. In Symposium on Sustainable Management of Hemlock Ecosystems in Eastern North America. Gen. Tech. Rep. NE-267; McManus Katherine, A., Shields Kathleen, S., Souto Dennis, R., Eds.; US Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Newtown Square, PA, USA, 2000; pp. 14–22. [Google Scholar]
- Ashton, M.S.; Kelty, M.J. Chater 5. Ecology of regerneration. In The Practice of Silviculture: Applied Forest Ecology; John Wiley & Sons Ltd.: West Sussex, UK, 2018; pp. 80–116. [Google Scholar]
- Dey, D.C.; Knapp, B.O.; Battaglia, M.A.; Deal, R.L.; Hart, J.L.; O’Hara, K.L.; Schweitzer, C.J.; Schuler, T.M. Barriers to natural regeneration in temperate forests across the USA. New For. 2019, 50, 11–40. [Google Scholar] [CrossRef]
- Kaliszewski, A. Cost analysis of artificial and natural oak regeneration in selected forest districts. For. Res. Pap. 2018, 78, 315–321. [Google Scholar] [CrossRef]
- Minami, S.; Azuma, A. Various flying modes of wind-dispersal seeds. J. Theor. Biol. 2003, 225, 1–14. [Google Scholar] [CrossRef]
- Tan, K.; DONG, S.-P.; Lu, T.; ZHANG, Y.-J.; XU, S.-T.; REN, M.-X. Diversity and evolution of samara in angiosperm. Chin. J. Plant Ecol. 2018, 42, 806. [Google Scholar] [CrossRef] [Green Version]
- Landhäusser, S.M.; Pinno, B.D.; Mock, K.E. Tamm Review: Seedling-based ecology, management, and restoration in aspen (Populus tremuloides). For. Ecol. Manag. 2019, 432, 231–245. [Google Scholar] [CrossRef]
- Zakaria, M.Y.; Dos Santos, C.R.; Dayhoum, A.; Marques, F.; Hajj, M.R. Modeling and prediction of aerodynamic characteristics of free fall rotating wing based on experiments. In Proceedings of the International Conference on Aerospace Sciences and Aviation Technology, Cairo, Egypt, 9–11 April 2019; pp. 1–15. [Google Scholar]
- Lentink, D.; Dickson, W.B.; Van Leeuwen, J.L.; Dickinson, M.H. Leading-edge vortices elevate lift of autorotating plant seeds. Science 2009, 324, 1438–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaproth, M.A.; McGraw, J.B. Seed viability and dispersal of the wind-dispersed invasive Ailanthus altissima in aqueous environments. For. Sci. 2008, 54, 490–496. [Google Scholar] [CrossRef]
- Zhu, J.; Liu, M.; Xin, Z.; Liu, Z.; Schurr, F.M. A trade-off between primary and secondary seed dispersal by wind. Plant Ecol. 2019, 220, 541–552. [Google Scholar] [CrossRef]
- Carlo, T.A.; García, D.; Martínez, D.; Gleditsch, J.M.; Morales, J.M. Where do seeds go when they go far? Distance and directionality of avian seed dispersal in heterogeneous landscapes. Ecology 2013, 94, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Montoya, D.; Zavala, M.A.; Rodríguez, M.A.; Purves, D.W. Animal versus wind dispersal and the robustness of tree species to deforestation. Science 2008, 320, 1502–1504. [Google Scholar] [CrossRef] [Green Version]
- Thomson, F.J.; Moles, A.T.; Auld, T.D.; Kingsford, R.T. Seed dispersal distance is more strongly correlated with plant height than with seed mass. J. Ecol. 2011, 99, 1299–1307. [Google Scholar] [CrossRef]
- Harmer, R. Natural regeneration of broadleaved trees in Britain: II Seed production and predation. Forestry 1994, 67, 275–286. [Google Scholar] [CrossRef]
- Barna, M. The effects of cutting regimes on natural regeneration in submountain beech forests: Species diversity and abundance. J. For. Sci. 2008, 54, 533–544. [Google Scholar] [CrossRef] [Green Version]
- Stoehr, M.U. Seed production of western larch in seed-tree systems in the southern interior of British Columbia. For. Ecol. Manag. 2000, 130, 7–15. [Google Scholar] [CrossRef]
- Kuparinen, A. Mechanistic models for wind dispersal. Trends Plant Sci. 2006, 11, 296–301. [Google Scholar] [CrossRef]
- Gregory, P. Interpreting plant disease dispersal gradients. Annu. Rev. Phytopathol. 1968, 6, 189–212. [Google Scholar] [CrossRef]
- Clark, J.S.; Silman, M.; Kern, R.; Macklin, E.; HilleRisLambers, J. Seed dispersal near and far: Patterns across temperate and tropical forests. Ecology 1999, 80, 1475–1494. [Google Scholar] [CrossRef]
- Frampton, V.L. The spread of virus diseases of the yellows type under field conditions. Phytopathology 1942, 32, 799–808. [Google Scholar]
- Katul, G.; Porporato, A.; Nathan, R.; Siqueira, M.; Soons, M.; Poggi, D.; Horn, H.; Levin, S. Mechanistic analytical models for long-distance seed dispersal by wind. Am. Nat. 2005, 166, 368–381. [Google Scholar] [CrossRef] [PubMed]
- Portnoy, S.; Willson, M.F. Seed dispersal curves: Behavior of the tail of the distribution. Evol. Ecol. 1993, 7, 25–44. [Google Scholar] [CrossRef]
- Nathan, R.; Safriel, U.N.; Noy-Meir, I. Field validation and sensitivity analysis of a mechanistic model for tree seed dispersal by wind. Ecology 2001, 82, 374–388. [Google Scholar] [CrossRef]
- Kruse, S.; Gerdes, A.; Kath, N.J.; Herzschuh, U. Implementing spatially explicit wind-driven seed and pollen dispersal in the individual-based larch simulation model: LAVESI-WIND 1.0. Geosci. Model Dev. 2018, 11, 4451–4467. [Google Scholar] [CrossRef] [Green Version]
- Okubo, A.; Levin, S.A. A theoretical framework for data analysis of wind dispersal of seeds and pollen. Ecology 1989, 70, 329–338. [Google Scholar] [CrossRef]
- Tackenberg, O. Modeling long-distance dispersal of plant diaspores by wind. Ecol. Monogr. 2003, 73, 173–189. [Google Scholar] [CrossRef]
- Horn, S.; Raabe, A.; Will, H.; Tackenberg, O. TurbSeed—a model for wind dispersal of seeds in turbulent currents based on publicly available climate data. Ecol. Model. 2012, 237, 1–10. [Google Scholar] [CrossRef]
- Loos, C.; Seppelt, R.; Meier-Bethke, S.; Schiemann, J.; Richter, O. Spatially explicit modelling of transgenic maize pollen dispersal and cross-pollination. J. Theor. Biol. 2003, 225, 241–255. [Google Scholar] [CrossRef]
- Aylor, D.E.; Flesch, T.K. Estimating spore release rates using a Lagrangian stochastic simulation model. J. Appl. Meteorol. Climatol. 2001, 40, 1196–1208. [Google Scholar] [CrossRef]
- Bohrer, G.; Katul, G.G.; Walko, R.L.; Avissar, R. Exploring the effects of microscale structural heterogeneity of forest canopies using large-eddy simulations. Bound.-Layer Meteorol. 2009, 132, 351–382. [Google Scholar] [CrossRef]
- Di-Giovanni, F.; Beckett, P. On the mathematical modeling of pollen dispersal and deposition. J. Appl. Meteorol. Climatol. 1990, 29, 1352–1357. [Google Scholar] [CrossRef] [Green Version]
- Maurer, K.D.; Bohrer, G.; Medvigy, D.; Wright, S.J. The timing of abscission affects dispersal distance in a wind-dispersed tropical tree. Funct. Ecol. 2013, 27, 208–218. [Google Scholar] [CrossRef]
- Nathan, R.; Katul, G.G.; Bohrer, G.; Kuparinen, A.; Soons, M.B.; Thompson, S.E.; Trakhtenbrot, A.; Horn, H.S. Mechanistic models of seed dispersal by wind. Theor. Ecol. 2011, 4, 113–132. [Google Scholar] [CrossRef]
- Thompson, S.E.; Assouline, S.; Chen, L.; Trahktenbrot, A.; Svoray, T.; Katul, G.G. Secondary dispersal driven by overland flow in drylands: Review and mechanistic model development. Mov. Ecol. 2014, 2, 7. [Google Scholar] [CrossRef] [Green Version]
- Treep, J.; de Jager, M.; Kuiper, L.S.; Duman, T.; Katul, G.G.; Soons, M.B. Costs and benefits of non-random seed release for long-distance dispersal in wind-dispersed plant species. Oikos 2018, 127, 1330–1343. [Google Scholar] [CrossRef] [Green Version]
- Treep, J.; de Jager, M.; Bartumeus, F.; Soons, M.B. Seed dispersal as a search strategy: Dynamic and fragmented landscapes select for multi-scale movement strategies in plants. Mov. Ecol. 2021, 9, 4. [Google Scholar] [CrossRef]
- Williams, C.G.; LaDeau, S.L.; Oren, R.; Katul, G.G. Modeling seed dispersal distances: Implications for transgenic Pinus taeda. Ecol. Appl. 2006, 16, 117–124. [Google Scholar] [CrossRef]
- Wright, S.J.; Trakhtenbrot, A.; Bohrer, G.; Detto, M.; Katul, G.G.; Horvitz, N.; Muller-Landau, H.C.; Jones, F.A.; Nathan, R. Understanding strategies for seed dispersal by wind under contrasting atmospheric conditions. PNAS 2008, 105, 19084–19089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiyosawa, S.; Shiyomi, M. A theoretical evaluation of the effect of mixing resistant variety with susceptible variety for controlling plant diseases. JJP 1972, 38, 41–51. [Google Scholar] [CrossRef]
- McCartney, H.; Bainbridge, A. Deposition gradients near to a point source in a barley crop. J. Phytopathol. 1984, 109, 219–236. [Google Scholar] [CrossRef]
- Ribbens, E.; Silander, J.A., Jr.; Pacala, S.W. Seedling recruitment in forests: Calibrating models to predict patterns of tree seedling dispersion. Ecology 1994, 75, 1794–1806. [Google Scholar] [CrossRef]
- Stoyon, D.; Wagner, S. Estimating the fruit dispersion of anemochorous trees. Ecol. Model. 2001, 145, 35–47. [Google Scholar] [CrossRef]
- Skarpaas, O.; Stabbetorp, O.E.; Rønning, I.; Svennungsen, T.O. How far can a hawk’s beard fly? Measuring and modelling the dispersal of Crepis praemorsa. J. Ecol. 2004, 92, 747–757. [Google Scholar] [CrossRef]
- Morales, J.M.; López, T.M. Mechanistic models of seed dispersal by animals. Oikos 2022. [Google Scholar] [CrossRef]
- Wichmann, M.C.; Alexander, M.J.; Soons, M.B.; Galsworthy, S.; Dunne, L.; Gould, R.; Fairfax, M.; Niggemann, M.; Hails, R.S.; Bullock, J.M. Human-mediated dispersal of seeds over long distances. Proc. R. Soc. B: Biol. Sci. 2009, 276, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Greene, D.F.; Canham, C.D.; Coates, K.D.; Lepage, P.T. An evaluation of alternative dispersal functions for trees. J. Ecol. 2004, 92, 758–766. [Google Scholar] [CrossRef]
- LePage, P.T.; Canham, C.D.; Coates, K.D.; Bartemucci, P. Seed abundance versus substrate limitation of seedling recruitment in northern temperate forests of British Columbia. Can. J. For. Res. 2000, 30, 415–427. [Google Scholar] [CrossRef]
- Soons, M.B.; Bullock, J.M. Non-random seed abscission, long-distance wind dispersal and plant migration rates. J. Ecol. 2008, 96, 581–590. [Google Scholar] [CrossRef]
- Greene, D.; Johnson, E. A model of wind dispersal of winged or plumed seeds. Ecology 1989, 70, 339–347. [Google Scholar] [CrossRef]
- Levin, S.A.; Muller-Landau, H.C.; Nathan, R.; Chave, J. The ecology and evolution of seed dispersal: A theoretical perspective. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 575–604. [Google Scholar] [CrossRef]
- Nathan, R.; Getz, W.M.; Revilla, E.; Holyoak, M.; Kadmon, R.; Saltz, D.; Smouse, P.E. A movement ecology paradigm for unifying organismal movement research. Proc. Natl. Acad. Sci. USA 2008, 105, 19052–19059. [Google Scholar] [CrossRef] [Green Version]
- Nuttle, T.; Haefner, J.W. Seed dispersal in heterogeneous environments: Bridging the gap between mechanistic dispersal and forest dynamics models. Am. Nat. 2005, 165, 336–349. [Google Scholar] [CrossRef] [PubMed]
- Axer, M.; Schlicht, R.; Wagner, S. Modelling potential density of natural regeneration of European oak species (Quercus robur L., Quercus petraea (Matt.) Liebl.) depending on the distance to the potential seed source: Methodological approach for modelling dispersal from inventory data at forest enterprise level. For. Ecol. Manag. 2021, 482, 118802. [Google Scholar] [CrossRef]
- de Andrés, E.G.; Camarero, J.J.; Martínez, I.; Coll, L. Uncoupled spatiotemporal patterns of seed dispersal and regeneration in Pyrenean silver fir populations. For. Ecol. Manag. 2014, 319, 18–28. [Google Scholar] [CrossRef]
- Holmström, E.; Karlsson, M.; Nilsson, U. Modeling birch seed supply and seedling establishment during forest regeneration. Ecol. Model. 2017, 352, 31–39. [Google Scholar] [CrossRef]
- Manso, R.; Pardos, M.; Keyes, C.R.; Calama, R. Modelling the spatio-temporal pattern of primary dispersal in stone pine (Pinus pinea L.) stands in the Northern Plateau (Spain). Ecol. Model. 2012, 226, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Katul, G.G.; Albertson, J.D. Modeling CO2 sources, sinks, and fluxes within a forest canopy. J. Geophys. Res. Atmos. 1999, 104, 6081–6091. [Google Scholar] [CrossRef] [Green Version]
- Stockie, J.M. The mathematics of atmospheric dispersion modeling. Siam Rev. 2011, 53, 349–372. [Google Scholar] [CrossRef]
- Di-Giovanni, F.; Beckett, P.; Flenley, J. Modelling of dispersion and deposition of tree pollen within a forest canopy. Grana 1989, 28, 129–139. [Google Scholar] [CrossRef]
- McCartney, H.; Lacey, M.E. Wind dispersal of pollen from crops of oilseed rape (Brassica napus L.). J. Aerosol. Sci. 1991, 22, 467–477. [Google Scholar] [CrossRef]
- Andersen, M. Mechanistic models for the seed shadows of wind-dispersed plants. Am. Nat. 1991, 137, 476–497. [Google Scholar] [CrossRef]
- Soons, M.B.; Heil, G.W.; Nathan, R.; Katul, G.G. Determinants of long-distance seed dispersal by wind in grasslands. Ecology 2004, 85, 3056–3068. [Google Scholar] [CrossRef] [Green Version]
- Jarosz, N.; Loubet, B.; Huber, L. Modelling airborne concentration and deposition rate of maize pollen. Atmos. Environ. 2004, 38, 5555–5566. [Google Scholar] [CrossRef] [Green Version]
- Nathan, R.; Katul, G.G.; Horn, H.S.; Thomas, S.M.; Oren, R.; Avissar, R.; Pacala, S.W.; Levin, S.A. Mechanisms of long-distance dispersal of seeds by wind. Nature 2002, 418, 409–413. [Google Scholar] [CrossRef]
- Boehm, M.T.; Aylor, D.E. Lagrangian stochastic modeling of heavy particle transport in the convective boundary layer. Atmos. Environ. 2005, 39, 4841–4850. [Google Scholar] [CrossRef]
- Poggi, D.; Katul, G.; Albertson, J. Scalar dispersion within a model canopy: Measurements and three-dimensional Lagrangian models. Adv. Water Resour. 2006, 29, 326–335. [Google Scholar] [CrossRef]
- Nathan, R.; Katul, G.G. Foliage shedding in deciduous forests lifts up long-distance seed dispersal by wind. Proc. Natl. Acad. Sci. USA 2005, 102, 8251–8256. [Google Scholar] [CrossRef] [Green Version]
- Trakhtenbrot, A.; Katul, G.; Nathan, R. Mechanistic modeling of seed dispersal by wind over hilly terrain. Ecol. Model. 2014, 274, 29–40. [Google Scholar] [CrossRef]
- Bullock, J.M.; Clarke, R.T. Long distance seed dispersal by wind: Measuring and modelling the tail of the curve. Oecologia 2000, 124, 506–521. [Google Scholar] [CrossRef] [PubMed]
- Dauer, J.T.; Mortensen, D.A.; Vangessel, M.J. Temporal and spatial dynamics of long-distance Conyza canadensis seed dispersal. J. Appl. Ecol. 2007, 44, 105–114. [Google Scholar] [CrossRef]
- Levey, D.J.; Tewksbury, J.J.; Bolker, B.M. Modelling long-distance seed dispersal in heterogeneous landscapes. J. Ecol. 2008, 96, 599–608. [Google Scholar] [CrossRef] [Green Version]
- Cain, M.L.; Milligan, B.G.; Strand, A.E. Long-distance seed dispersal in plant populations. Am. J. Bot. 2000, 87, 1217–1227. [Google Scholar] [CrossRef] [Green Version]
- Kot, M.; Lewis, M.A.; van den Driessche, P. Dispersal data and the spread of invading organisms. Ecology 1996, 77, 2027–2042. [Google Scholar] [CrossRef]
- Ouborg, N.; Piquot, Y.; Van Groenendael, J. Population genetics, molecular markers and the study of dispersal in plants. J. Ecol. 1999, 87, 551–568. [Google Scholar] [CrossRef]
- Turchin, P. Quantitative Analysis of Movement: Measuring and Modeling Population Redistribution in Animals and Plants; Sinauer Associates: Sunderland, MA, USA, 1998. [Google Scholar]
- Webster, M.S.; Marra, P.P.; Haig, S.M.; Bensch, S.; Holmes, R.T. Links between worlds: Unraveling migratory connectivity. Trends Ecol. Evol. 2002, 17, 76–83. [Google Scholar] [CrossRef]
- Nathan, R.; Perry, G.; Cronin, J.T.; Strand, A.E.; Cain, M.L. Methods for estimating long-distance dispersal. Oikos 2003, 103, 261–273. [Google Scholar] [CrossRef] [Green Version]
- National Geographic Information Institute. The National Atlas of Korea II.; National Geographic Information Institute, Gyeonggi–do 16517, Korea. Available online: http://nationalatlas.ngii.go.kr/ (accessed on 20 August 2021).
- Katul, G.; Poggi, D. The effects of gentle topographic variation on dispersal kernels of inertial particles. Geophys. Res. Lett. 2012, 39. [Google Scholar] [CrossRef] [Green Version]
- Wyse, S.V.; Hulme, P.E.; Holland, E.P. Partitioning intraspecific variation in seed dispersal potential using a low-cost method for rapid estimation of samara terminal velocity. Methods Ecol. Evol. 2019, 10, 1298–1307. [Google Scholar] [CrossRef]
- Pazos, G.E.; Greene, D.F.; Katul, G.; Bertiller, M.B.; Soons, M.B. Seed dispersal by wind: Towards a conceptual framework of seed abscission and its contribution to long-distance dispersal. J. Ecol. 2013, 101, 889–904. [Google Scholar] [CrossRef]
- Schippers, P.; Jongejans, E. Release thresholds strongly determine the range of seed dispersal by wind. Ecol. Model. 2005, 185, 93–103. [Google Scholar] [CrossRef]
- Darling, E.; Samis, K.E.; Eckert, C.G. Increased seed dispersal potential towards geographic range limits in a Pacific coast dune plant. New Phytol. 2008, 178, 424–435. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, B.; González, E. Pulses of seed release in riparian Salicaceae coincide with high atmospheric temperature. River Res. Appl. 2019, 35, 1590–1596. [Google Scholar] [CrossRef]
- Murray, B.R.; Brown, A.; Dickman, C.; Crowther, M. Geographical gradients in seed mass in relation to climate. J. Biogeogr. 2004, 31, 379–388. [Google Scholar] [CrossRef]
- Niklas, K.J. The aerodynamics of wind pollination. Bot. Rev. 1985, 51, 328–386. [Google Scholar] [CrossRef]
- Dawson, C.; Vincent, J.F.; Rocca, A.-M. How pine cones open. Nature 1997, 390, 668. [Google Scholar] [CrossRef]
- Song, K.; Chang, S.S.; Lee, S.J. How the pine seeds attach to/detach from the pine cone scale? Front. Life Sci. 2017, 10, 38–47. [Google Scholar] [CrossRef]
- Greene, D.F. The role of abscission in long-distance seed dispersal by the wind. Ecology 2005, 86, 3105–3110. [Google Scholar] [CrossRef]
- Endalew, A.M.; Hertog, M.; Delele, M.; Baetens, K.; Persoons, T.; Baelmans, M.; Ramon, H.; Nicolaï, B.; Verboven, P. CFD modelling and wind tunnel validation of airflow through plant canopies using 3D canopy architecture. Int. J. Heat Fluid Flow 2009, 30, 356–368. [Google Scholar] [CrossRef]
- Lee, J.P.; Lee, S.J. PIV analysis on the shelter effect of a bank of real fir trees. J. Wind Eng. Ind. Aerodyn. 2012, 110, 40–49. [Google Scholar] [CrossRef]
- Clark, T.L.; Mitchell, S.J. Three-dimensional simulations of air flow and momentum transfer in partially harvested forests. Bound. Layer Meteorol. 2007, 125, 505–524. [Google Scholar] [CrossRef]
- Novak, M.D.; Warland, J.S.; Orchansky, A.L.; Ketler, R.; Green, S. Wind tunnel and field measurements of turbulent flow in forests. Part I: Uniformly thinned stands. Bound. Layer Meteorol. 2000, 95, 457–495. [Google Scholar] [CrossRef]
- Daikoku, K.; Hattori, S.; Deguchi, A.; Fujita, Y.; Park, H.; Matsumoto, K. Impact of wind direction on diurnal and seasonal changes in wind profiles. J. For. Res. 2007, 12, 452–466. [Google Scholar] [CrossRef]
- Ha, T.; Lee, I.-b.; Kwon, K.-s.; Lee, S.-J. Development of a micro-scale CFD model to predict wind environment on mountainous terrain. Comput. Electron. Agric. 2018, 149, 110–120. [Google Scholar] [CrossRef]
- Domínguez-Brito, A.C.; Cabrera-Gámez, J.; Viera-Pérez, M.; Rodríguez-Barrera, E.; Hernández-Calvento, L. A DIY low-cost wireless wind data acquisition system used to study an arid coastal foredune. Sensors 2020, 20, 1064. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, K.; Inoue, M.; Shimura, T.; Iguchi, M. In Situ, Rotor-Based Drone measurement of wind vector and aerosol concentration in volcanic areas. Atmosphere 2021, 12, 376. [Google Scholar] [CrossRef]
- Thielicke, W.; Hübert, W.; Müller, U.; Eggert, M.; Wilhelm, P. Towards accurate and practical drone-based wind measurements with an ultrasonic anemometer. Atmos. Meas. Tech. 2021, 14, 1303–1318. [Google Scholar] [CrossRef]
- Yoon, S.; Jang, K.; Won, M. The spatial distribution characteristics of Automatic Weather Stations in the mountainous area over South Korea. Korean J. Agric. For. Meteorol. 2018, 20, 117–126. [Google Scholar] [CrossRef]
- Pounden, E.; Greene, D.; Quesada, M.; Contreras Sánchez, J. The effect of collisions with vegetation elements on the dispersal of winged and plumed seeds. J. Ecol. 2008, 96, 591–598. [Google Scholar] [CrossRef]
- Bohrer, G.; Katul, G.G.; Nathan, R.; Walko, R.L.; Avissar, R. Effects of canopy heterogeneity, seed abscission and inertia on wind-driven dispersal kernels of tree seeds. J. Ecol. 2008, 96, 569–580. [Google Scholar] [CrossRef]
- Esmail, S.; Agrawal, P.; Aly, S. A novel analytical approach for advection diffusion equation for radionuclide release from an area source. Nucl. Eng. Technol. 2020, 52, 819–826. [Google Scholar] [CrossRef]
- Jeong, S.H.; Lee, S.H. Effects of windbreak Forest according to tree species and planting methods based on wind tunnel experiments. For. Sci. Technol. 2020, 16, 188–194. [Google Scholar] [CrossRef]
- Vollsinger, S.; Mitchell, S.J.; Byrne, K.E.; Novak, M.D.; Rudnicki, M. Wind tunnel measurements of crown streamlining and drag relationships for several hardwood species. Can. J. For. Res. 2005, 35, 1238–1249. [Google Scholar] [CrossRef]
- der Weduwen, D.; Ruxton, G.D. Secondary dispersal mechanisms of winged seeds: A review. Biol. Rev. 2019, 94, 1830–1838. [Google Scholar] [CrossRef] [PubMed]
- Schurr, F.M.; Bond, W.J.; Midgley, G.F.; Higgins, S.I. A mechanistic model for secondary seed dispersal by wind and its experimental validation. J. Ecol. 2005, 93, 1017–1028. [Google Scholar] [CrossRef]
- Higgins, S.I.; Nathan, R.; Cain, M.L. Are long-distance dispersal events in plants usually caused by nonstandard means of dispersal? Ecology 2003, 84, 1945–1956. [Google Scholar] [CrossRef]
- Fornara, D.; Dalling, J.W. Post-dispersal removal of seeds of pioneer species from five Panamanian forests. J. Trop. Ecol. 2005, 21, 79–84. [Google Scholar] [CrossRef]
- Kim, G.-T.; Kim, H.-J. Secondary dispersion of several broadleaved tree seeds by wildlife in Mt. Jungwang, Pyeongchang-gun, Korea. Korean J. Environ. Ecol. 2013, 27, 64–70. [Google Scholar]
- Vander Wall, S.B.; Joyner, J.W. Secondary dispersal by the wind of winged pine seeds across the ground surface. Am. Midl. Nat. 1998, 139, 365–373. [Google Scholar] [CrossRef]
- Von der Lippe, M.; Bullock, J.M.; Kowarik, I.; Knopp, T.; Wichmann, M. Human-mediated dispersal of seeds by the airflow of vehicles. PLoS ONE 2013, 8, e52733. [Google Scholar] [CrossRef]
- Liang, W.; Liu, Z.; Liu, M.; Qin, X.; Baskin, C.C.; Baskin, J.M.; Xin, Z.; Wang, Z.; Su, Z.; Zhou, Q. Wing loading, not terminal velocity, is the best parameter to predict capacity of diaspores for secondary wind dispersal. J. Exp. Bot. 2020, 71, 4298–4307. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Madrigal, J.; Candel-Pérez, D.; Jiménez, E.; Moya, D.; de las Heras, J.; Guijarro, M.; Vega, J.A.; Fernández, C.; Hernando, C. Effects of prescribed burning, vegetation treatment and seed predation on natural regeneration of Spanish black pine (Pinus nigra Arn. ssp. salzmannii) in pure and mixed forest stands. For. Ecol. Manag. 2016, 378, 24–30. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Silva-Santos, P.; Fonseca, T.; López-Serrano, F.R.; Tiscar, P.A.; Martínez-García, E.; Andrés, M.; Del Cerro, A. Modelling Spanish black pine postdispersal seed predation in Central-eastern Spain. For. Syst. 2010, 19, 393–403. [Google Scholar] [CrossRef] [Green Version]
- Worthy, F.R.; Law, R.; Hulme, P.E. Modelling the quantitative effects of pre-and post-dispersal seed predation in Pinus sylvestris L. J. Ecol. 2006, 94, 1201–1213. [Google Scholar] [CrossRef]
- Castro, J.; Gómez, J.M.; García, D.; Zamora, R.; Hódar, J.A. Seed predation and dispersal in relict Scots pine forests in southern Spain. Plant Ecol. 1999, 145, 115–123. [Google Scholar] [CrossRef]
- Manso, R.; Pardos, M.; Calama, R. Climatic factors control rodent seed predation in Pinus pinea L. stands in Central Spain. Ann. For. Sci. 2014, 71, 873–883. [Google Scholar] [CrossRef] [Green Version]
- Ruano, I.; Del Peso, C.; Bravo, F. Post-dispersal predation of Pinus pinaster Aiton seeds: Key factors and effects on belowground seed bank. Eur. J. For. Res. 2015, 134, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Bae, J.S.; Joo, R.W.; Kim, Y.-S. Forest transition in South Korea: Reality, path and drivers. Land Use Policy 2012, 29, 198–207. [Google Scholar] [CrossRef]
- Kim, M.; Kraxner, F.; Son, Y.; Jeon, S.W.; Shvidenko, A.; Schepaschenko, D.; Ham, B.-Y.; Lim, C.-H.; Song, C.; Hong, M. Quantifying impacts of national-scale afforestation on carbon budgets in South Korea from 1961 to 2014. Forests 2019, 10, 579. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Yoon, T.; Han, S.; Kim, S.; Yi, M.; Park, G.; Kim, C.; Son, Y.; Kim, R.; Son, Y. Estimating the carbon dynamics of South Korean forests from 1954 to 2012. Biogeosciences 2014, 11, 4637–4650. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Lee, J.Y.; Song, M. Scientific activities responsible for successful forest greening in Korea. For. Sci. Technol. 2017, 13, 1–8. [Google Scholar] [CrossRef] [Green Version]
- You, J.W.; Won, H.K.; Han, H.; Kim, H.S. A comparative analysis of management efficiency between natural regeneration and plantation. Korean J. For. Econ. 2020, 27, 85–91. [Google Scholar] [CrossRef]
- Bae, S.W. Structure and tending method for naturally regenerated young Pinus densiflora Sieb. et Zucc. stands. J. Korean Soc. For. Sci. 1994, 83, 50–62. [Google Scholar]
- Kim, J.H.; Yang, H.M.; Jin, G.Z. The pattern of natural regeneration by three different silvicultural systems in a natural deciduous forest. J. Korean For. Soc. 1999, 88, 169–178. [Google Scholar]
- Na, S.J.; Woo, K.S.; Kim, C.S.; Yoon, J.H.; Lee, H.H.; Lee, D.H. Above-ground growth characteristics between naturally regenerated and planted stands of Pinus densiflora for. erecta Uyeki in Gangwon province. J. Korean Soc. For. Sci. 2010, 99, 323–330. [Google Scholar]
- Chung, J.; Kim, H.S.; Chung, S. Seed fructification and seed dispersal characteristics by harvest types in Natural Regeneration Sites of Larix kaempferi. J. Korean Soc. For. Sci. 2021, 110, 295–307. [Google Scholar] [CrossRef]
- Kim, M.; Kraxner, F.; Forsell, N.; Song, C.; Lee, W.-K. Enhancing the provisioning of ecosystem services in South Korea under climate change: The benefits and pitfalls of current forest management strategies. Reg. Environ. Chang. 2021, 21, 6. [Google Scholar] [CrossRef]
- Korea Forest Service. Statistical Yearbook of Forestry; Korea Forest Service: Daejon, Korea, 2021. (In Korean) [Google Scholar]
- Chung, J.; Kim, H. Occurrence and growth characteristics of natural seedlings by harvest type in a Larix kaempferi Forest. J. Korean For. Soc. 2020, 109, 400–412. [Google Scholar] [CrossRef]
- Liu, M.; Xin, Z.; Su, Z.; Zhao, Y.; Li, X.; Liu, Z.; Cony, M.A.; Liang, W.; Qin, X.; Qian, J. A video camera recording method for measuring terminal velocity of seed dispersal by wind. J. For. Res. 2021, 32, 81–90. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing (R Version 4.0. 3, R Foundation for Statistical Computing, Vienna, Austria, 2020). 2021. Available online: https://www.R-project.org/ (accessed on 17 September 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation Project. Available online: http://qgis.osgeo.org (accessed on 24 November 2021).



| Model Type | Characteristic | Basic Equation and Key Input Variables | References |
|---|---|---|---|
| Empirical models | Typical examples: Power law or exponential models, The 2Dt (bivariate Student’s t distribution) model Strengths: (1) simple and easy to obtain data for fitting the model; (2) effective in approximating the spatial pattern of seed dispersal; (3) widely used in various research Weaknesses: (1) do not reflect physical mechanisms of seed dispersal; (2) require site-specific seed dispersal data | Source: Gregory [24] Source: Clark et al. [25] Tree-dependent variables:dispersal distances, some parameters for exponential curve, seed release height, seed production rate, parameters for bivariate kernels, fall velocities, etc. Environmental variables:none or wind speed | Clark et al. [25]; Frampton et al. [26]; Gregory [24]; Katul et al. [27]; Portnoy and Willson [28] |
| Mechanistic models | Typical examples: Ballistic model, long-distance dispersal (LDD) model Strengths: (1) realistic representation of dispersal processes; (2) more accurate description of LDD Weakness: atmospheric diffusion is not applied Adapted in the WINDISPER (Nathan et al. [29]) and the LAVESI-WIND (Kruse et al. [30]) | Source: Okubo and Levin [31] Tree-dependent variables: mass of seed, falling velocity of seed, height of seed release, etc. Environmental variables:mean wind speed; modal dispersal distance, etc. | Kruse et al. [30]; Nathan et al. [29]; Okubo and Levin [31] |
| Eulerian and Lagrangian model | Typical examples: Gaussian Plume model, Coupled Eulerian–Lagrangian closure models, (CELC), Large-eddy simulations (LES) model Strengths: (1) predict seed dispersal based on the motions of eddies; (2) high accuracy and more realistic representation of seed dispersal; (3) useful to describe ecological phenomena associated with seed dispersal; (4) account for the effect of the canopy structure of the seed trees; (5) simulate seed dispersal pattern for the individual seed Weakness: might be too complicated for the broader use across regions Adapted by PAPPUS (Tackenberg et al. [32]) and TurbSeed (Horn et al. [33]) | Source: Loos et al. 2003 [34] Source: Aylor and Flesh [35] Source: Bohrer et al. [36] Tree-dependent variables:canopy leaf area density, seed release height, seed quantity, seed terminal velocity, fraction velocity, the height above the ground surface, dry deposition velocity, point source concentration, etc. Environmental variables:above-canopy wind statistics, air temperature, wind velocity, air density, acceleration due to gravity, geostrophic wind, parameters of eddy diffusivity for near and far distribution, etc. | Bohrer et al. [36]; Di-Giovanni and Beckett [37]; Loos et al. [34]; Maurer et al. [38]; Nathan et al., [39]; Thompson et al. [40]; Treep et al. [41]; Treep et al. [42]; Williams et al. [43]; Wright et al. [44] |
| Slope Degree (°) | Correction Factor | |
|---|---|---|
| Up Slope | Down Slope | |
| 5 | 0.92 | 1.10 |
| 10 | 0.85 | 1.21 |
| 15 | 0.79 | 1.37 |
| 20 | 0.73 | 1.57 |
| 25 | 0.68 | 1.87 |
| 30 | 0.63 | 2.37 |
| 35 | 0.59 | 3.34 |
| 40 | 0.54 | 6.22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.; Lee, S.; Lee, S.; Yi, K.; Kim, H.-S.; Chung, S.; Chung, J.; Kim, H.S.; Yoon, T.K. Seed Dispersal Models for Natural Regeneration: A Review and Prospects. Forests 2022, 13, 659. https://doi.org/10.3390/f13050659
Kim M, Lee S, Lee S, Yi K, Kim H-S, Chung S, Chung J, Kim HS, Yoon TK. Seed Dispersal Models for Natural Regeneration: A Review and Prospects. Forests. 2022; 13(5):659. https://doi.org/10.3390/f13050659
Chicago/Turabian StyleKim, Moonil, Seonghun Lee, Songhee Lee, Koong Yi, Hyung-Sub Kim, Sanghoon Chung, Junmo Chung, Hyun Seop Kim, and Tae Kyung Yoon. 2022. "Seed Dispersal Models for Natural Regeneration: A Review and Prospects" Forests 13, no. 5: 659. https://doi.org/10.3390/f13050659
APA StyleKim, M., Lee, S., Lee, S., Yi, K., Kim, H. -S., Chung, S., Chung, J., Kim, H. S., & Yoon, T. K. (2022). Seed Dispersal Models for Natural Regeneration: A Review and Prospects. Forests, 13(5), 659. https://doi.org/10.3390/f13050659