
Fire as a Major Factor in Dynamics of Tree-Growth and Stable δ13C and δ18O Variations in Larch in the Permafrost Zone

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
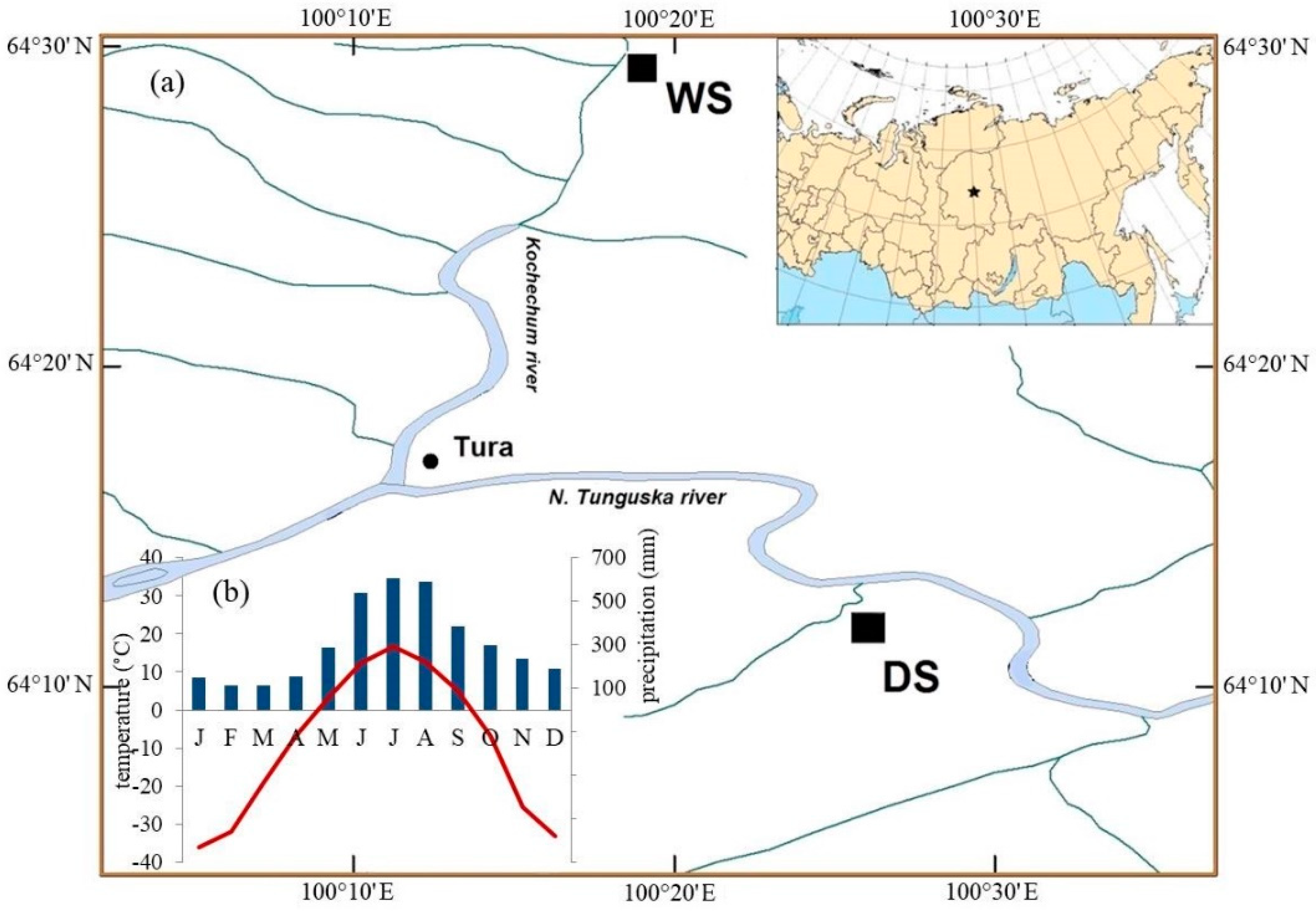
2.1. Site Description
2.2. Basic Principles of Isotope Fractionation in C3 Plants
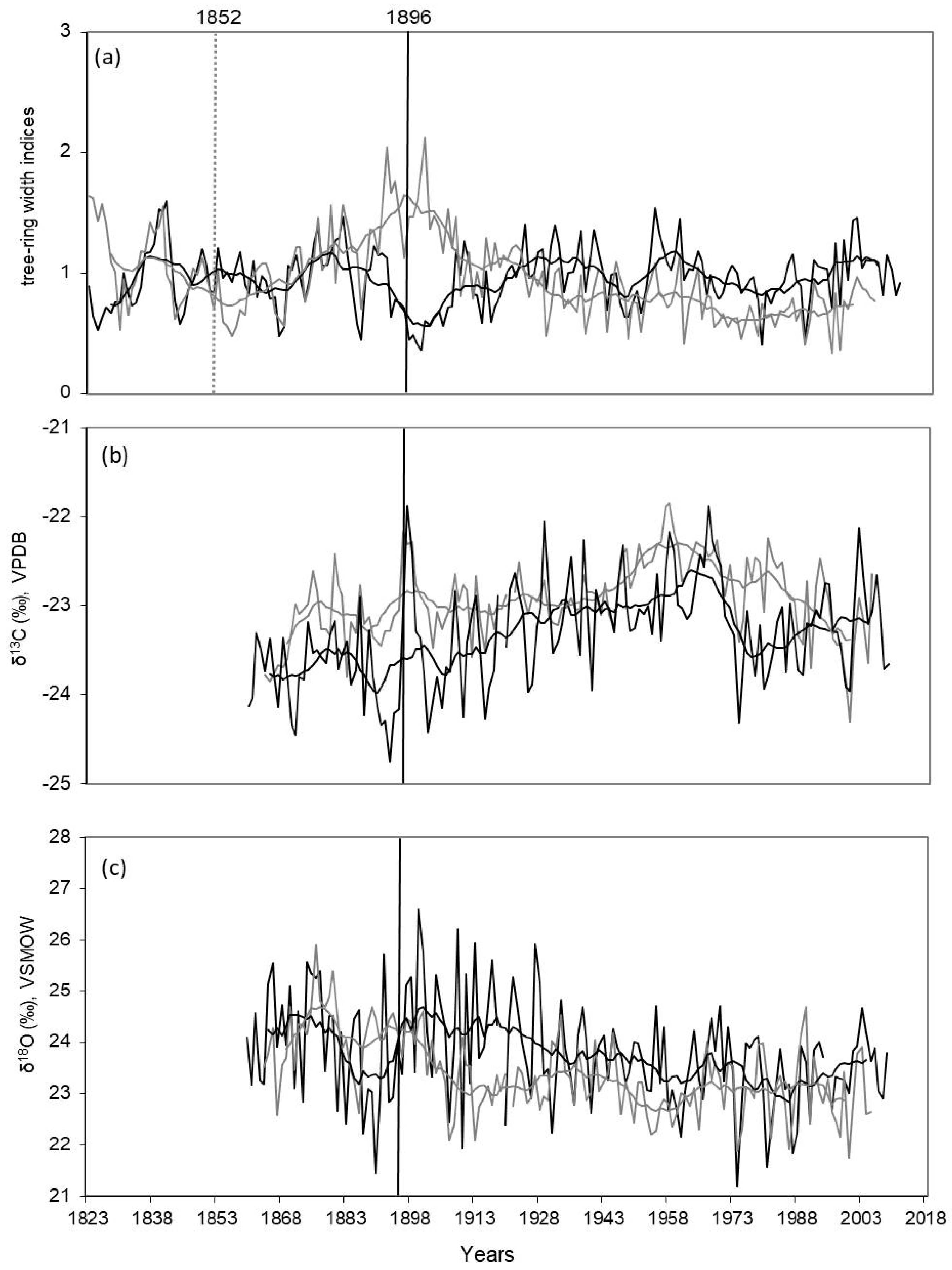
3. Results
3.1. Tree-Ring width, δ13C and δ18O Dynamics
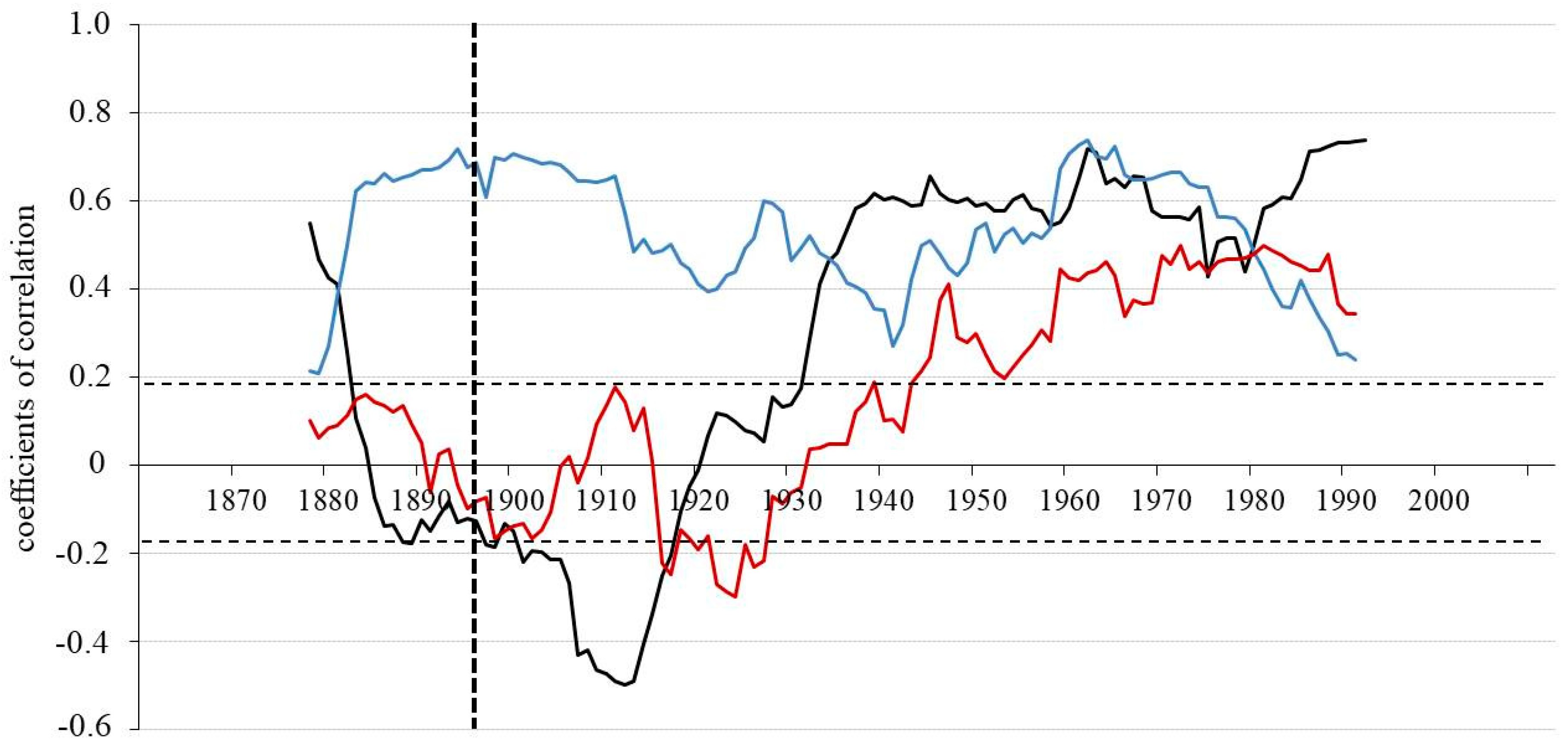
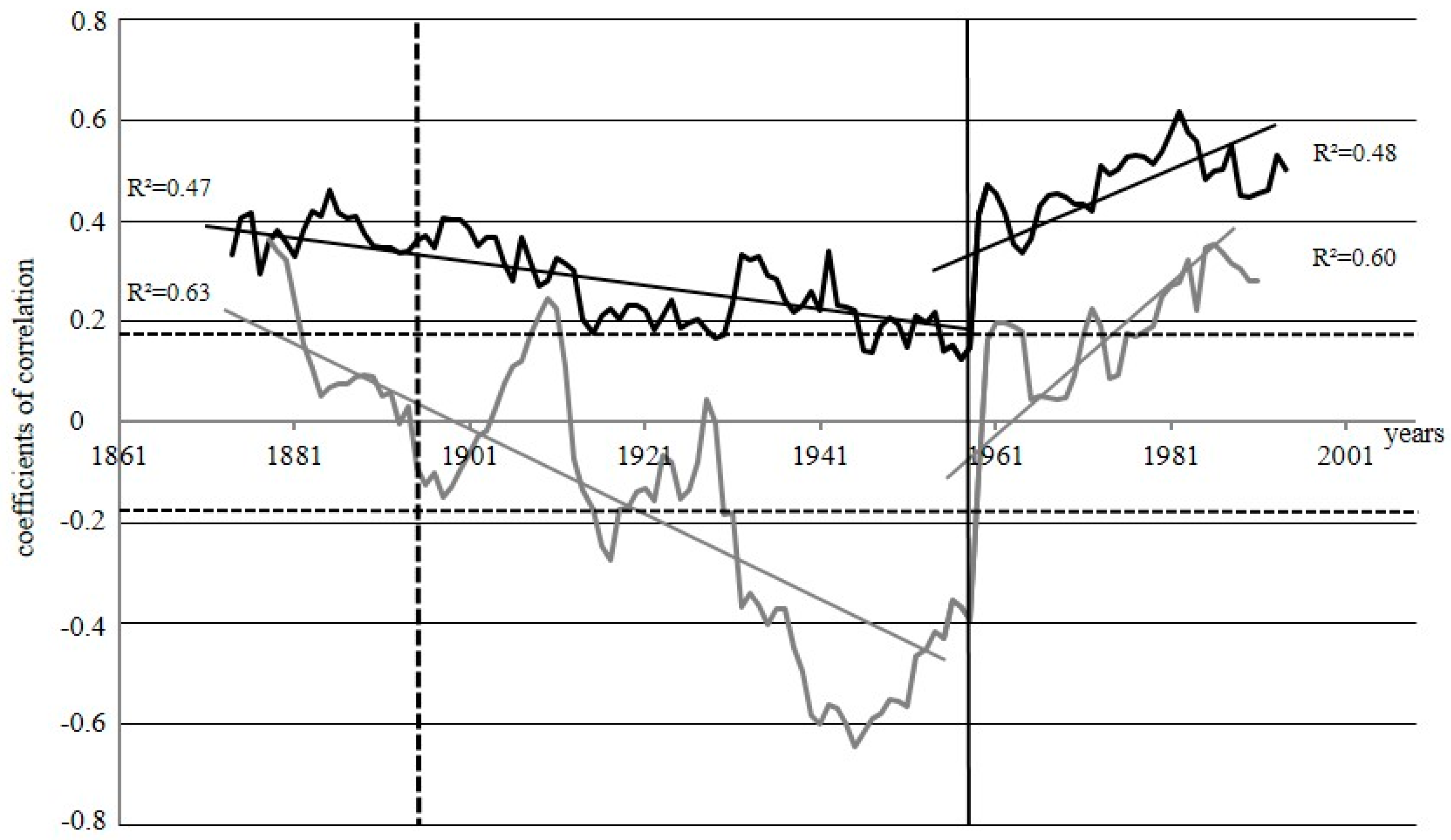
3.2. Relationships between Tree-Ring Parameters
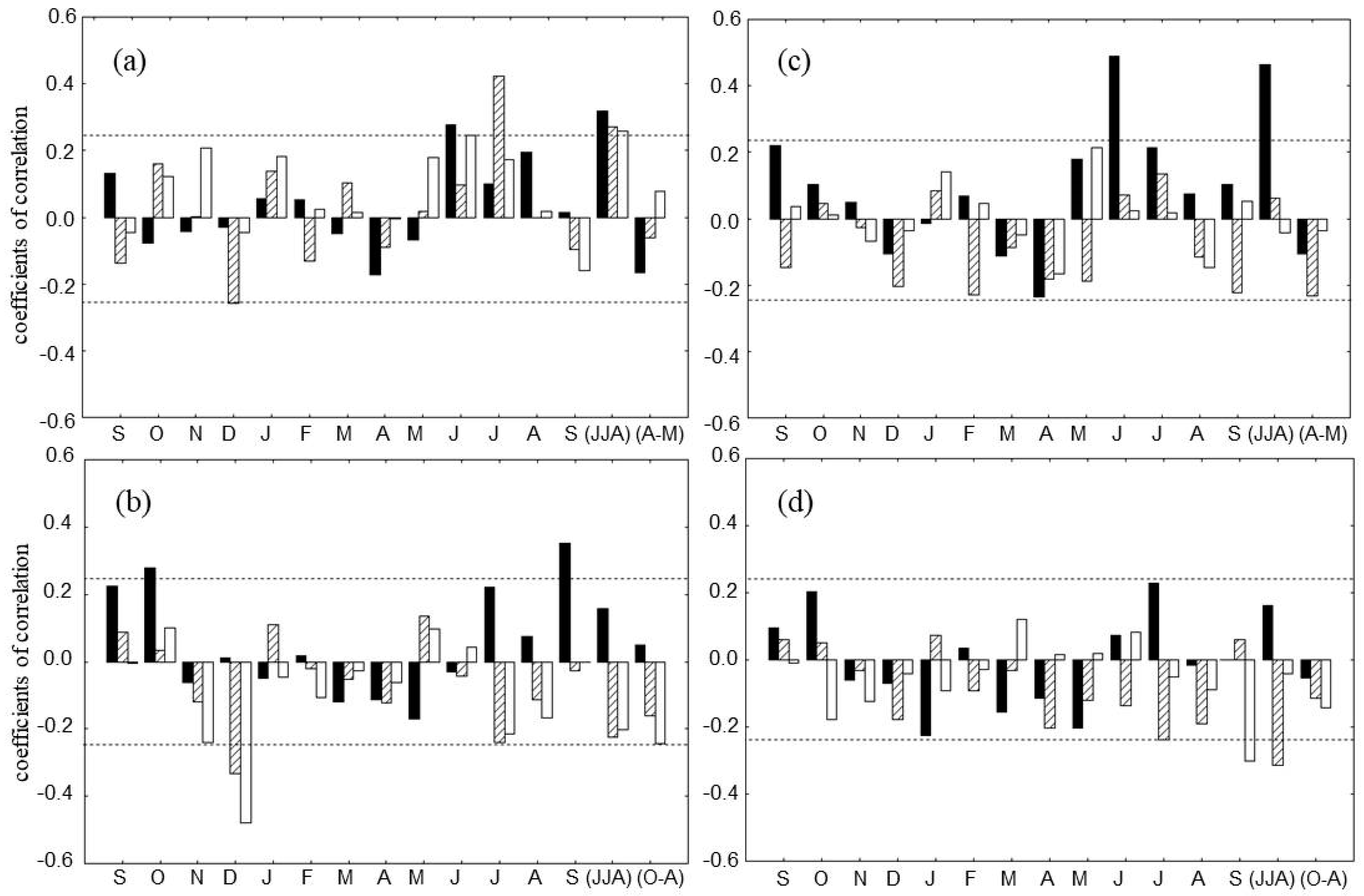
3.3. Climatic Response
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Osawa, A.; Zyryanova, O.A.; Matsuura, Y.; Kajimoto, T.; Wein, R.W. (Eds.) Permafrost Ecosystems: Siberian Larch Forests. In Ecological Studies; Springer: Berlin/Heidelberg, Germany, 2010; Volume 209. [Google Scholar] [CrossRef]
- Biskaborn, B.K.; Smith, S.L.; Noetzli, J.; Matthes, H.; Vieira, G.; Streletskiy, D.A.; Schoeneich, P.; Romanovsky, V.E.; Lewkowicz, A.G.; Abramov, A.; et al. Permafrost is warming at a global scale. Nat. Commun. 2019, 10, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, H.; Meissner, K.J.; Mix, A.C.; Abram, N.J.; Austermann, J.; Brovkin, V.; Capron, E.; Colombaroli, D.; Daniau, A.-L.; Dyez, K.; et al. Palaeoclimate constraints on the impact of 2 °C anthropogenic warming and beyond. Nat. Geosci. 2018, 11, 474–485. [Google Scholar] [CrossRef]
- Schaefer, K.; Lantuit, H.; Romanovsky, V.E.; Schuur, E.A.G.; Witt, R. The impact of the permafrost carbon feedback on global climate. Environ. Res. Lett. 2014, 9, 085003. [Google Scholar] [CrossRef] [Green Version]
- Clow, G.D.; Urban, F.E. Largest Permafrost Warming in Northern Alaska during the 1990s Determined from GTN-P Borehole Temperature Measurements; EOS, Transactions of the AGU: Washington, DC, USA, 2002; Volume 47, p. F258. [Google Scholar]
- Turetsky, M.R.; Kelman, R.; Vitt, D.H. Boreal peatland C fluxes under varying permafrost regimes. Soil Biol. Biochem. 2002, 34, 907–912. [Google Scholar] [CrossRef]
- Romanovsky, V.E.; Sazonova, T.S.; Balobaev, V.T.; Shender, N.I.; Sergueev, D.O. Past and recent changes in permafrost and air temperatures in Eastern Siberia. Glob. Planet. Change 2007, 56, 399–413. [Google Scholar] [CrossRef]
- Romanovsky, V.E.; Kholodov, A.L.; Marchenko, S.S.; Oberman, N.G.; Drozdov, D.S.; Malkova, G.V.; Moskalenko, N.G.; Vasiliev, A.A.; Sergeev, D.O.; Zheleznyak, M.N. NICOP-Thermal State and Fate of Permafrost in Russia First Results of IPY. In Proceedings of the Ninth International Conference on Permafrost, Fairbanks, Alaska, 29 June–3 July 2008. [Google Scholar]
- Romanovsky, V.E.; Smith, S.L.; Christiansen, H.H. Permafrost thermal state in the Polar Northern Hemisphere during the international polar year 2007–2009: A synthesis. Permafr. Periglac. Process. 2010, 21, 106–116. [Google Scholar] [CrossRef] [Green Version]
- Euskirchen, E.S.; McGuire, A.D.; Kicklighter, D.W.; Zhuang, Q.; Clein, J.S.; Dargaville, R.J.; Dye, D.G.; Kimball, J.S.; McDonald, K.C.; Melollo, J.M.; et al. Importance of recent shifts in soil thermal dynamics on growing season length, productivity, and carbon sequestration in terrestrial high-latitude ecosystems. Glob. Change Biol. 2006, 12, 731–750. [Google Scholar] [CrossRef] [Green Version]
- Knorre, A.A.; Kirdyanov, A.V.; Prokushkin, A.S.; Krusic, P.J.; Büntgen, U. Tree ring- based reconstruction of the long-term influence of wildfires on permafrost active layer dynamics in Central Siberia. Sci. Total Environ. 2019, 652, 314–319. [Google Scholar] [CrossRef] [Green Version]
- Köster, E.; Köster, K.; Berninger, F.; Aaltonen, H.; Zhou, X.; Pumpanen, J. Carbon dioxide, methane and nitrous oxide fluxes from a fire chronosequence in subarctic boreal forests of Canada. Sci. Total Environ. 2017, 601–602, 895–905. [Google Scholar] [CrossRef] [Green Version]
- Kirdyanov, A.V.; Prokushkin, A.S.; Tabakova, M.A. Tree-ring growth of Gmelin larch under contrasting local conditions in the north of Central Siberia. Dendrochronologia 2013, 31, 114–119. [Google Scholar] [CrossRef]
- Kirdyanov, A.; Treydte, K.S.; Nikolaev, A.; Helle, G.; Schleser, G.H. Climate signals in tree-ring width, wood density and δ13C from larches in Eastern Siberia (Russia). Chem. Geol. Incl. Isot. Geosci. 2008, 252, 31–41. [Google Scholar]
- Safronov, M.A.; Volokitina, A.V. Wildfire ecology in continuous permafrost zone. In Book Permafrost Ecosystems: Siberian Larch Forests; Osawa, A., Zyryanova, O.A., Matsuura, Y., Kajimoto, T., Wein, R.W., Eds.; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; London, UK; New York, NY, USA, 2010; Volume 209, pp. 59–83. [Google Scholar]
- Churakova (Sidorova), O.V.; Shashkin, A.V.; Siegwolf, R.T.W.; Spahni, R.; Launois, T.; Saurer, M.; Bryukhanova, M.V.; Benkova, A.V.; Kupzova, A.V.; Vaganov, E.A.; et al. Application of eco-physiological models to the climatic interpretation of δ13C and δ18O measured in Siberian larch tree-rings. Dendrochronologia 2016, 39, 51–59. [Google Scholar] [CrossRef]
- Churakova-Sidorova, O.V.; Lienert, S.; Timofeeva, G.; Siegwolf, R.; Roden, J.; Joos, F.; Saurer, M. Measured and modelled source water δ18O based on tree-ring cellulose of larch and pine trees from the permafrost zone. For.-Biogeosci. For. 2020, 13, 224–229. [Google Scholar] [CrossRef]
- Saurer, M.; Kirdyanov, A.V.; Prokushkin, A.S.; Rinne, K.T.; Siegwolf, R.T.W. The relationship between soil and needle water oxygen isotope composition of Larix gmelinii in Siberia. New Phytol. 2016, 209, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Prokushkin, A.S.; Hobara, S.; Prokushkin, S.G. Behavior of Dissolved Organic Carbon in Larch Ecosystems. In Book Permafrost Ecosystems: Siberian Larch Forests; Osawa, A., Zyryanova, O.A., Matsuura, Y., Kajimoto, T., Wein, R.W., Eds.; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; London, UK; New York, NY, USA, 2010; Volume 209, pp. 205–228. [Google Scholar]
- Kharuk, V.I.; Ranson, K.J.; Dvinskaya, M.L. Wildfires dynamic in the larch dominance zone. Geophys. Res. Lett. 2008, 35, L01402. [Google Scholar] [CrossRef]
- Furyaev, V.V.; Vaganov, E.A.; Tchebakova, N.M.; Valendik, E.N. Effects of Fire and climate on Succession and structural changes in The Siberian Boreal Forest. Eurasian J. For. Res. 2001, 2, 1–15. [Google Scholar]
- Ivanova, G.A. Extremely fire risk seasons in forest of Evenkia. Sib. J. Ecol. 1996, 1, 29–34. (In Russian) [Google Scholar]
- Arbatskaya, M.K. Long-Term Variability of Climate, Tree Growth and Fire Frequencies in Taiga of Middle Siberia (In Russian). Ph.D. Thesis, Institute of Forest SB RAS, Krasnoyarsk, Russia, 1998. [Google Scholar]
- Swetnam, T.W. Fire history and climate change in giant sequoia groves. Science 1993, 262, 885–889. [Google Scholar] [CrossRef]
- Grissino-Mayer, H.D.; Romme, W.H.; Floyd, M.L.; Hanna, D.D. Climatic and human influences on fire regime of the southern San Juan Mountains, Colorado, USA. Ecology 2004, 85, 1708–1724. [Google Scholar] [CrossRef] [Green Version]
- Kharuk, V.I.; Kenneth, J.R.; Dvinskaya, M.L.; Im, S.T. Wildfires in northern Siberian larch dominated communities. Environ. Res. Lett. 2011, 6, 045208. [Google Scholar] [CrossRef]
- McCarroll, D.; Loader, N.J. Stable isotopes in tree rings. Quat. Sci. Rev. 2004, 23, 771–801. [Google Scholar] [CrossRef]
- Gagen, M.H.; McCarrol, D.; Loader, N.J.; Robertson, I.; Jalkanen, R.; Anchukaitis, K.J. Exorcising the ‘segment length curse’ summer temperature reconstruction since AD 1640 using non de-trend stable carbon isotope ratios from line trees in northern Finland. Holocene 2007, 17, 433–444. [Google Scholar] [CrossRef]
- Treydte, K.; Frank, D.; Esper, J.; Andreu, L.; Bednarz, Z.; Berninger, F.; Boettger, T.; D’Alessandro, C.M.; Etien, N.; Filot, M.; et al. Signal strength and climate calibration of a European tree ring isotope network. Geophys. Res. Lett. 2007, 34, L24302. [Google Scholar] [CrossRef] [Green Version]
- Saurer, M.; Cherubini, P.; Reynolds-Henne, C.E.; Treydte, K.S.; Anderson, W.T.; Siegwolf, R.T.W. An investigation of the common signal in tree ring stable isotope chronologies at temperate sites. J. Geophys. Res. 2008, 113, 11. [Google Scholar] [CrossRef]
- Leavitt, S.W.; Danzer, S.R. Method for batch processing of small wood samples to holocellulose for stable carbon isotope analysis. Anal. Chem. 1993, 65, 87–89. [Google Scholar] [CrossRef]
- McCarroll, D.; Pawellek, F. Stable carbon isotope ratios of Pinus sylvestris from northern Finland and the potential for extracting a climate signal from long Fennoscandian chronologies. Holocene 2001, 11, 517–526. [Google Scholar] [CrossRef]
- Aguilera, M.; Espinar, C.; Ferrio, J.P.; Pérez, G.; Voltas, J. A map of autumn precipitation for the third millennium BP in the Eastern Iberian Peninsula from charcoal carbon isotopes. J. Geochem. Explor. 2009, 102, 157–165. [Google Scholar] [CrossRef]
- Aguilera, M.; Ferrio, J.P.; Araus, J.L.; Tarrús, J.; Voltas, J. Climate at the onset of western Mediterranean agriculture expansion: Evidence from stable isotopes of sub-fossil oak tree rings in Spain. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 299, 541–551. [Google Scholar] [CrossRef]
- Sidorova, O.V.; Siegwolf, R.T.W.; Saurer, M.; Shashkin, A.V.; Knorre, A.A.; Prokushkin, A.S.; Vaganov, E.A.; Kirdyanov, A.V. Do centennial tree-ring and stable isotope trends of Larix gmelinii (Rupr.) Rupr. indicate increasing water shortage in the Siberian north? Oecologia 2009, 161, 825–835. [Google Scholar] [CrossRef] [Green Version]
- Scheidegger, Y.; Saurer, M.; Bahn, M.; Siegwolf, R. Linking stable isotopes with stomatal conductance and phytosynthetic capacity: A conceptual model. Oecologia 2000, 125, 350–357. [Google Scholar] [CrossRef]
- Saurer, M.; Siegwolf, R.T.W. Human impacts on tree ring growth reconstructed from stable isotopes. In Book Stable Isotopes as Indicators of Ecological Change; Dawson, T.E., Siegwolf, R.T.W., Eds.; Elsevier Inc.: London, UK, 2007; pp. 49–62. [Google Scholar]
- Kagawa, A.; Naito, D.; Sugimoto, A.; Maximov, T.C. Effects of spatial variability in soil moisture on widths and δ13C values of eastern Siberian tree rings. J. Geophys. Res. 2003, 108, 4500. [Google Scholar] [CrossRef]
- Kagawa, A.; Sugimoto, A.; Maximov, T.C. Seasonal course of translocation, storage and remobilization of 13C pulse-labeled photoassimilate in naturally growing Larix gmelinii saplings. New Phytol. 2006, 171, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Sidorova, O.V.; Siegwolf, R.T.W.; Saurer, M.; Naurzbaev, M.; Vaganov, E.A. Isotopic composition (δ13C, δ18O) in wood and cellulose of Siberian larch trees for early Medieval and recent periods. J. Geophys. Res. 2008, 113, G02019. [Google Scholar] [CrossRef] [Green Version]
- Churakova (Sidorova), O.V.; Fonti, M.V.; Saurer, M.; Guillet, S.; Corona, C.; Fonti, P.; Myglan, V.S.; Kirdyanov, A.V.; Naumova, O.V.; Ovchinnikov, D.V.; et al. Siberian tree-ring and stable isotope proxies as indicators of temperature and moisture changes after major stratospheric volcanic eruptions. Clim. Past. 2019, 15, 685–700. [Google Scholar] [CrossRef] [Green Version]
- Kirdyanov, A.V.; Saurer, M.; Siegwolf, R.; Knorre, A.A.; Prokushkin, A.S.; Churakova (Sidorova), O.V.; Fonti, M.V.; Büntgen, U. Long-term ecological consequences of forest fires in the continuous permafrost zone of Siberia. Environ. Res. Lett. 2020, 15, 034061. [Google Scholar] [CrossRef]
- Kirdyanov, A.V.; Hughes, M.; Vaganov, E.A.; Schweingruber, F.; Silkin, P. The importance of early summer temperature and date of snow melt for tree growth in the Siberian Subarctic. Trees 2003, 17, 61–69. [Google Scholar] [CrossRef]
- Sidorova, O.V.; Siegwolf, R.T.W.; Saurer, M.; Naurzbaev, M.M.; Shashkin, A.V.; Vaganov, E.A. Spatial patterns of climatic changes in the Eurasian north reflected in Siberian larch tree-ring parameters and stable isotopes. Glob. Change Biol. 2010, 16, 1003–1018. [Google Scholar] [CrossRef] [Green Version]
- Bryukhanova, M.V.; Fonti, P.; Kirdyanov, A.V.; Siegwolf, R.T.W.; Saurer, M.; Pochebyt, N.P.; Churakova (Sidorova), O.V.; Prokushkin, A.S. The response of δ13C, δ18O and cell anatomy of Larix gmelinii tree rings to differing soil active layer depths. Dendrochronologia 2015, 34, 51–59. [Google Scholar] [CrossRef]
- Abaimov, A.P.; Bondarev, A.I.; Zyryanova, O.A.; Shitova, S.A. Polar Forests of Krasnoyarsk Region; Nauka: Novosibirsk, Russia, 1997; 208p. [Google Scholar]
- USDA. Keys to Soil Taxonomy, 8th ed.; USDA: Washington, DC, USA, 1998; 326p.
- Vodop’yanova, N.S. Vegetation of Putorana. In Book Flora of Putorana; Malyshev, L.I., Ed.; Nauka: Novosibirsk, Russia, 1976; pp. 11–31. (In Russian) [Google Scholar]
- Speer, J.H. Fundamentals of Tree-Ring Research; University of Arizona Press: Tucson, AZ, USA, 2010; 509p. [Google Scholar]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Saurer, M.; Siegenthaler, U.; Schweingruber, F. The climate-carbon isotope relationship in tree rings and the significance of site conditions. Tellus Ser. B. 1995, 47, 320–330. [Google Scholar] [CrossRef] [Green Version]
- Borella, S.; Leuenberger, M.; Saurer, M. Reducing uncertainties in δ13C analysis of tree rings: Pooling, milling, and cellulose extraction. J. Geophys. Res. 1998, 103, 19519–19526. [Google Scholar] [CrossRef]
- Loader, N.J.; Robertson, I.; Barker, A.C.; Switsur, V.R.; Waterhouse, J.S. Improved technique for the batch processing of small whole wood samples to alpha-cellulose. Chem. Geol. 1997, 136, 313–317. [Google Scholar] [CrossRef]
- Saurer, M.; Robertson, L.; Siegwolf, R.; Leuenberger, M. Oxygen isotope analysis of cellulose: An interlaboratory comparison. Anal. Chem. 1998, 70, 2074–2080. [Google Scholar] [CrossRef]
- Werner, R.A.; Bruch, B.A.; Brand, W.A. ConFlo III—An Interface for High Precision δ13C and δ15N Analysis with an Extended Dynamic Range. Rapid Commun. Mass Spectrom. 1999, 13, 1237–1241. [Google Scholar] [CrossRef]
- Saurer, M.; Borella, S.; Schweingruber, F.; Siegwolf, R. Stable carbon isotopes in tree rings of beech: Climatic versus site-related influences. Tree 1997, 11, 291–297. [Google Scholar] [CrossRef] [Green Version]
- Francey, R.J.; Allison, C.E.; Etheridge, D.M. A 1000-year high precision record of δ13C in atmospheric CO2. Tellus Ser. B. 1999, 51, 170–193. [Google Scholar] [CrossRef] [Green Version]
- McCarroll, D.; Gagen, M.H.; Loader, N.J.; Robertson, I.; Anchukaitis, K.J.; Los, S.; Young, G.; Jalkanen, R.; Kirchhefer, A.; Waterhouse, J.S. Correction of tree ring stable carbon isotope chronologies for changes in the carbon dioxide content of the atmosphere. Geochim. Cosmochim. Acta 2009, 73, 1539–1547. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Lloyd, J. Carbon and oxygen isotope effects in the exchange of carbon dioxide between plants and the atmosphere. In Book J.R. Stable Isotope and Plant Carbon/Water Relation; Ehleringer, A.E.H., Farquhar, G.D., Eds.; Academic Press: San Diego, CA, USA, 1993; pp. 47–70. [Google Scholar]
- Barbour, M.; Cernusak, L.A.; Farquhar, G.D. Factors affecting the Oxygen isotope ratio of plant organic matter. In Book Stable Isotopes and Biosphere—Atmosphere Interactions: Processes and Biological Controls; Flanagan, L.B., Ehleringer, J.R., Pataki, D.E., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Dongmann, G.; Nürnberg, H.W.; Förstel, H.; Wagener, K. On the enrichment of H218O in the leaves of transpiring plants. Radiat. Environ. Biophys. 1974, 11, 41–52. [Google Scholar] [CrossRef]
- Lehmann, M.M.; Goldsmith, G.R.; Mirande-Ney, C.; Weigt, R.B.; Schoenbeck, L.; Kahmen, A.; Gessler, A.; Siegwolf, R.T.W.; Saurer, M. The O-18-signal transfer from water vapour to leaf water and assimilates varies among plant species and growth forms. Plant Cell Environ. 2020, 43, 510–523. [Google Scholar] [CrossRef]
- Prokushkin, A.S.; Hagedorn, F.; Pokrovsky, O.S.; Viers, J.; Kirdyanov, A.V.; Masyagina, O.V.; Prokushkina, M.P.; McDowell, W.H. Permafrost regime affects the nutritional status and productivity of larches in central Siberia. Forests 2018, 9, 314. [Google Scholar] [CrossRef] [Green Version]
- Zyryanova, O.A.; Bugaenko, T.N.; Abaimov, A.P.; Bugaenko, N.N. Pyrogenic Transformation of Species Diversity in Larch Forests of the Permafrost Zone. In Permafrost Ecosystems: Siberian Larch Forests; Springer: Dordrecht, The Netherlands, 2010; Volume 209, pp. 135–146. [Google Scholar]
- Knorre, A.A.; Kirdyanov, A.V.; Prokushkin, A.S. Estimation of Rate of Elevating Permafrost Table under Larch Stands in the Cryolithozone of Central Evenkia by Dendrochronological Methods (In Russian). Lesovedenie 2009, 2, 77–80. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | ||
|---|---|---|
| Parameters | WS | DS |
| Location | 64° 32′ N 100°14′ E 204 m a.s.l | 64°12′ N 100°27′ E 218 m a.s.l |
| Type of vegetation | larch with dwarf-shrubs, lichens (10%) and mosses (90%) cover | larch with shrubs, dwarf-shrubs, mosses (40%) and lichens (20%) cover |
| Depth of ASL, cm | 25.5 ± 21.9 | 57.7 ± 18.6 |
| Last detected fire events, year | 1852 | 1896 |
| Mean height of trees (H), m | 10.5 ± 6.2 | 16.0 ± 1.3 |
| Mean diameter on the breast height (DBH), cm | 8.7 ± 3.8 | 25.9 ± 3.2 |
| Forest density, N/ha | 1325 | 400 |
| TRW chronology, period | 1742–2005 | 1823–2011 |
| Mean TRW values, mm | 0.34 ± 0.19 | 0.64 ± 0.45 |
| Stable isotope chronology, period | 1864–2005 | 1860–2011 |
| Mean δ13CCell values, ‰ VPDB | −22.93 ± 0.44 | −23.32 ± 0.57 |
| Mean δ18OCell values, ‰ VSMOW | 23.40 ± 0.77 | 23.78 ± 1.0 |
| WS | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1867–1896 | 1897–1926 | 1927–1956 | 1957–1986 | ||||||||||
| TRW | δ13C | δ18O | TRW | δ13C | δ18O | TRW | δ13C | δ18O | TRW | δ13C | δ18O | ||
| DS | TRW | 0.39 | 0.35 | 0.62 | −0.46 | 0.03 | −0.36 | 0.65 | −0.27 | −0.08 | 0.49 | 0.16 | −0.41 |
| δ13C | −0.38 | 0.18 | 0.05 | 0.09 | 0.66 | 0.12 | −0.40 | 0.27 | −0.20 | 0.17 | 0.67 | 0.19 | |
| δ18O | −0.26 | 0.11 | 0.08 | 0.35 | 0.22 | 0.17 | 0.15 | 0.01 | 0.11 | 0.16 | 0.28 | 0.46 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knorre, A.A.; Siegwolf, R.T.W.; Kirdyanov, A.V.; Saurer, M.; Churakova, O.V.; Prokushkin, A.S. Fire as a Major Factor in Dynamics of Tree-Growth and Stable δ13C and δ18O Variations in Larch in the Permafrost Zone. Forests 2022, 13, 725. https://doi.org/10.3390/f13050725
Knorre AA, Siegwolf RTW, Kirdyanov AV, Saurer M, Churakova OV, Prokushkin AS. Fire as a Major Factor in Dynamics of Tree-Growth and Stable δ13C and δ18O Variations in Larch in the Permafrost Zone. Forests. 2022; 13(5):725. https://doi.org/10.3390/f13050725
Chicago/Turabian StyleKnorre, Anastasia A., Rolf T. W. Siegwolf, Alexander V. Kirdyanov, Matthias Saurer, Olga V. Churakova (Sidorova), and Anatoly S. Prokushkin. 2022. "Fire as a Major Factor in Dynamics of Tree-Growth and Stable δ13C and δ18O Variations in Larch in the Permafrost Zone" Forests 13, no. 5: 725. https://doi.org/10.3390/f13050725
APA StyleKnorre, A. A., Siegwolf, R. T. W., Kirdyanov, A. V., Saurer, M., Churakova, O. V., & Prokushkin, A. S. (2022). Fire as a Major Factor in Dynamics of Tree-Growth and Stable δ13C and δ18O Variations in Larch in the Permafrost Zone. Forests, 13(5), 725. https://doi.org/10.3390/f13050725