
The Historical Complexity of Tree Height Growth Dynamic Associated with Climate Change in Western North America

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
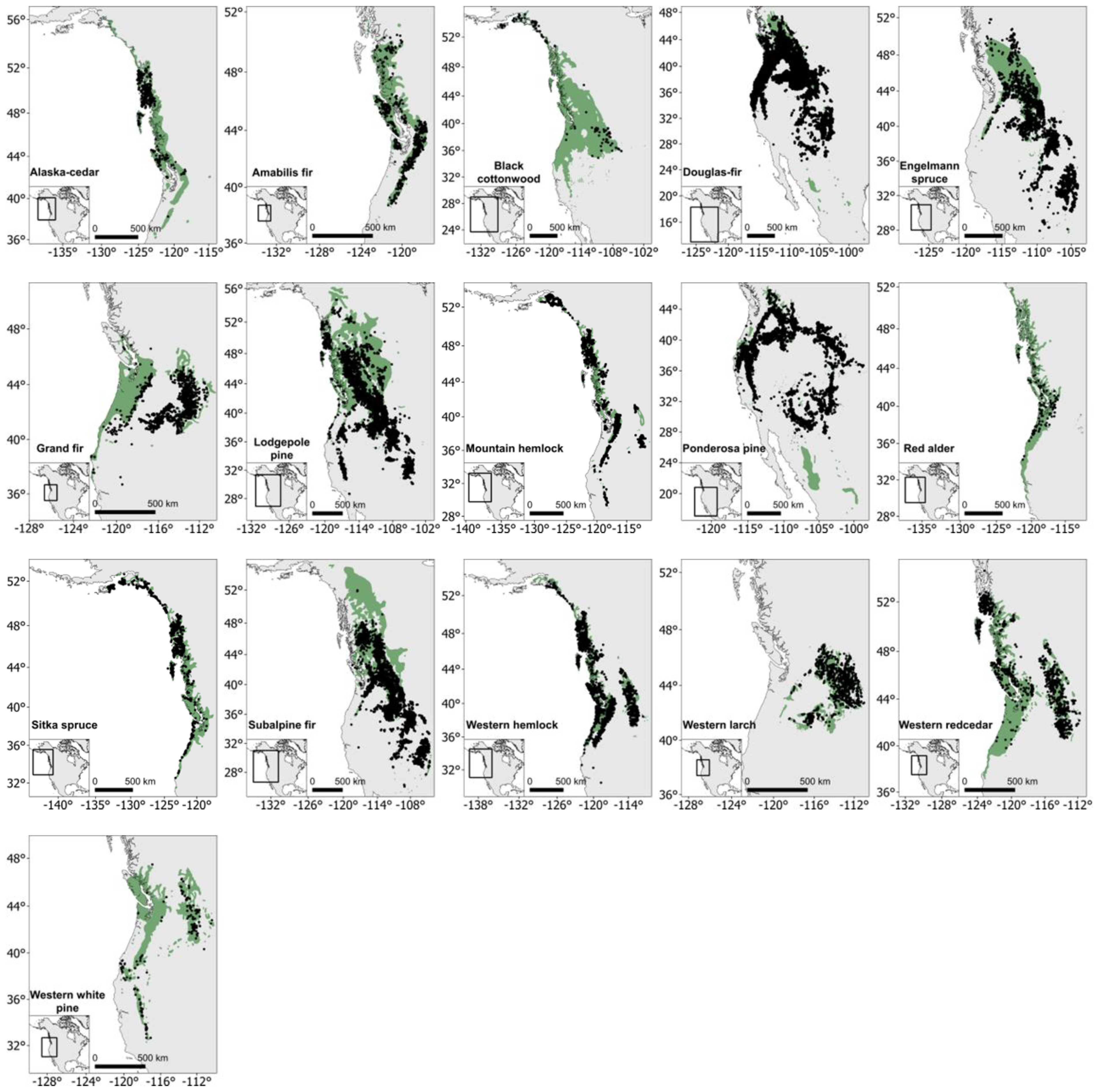
2.1. Study Area
2.2. Sampling Design
2.3. Tree Height and Site Index
2.4. Study Species
2.5. Soil Properties
2.6. Climate Variables
2.7. Statistical Analyses
2.8. Anomaly of Site Index
3. Results
3.1. Tree Site Index and Climate
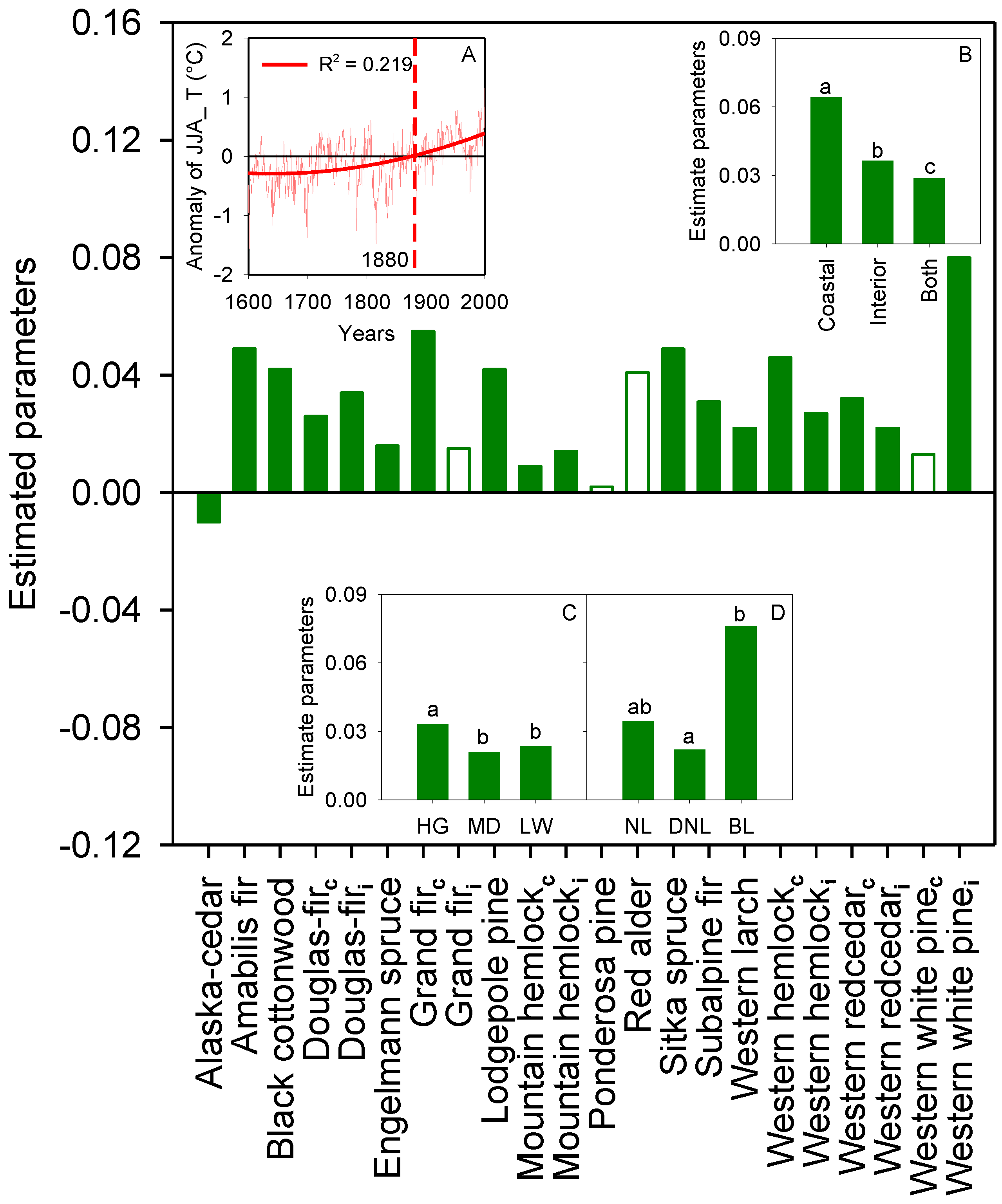
3.2. Global Growth Patterns
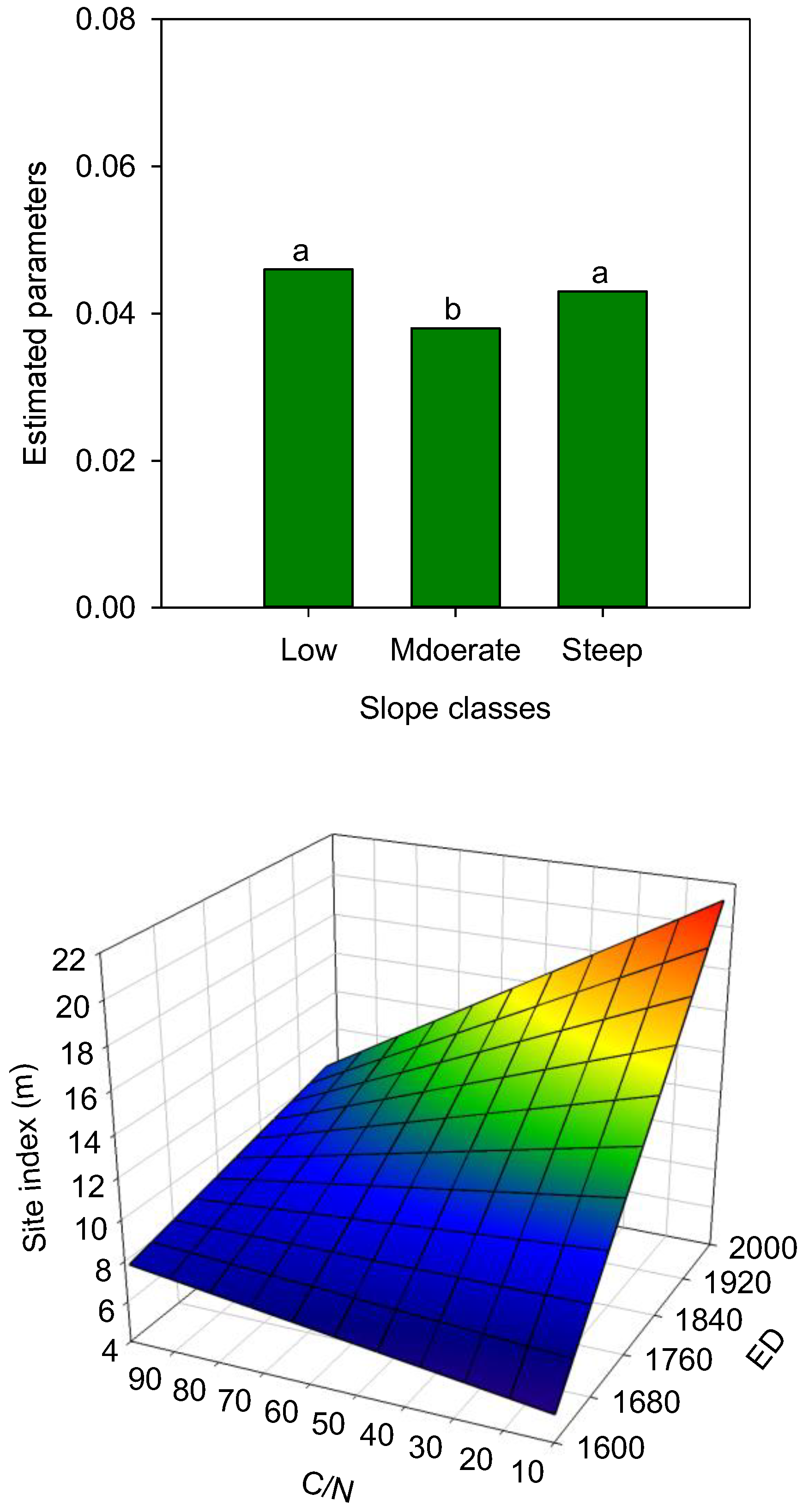
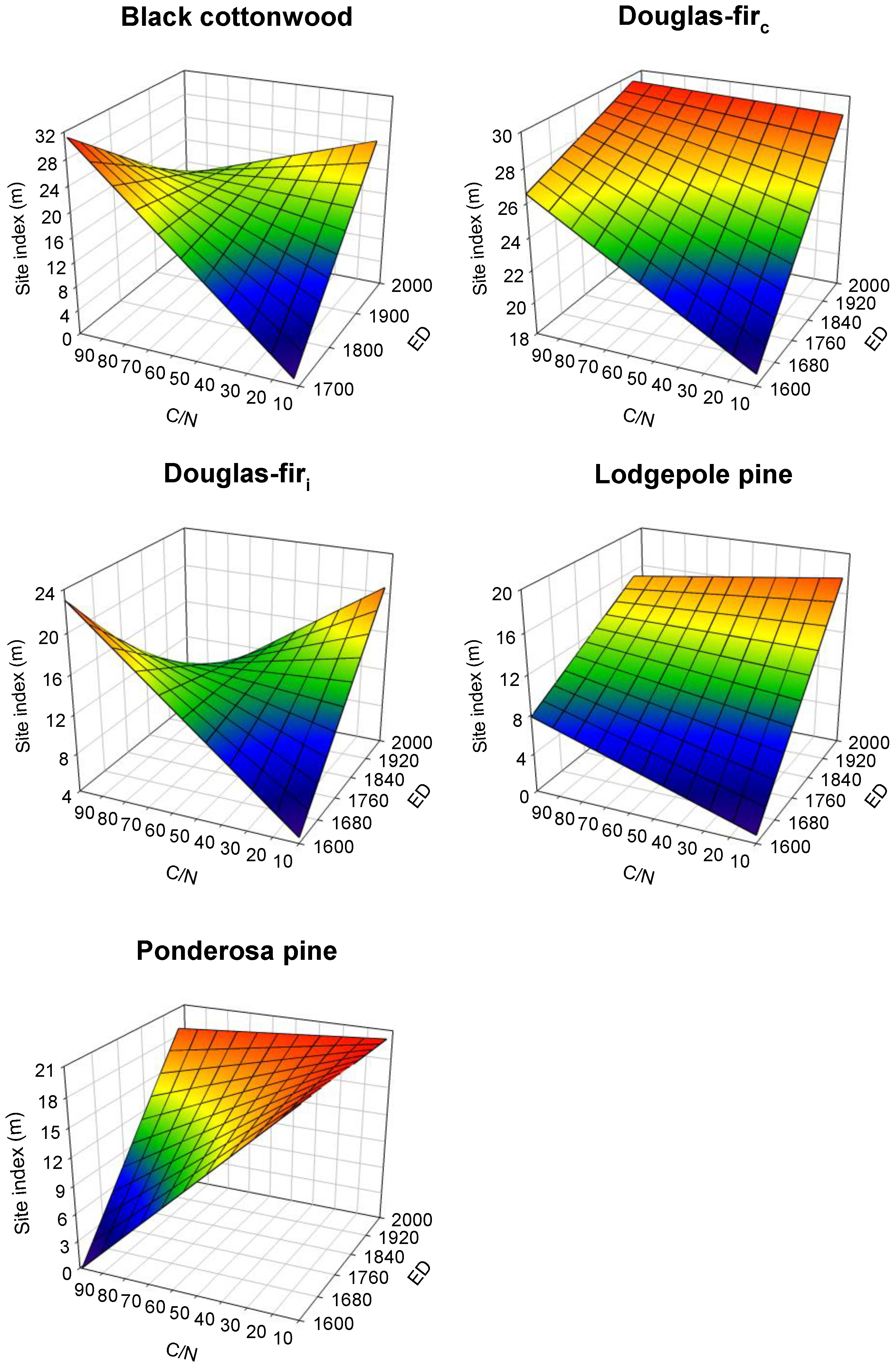
3.3. Species-Scale Growth Patterns
3.4. Anomaly of Species Site Index and Climate Change
4. Discussion
4.1. Relationship between Height Growth and Temporal Variables
4.2. Global Height Growth Patterns
4.3. Species-Specific Growth Patterns
4.4. Spatio-Temporal Variation in Species’ Tree Growth
4.5. Growth Patterns with Distinct Geographic Ranges
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brecka, A.F.J.; Shahi, C.; Chen, H.Y.H. Climate change impacts on boreal forest timber supply. For. Policy Econ. 2018, 92, 11–21. [Google Scholar] [CrossRef]
- Holz, A.; Hart, S.J.; Williamson, G.J.; Veblen, T.T.; Aravena, J.C. Radial growth response to climate change along the latitudinal range of the world’s southernmost conifer in southern South America. J. Biogeogr. 2018, 45, 1140–1152. [Google Scholar] [CrossRef]
- Tei, S.; Sugimoto, A. Time lag and negative responses of forest greenness and tree growth to warming over circumboreal forests. Glob. Change Biol. 2018, 24, 4225–4237. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guo, M.; Wang, X.; Gu, F.; Liu, S. Divergent tree growth response to recent climate warming of Abies faxoniana at alpine treelines in east edge of Tibetan Plateau. Ecol. Res. 2017, 33, 303–311. [Google Scholar] [CrossRef]
- Wieser, G.; Matyssek, R.; Luzian, R.; Zwerger, P.; Pindur, P.; Oberhuber, W.; Gruber, A. Effects of atmospheric and climate change at the timberline of the Central European Alps. Ann. Sci. 2009, 66, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwab, N.; Kaczka, R.; Janecka, K.; Böhner, J.; Chaudhary, R.; Scholten, T.; Schickhoff, U. Climate change-Induced shift of tree growth sensitivity at a central himalayan treeline ecotone. Forests 2018, 9, 267. [Google Scholar] [CrossRef] [Green Version]
- Devi, N.; Hagedorn, F.; Moiseev, P.; Bugmann, H.; Shiyatov, S.; Mazepa, V.; Rigling, A. Expanding forests and changing growth forms of Siberian larch at the Polar Urals treeline during the 20th century. Glob. Change Biol. 2008, 14, 1581–1591. [Google Scholar] [CrossRef]
- Cienciala, E.; Altman, J.; Dolezal, J.; Kopacek, J.; Stepanek, P.; Stahl, G.; Tumajer, J. Increased spruce tree growth in Central Europe since 1960s. Sci. Total Environ. 2018, 619–620, 1637–1647. [Google Scholar] [CrossRef]
- Tchebakova, N.M.; Parfenova, E.I.; Korets, M.A.; Conard, S.G. Potential change in forest types and stand heights in central Siberia in a warming climate. Environ. Res. Lett. 2016, 11, 035016. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, A.H.; Bunn, A.G. Responses of the circumpolar boreal forest to 20th century climate variability. Environ. Res. Lett. 2007, 2, 045013. [Google Scholar] [CrossRef]
- Dietrich, R.; Bell, F.W.; Silva, L.C.R.; Cecile, A.; Horwath, W.R.; Anand, M. Climatic sensitivity, water-use efficiency, and growth decline in boreal jack pine (Pinus banksiana) forests in Northern Ontario. J. Geophys. Res. Biogeosci. 2016, 121, 2761–2774. [Google Scholar] [CrossRef]
- Sangines de Carcer, P.; Vitasse, Y.; Penuelas, J.; Jassey, V.E.J.; Buttler, A.; Signarbieux, C. Vapor-pressure deficit and extreme climatic variables limit tree growth. Glob. Change Biol. 2018, 24, 1108–1122. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, M.J.; Engelbrecht, B.M.J.; Joswig, J.; Pereyra, G.; Schuldt, B.; Jansen, S.; Kattge, J.; Landhäusser, S.M.; Levick, S.R.; Preisler, Y.; et al. A synthesis of tree functional traits related to drought-induced mortality in forests across climatic zones. J. Appl. Ecol. 2017, 54, 1669–1686. [Google Scholar] [CrossRef]
- Pallardy, S.G.; Kozlowski, T.T. Physiology of Woody Plants. Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Woodward, F.I. Ecophysiological Controls of Conifer Distributions. In Physiological Ecology; Academic Press: San Diego, CA, USA, 1995. [Google Scholar]
- Saxe, H.; Cannell, M.G.R.; Johnsen, B.; Ryan, M.G.; Vourlitis, G. Tree and forest functioning in response to global warming. New Phytol. 2002, 149, 369–400. [Google Scholar] [CrossRef]
- Cahoon, S.M.P.; Sullivan, P.F.; Brownlee, A.H.; Pattison, R.R.; Andersen, H.E.; Legner, K.; Hollingsworth, T.N. Contrasting drivers and trends of coniferous and deciduous tree growth in interior Alaska. Ecology 2018, 99, 1284–1295. [Google Scholar] [CrossRef]
- Lacourse, T. Environmental change controls postglacial forest dynamics through interspecific differences in life-history traits. Ecology 2009, 90, 2149–2160. [Google Scholar] [CrossRef]
- Littell, J.S.; Peterson, D.L.; Tjoelker, M. Douglas-fir growth in mountain ecosystems: Water limits tree growth from stand to region. Ecol. Monogr. 2008, 78, 349–368. [Google Scholar] [CrossRef]
- Gao, L.; Zhang, Y.; Wang, X.; Zhang, C.; Zhao, Y.; Liu, L. Sensitivity of Three Dominant Tree Species from the Upper Boundary of Their Forest Type to Climate Change at Changbai Mountain, Northeastern China. Tree Ring Res. 2018, 74, 39–49. [Google Scholar] [CrossRef]
- Kremenetski, C.V.; Sulerzhitsky, L.D.; Hantemirov, R. Holocene history of the northern range limits of some trees and shrubs in Russia. Arct. Alp Res. 1998, 30, 317–333. [Google Scholar] [CrossRef]
- Martin-Benito, D.; Pederson, N.; Köse, N.; Doğan, M.; Bugmann, H.; Mosulishvili, M.; Bigler, C. Pervasive effects of drought on tree growth across a wide climatic gradient in the temperate forests of the Caucasus. Glob. Ecol Biogeogr. 2018, 27, 1314–1325. [Google Scholar] [CrossRef]
- Babst, F.; Poulter, B.; Trouet, V.; Tan, K.; Neuwirth, B.; Wilson, R.; Carrer, M.; Grabner, M.; Tegel, W.; Levanic, T.; et al. Site-and species-specific responses of forest growth to climate across the European continent. Glob. Ecol. Biogeogr. 2013, 22, 706–717. [Google Scholar] [CrossRef]
- Lopatin, E. Long-term trends in height growth of Picea obovata and Pinus sylvestris during the past 100 years in Komi Republic (north-western Russia). Scand. J. For. Res. 2007, 22, 310–323. [Google Scholar] [CrossRef]
- Barber, V.A.; Juday, G.P.; Finney, B.P. Reduced growth of Alaskan white spruce in the twentieth century from temperature-induced drought stress. Nature 2000, 405, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Hynes, A.; Hamann, A. Moisture deficits limit growth of white spruce in the west-central boreal forest of North America. For. Ecol. Manag. 2020, 461, 117944. [Google Scholar] [CrossRef]
- Hogg, E.H.; Brandt, J.P.; Michaelian, M. Impacts of a regional drought on the productivity, dieback, and biomass of western Canadian aspen forests. Can. J. For. Res. 2008, 38, 1373–1384. [Google Scholar] [CrossRef] [Green Version]
- Hogg, E.H.; Brandt, J.P.; Kochtubajda, B. Growth and dieback of aspen forests in northwestern Alberta, Canada, in relation to climate and insects. Can. J. For. Res. 2002, 32, 823–832. [Google Scholar] [CrossRef]
- Hararuk, O.; Campbell, E.M.; Antos, J.A.; Parish, R. Tree rings provide no evidence of a CO2 fertilization effect in old-growth subalpine forests of western Canada. Glob. Change Biol. 2018, 25, 1222–1234. [Google Scholar] [CrossRef]
- Messaoud, Y.; Chen, H.Y. The influence of recent climate change on tree height growth differs with species and spatial environment. PLoS ONE 2011, 6, e14691. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, P.F.; Pattison, R.R.; Brownlee, A.H.; Cahoon, S.M.P.; Hollingsworth, T.N. Limited evidence of declining growth among moisture-limited black and white spruce in interior Alaska. Sci. Rep. 2017, 7, 15344. [Google Scholar] [CrossRef]
- Peterson, D.W.; Peterson, D.L. Effects of climate on radial growth of sub-alpine conifers in the north Cascade Mountains. Can. J. For. Res. 1994, 24, 1921–1932. [Google Scholar] [CrossRef]
- Miyamoto, Y.; Griesbauer, H.P.; Scott Green, D. Growth responses of three coexisting conifer species to climate across wide geographic and climate ranges in Yukon and British Columbia. For. Ecol. Manag. 2010, 259, 514–523. [Google Scholar] [CrossRef]
- Lévesque, M.; Walthert, L.; Weber, P.; Jones, R. Soil nutrients influence growth response of temperate tree species to drought. J. Ecol. 2016, 104, 377–387. [Google Scholar] [CrossRef]
- Gleason, K.E.; Bradford, J.B.; D’Amato, A.W.; Fraver, S.; Palik, B.J.; Battaglia, M.A. Forest density intensifies the importance of snowpack to growth in water-limited pine forests. Ecol. Appl. 2020, 31, e02211. [Google Scholar] [CrossRef] [PubMed]
- Hofgaard, A.; Løkken, J.O.; Dalen, L.; Hytteborn, H. Comparing warming and grazing effects on birch growth in an alpine environment—A 10-year experiment. Plant Ecol. Divers. 2010, 3, 19–27. [Google Scholar] [CrossRef]
- Sanders-DeMott, R.; McNellis, R.; Jabouri, M.; Templer, P.H.; Wurzburger, N. Snow depth, soil temperature and plant-herbivore interactions mediate plant response to climate change. J. Ecol. 2018, 106, 1508–1519. [Google Scholar] [CrossRef]
- Wang, X.; Pederson, N.; Chen, Z.; Lawton, K.; Zhu, C.; Han, S. Recent rising temperatures drive younger and southern Korean pine growth decline. Sci. Total Environ. 2019, 649, 1105–1116. [Google Scholar] [CrossRef]
- Nigh, G.D. Impact of climate, moisture regime, and nutrient regime on the productivity of Douglas-Fir in coastal British Columbia, Canada. Clim. Change 2006, 76, 321–337. [Google Scholar] [CrossRef]
- Chen, H.Y.H.; Klinka, K.; Kabzems, R.D. Site index, site quality, and foliar nutrients of trembling aspen: Relationships and predictions. Can. J. For. Res. 1998, 28, 1743–1755. [Google Scholar] [CrossRef]
- Monserud, R.A. Height growth and site index curves for inland Douglas-fir based on stem analysis data and forest habitat type. For. Sci. 1984, 4, 943–965. [Google Scholar]
- Ritchie, M.; Zhang, J.; Hamilton, T. Effects of Stand Density on Top Height Estimation for Ponderosa Pine. West. J. Appl. For. 2012, 27, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Burns, R.M.; Honkala, B.H. Silvics of North America; United States Department of Agriculture, USDA Forest Service: Washington, DC, USA, 1990.
- Klinka, K.; Worrall, J.; Skoda, L.; Varga, P. The Distribution and Synopsis of Ecological and Silvical Characteristics of Tree Species of British Columbia’s Forests; The Forest Sciences Department, The University of British Columbia: Vancouver, BC, Canada, 2000. [Google Scholar]
- Viereck, L.A.; Little, E.L. Alaska Trees and Shrubs; University of Alaska Press: Fairbanks, AK, USA, 2007. [Google Scholar]
- Soil Landscapes of Canada Working Group. Soil Landscapes of Canada; version 3.2; Agriculture and Agri-Food Canada: Edmonton, AB, Canada, 2010.
- MacMillan, R.A.; Pettapiece, W.W.; Brierley, J.A. An expert system for allocating soils to landforms through the application of soil survey tacit knowledge. Can. J. Soil Sci. 2005, 85, 103–112. [Google Scholar] [CrossRef]
- Guillet, S.; Corona, C.; Stoffel, M.; Khodri, M.; Lavigne, F.; Ortega, P.; Eckert, N.; Sielenou, P.D.; Daux, V.; Churakova, O.V.; et al. Climate response to the Samalas volcanic eruption in 1257 revealed by proxy records. Nat. Geo. 2017, 10, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Shi, F.; Yang, B.; Mairesse, A.; Von Gunten, L.; Li, J.; Yang, F.; Xiao, X. Northern Hemisphere temperature reconstruction during the last millennium using multiple annual proxies. Clim. Res. 2013, 56, 231–244. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Hamann, A.; Spittlehouse, D.; Carroll, C. Locally Downscaled and Spatially Customizable Climate Data for Historical and Future Periods for North America. PLoS ONE 2016, 11, e0156720. [Google Scholar] [CrossRef]
- Dai, A.; Trenberth, K.E.; Qian, T.T. A global dataset of Palmer Drought Severity Index for 1870–2002: Relationship with soil moisture and effects of surface warming. J. Hydrometeorol. 2004, 5, 1117–1130. [Google Scholar] [CrossRef]
- Steiger, N.J.; Smerdon, J.E.; Cook, E.R.; Cook, B.I. A reconstruction of global hydroclimate and dynamical variables over the Common Era. Sci. Data 2018, 5, 180086. [Google Scholar] [CrossRef]
- Legates, D.R. Climate models and their simulation of precipitation. Energy Environ. 2014, 25, 1163–1175. [Google Scholar] [CrossRef]
- Tapiador, F.J.; Roca, R.; Del Genio, A.; Dewitte, B.; Petersen, W.; Zhang, F. Is precipitation a good metric for model performance? Bull. Am. Meteorol. Soc. 2019, 100, 223–233. [Google Scholar] [CrossRef]
- Messaoud, Y. Les Facteurs Écologiques et les Mécanismes Expliquant la Localisation de L’écotone Sapinière/Pessière Dans L’ouest du Québec; Université du Québec en Abitibi-Témiscamingue: Rouyn-Noranda, QC, Canada, 2007. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Mote, P.W. Trends in snow water equivalent in the Pacific Northwest and their climatic causes. Geophys. Res. Lett. 2003, 30. [Google Scholar] [CrossRef] [Green Version]
- Abatzoglou, J.T.; Rupp, D.E.; Mote, P.W. Seasonal Climate Variability and Change in the Pacific Northwest of the United States. J. Clim. 2014, 27, 2125–2142. [Google Scholar] [CrossRef]
- Mildrexler, D.; Yang, Z.; Cohen, W.B.; Bell, D.M. A forest vulnerability index based on drought and high temperatures. Remote Sens. Environ. 2016, 173, 314–325. [Google Scholar] [CrossRef] [Green Version]
- Lyu, S.; Wang, X.; Zhang, Y.; Li, Z. Different responses of Korean pine (Pinus koraiensis) and Mongolia oak (Quercus mongolica) growth to recent climate warming in northeast China. Dendrochronologia 2017, 45, 113–122. [Google Scholar] [CrossRef]
- Nigh, G.D.; Ying, C.C.; Qian, H. Climate and productivity of major conifer species in the interior of British Columbia, Canada. For. Sci. 2004, 50, 659–671. [Google Scholar]
- Sharma, M.; Parton, J. Climatic Effects on Site Productivity of Red Pine Plantations. For. Sci. 2018, 64, 544–554. [Google Scholar] [CrossRef]
- Adams, J.M. Vegetation-Climate Interaction: How Vegetation Makes the Global Environment; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2007. [Google Scholar]
- Cortini, F.; Comeau, P.G.; Boateng, J.O.; Bedford, L.; McClarnon, J.; Powelson, A. Effects of climate on growth of lodgepole pine and white spruce following site preparation and its implications in a changing climate. Can. J. For. Res. 2011, 41, 180–194. [Google Scholar] [CrossRef]
- Zhou, Y.; Lei, Z.; Zhou, F.; Han, Y.; Yu, D.; Zhang, Y. Impact of climate factors on height growth of Pinus sylvestris var. mongolica. PLoS ONE 2019, 14, e0213509. [Google Scholar] [CrossRef] [Green Version]
- Erickson, C.C.; Waring, K.M. Old Pinus ponderosa growth responses to restoration treatments, climate and drought in a southwestern US landscape. App. Veg. Sci. 2014, 17, 97–108. [Google Scholar] [CrossRef]
- Finlayson, B. Köppen-Geiger Climate Classification. In Encyclopedia of Geography; SAGE: Thousand Oaks, CA, USA, 2010; pp. 1666–1669. [Google Scholar]
- Beedlow, P.A.; Lee, E.H.; Tingey, D.T.; Waschmann, R.S.; Burdick, C.A. The importance of seasonal temperature and moisture patterns on growth of Douglas-fir in western Oregon, USA. Agric. For. Meteorol. 2013, 169, 174–185. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.P.; Still, C.J.; Fischer, D.T.; Leavitt, S.W. The influence of summertime fog and overcast clouds on the growth of a coastal Californian pine: A tree-ring study. Oecologia 2008, 156, 601–611. [Google Scholar] [CrossRef]
- Cortini, F.; Comeau, P.G.; Wang, T.; Hibbs, D.E.; Bluhm, A. Climate effects on red alder growth in the Pacific Northwest of North America. For. Ecol. Manag. 2012, 277, 98–106. [Google Scholar] [CrossRef] [Green Version]
- Deal, R.L. Red Alder: A State of Knowledge; Department of Agriculture, Forest Service: Portland, OR, USA, 2006.
- Appleton, S.N.; George, S.S. High-elevation Mountain hemlock growth as a surrogate for cool-season precipitation in Crater Lake National Park, USA. Dendrochronologia 2018, 52, 20–28. [Google Scholar] [CrossRef]
- Gustafson, E.J.; Miranda, B.R.; De Bruijn, A.M.G.; Sturtevant, B.R.; Kubiske, M.E. Do rising temperatures always increase forest productivity? Interacting effects of temperature, precipitation, cloudiness and soil texture on tree species growth and competition. Environ. Model Soft. 2017, 97, 171–183. [Google Scholar] [CrossRef]
- Kajimoto, T.; Seki, T.; Ikeda, S.; Daimaru, H.; Okamoto, T.; Onodera, H. Effects of Snowfall Fluctuation on Tree Growth and Establishment of SubalpineAbies mariesiinear Upper Forest-limit of Mt. Yumori, Northern Japan. Arct. Antarct. Alp. Res. 2018, 34, 191–200. [Google Scholar] [CrossRef]
- Beier, C.M.; Sink, S.E.; Hennon, P.E.; D’Amore, D.V.; Juday, G.P. Twentieth-century warming and the dendroclimatology of declining yellow-cedar forests in southeastern Alaska. Can. J. For. Res. 2008, 38, 1319–1334. [Google Scholar] [CrossRef] [Green Version]
- Hennon, P.E.; D’Amore, D.V.; Schaberg, P.G.; Wittwer, D.T.; Shanley, C.S. Shifting Climate, Altered Niche, and a Dynamic Conservation Strategy for Yellow-Cedar in the North Pacific Coastal Rainforest. Bioscience 2012, 62, 147–158. [Google Scholar] [CrossRef] [Green Version]
- Schaberg, P.G.; D’Amore, D.V.; Hennon, P.E.; Halman, J.M.; Hawley, G.J. Do limited cold tolerance and shallow depth of roots contribute to yellow-cedar decline? For. Ecol. Manag. 2011, 262, 2142–2150. [Google Scholar] [CrossRef]
- Yener, I.; Altun, L. Predicting site index for oriental spruce (Picea orientalis L. (LINK)) using ecological factors in the eastern Black Sea, Turkey. Fresenius Environmental Bulletin 2018, 27, 3107–3116. [Google Scholar]
- Sedmáková, D.; Sedmák, R.; Bosela, M.; Ježík, M.; Blaženec, M.; Hlásny, T.; Marušák, R. Growth-climate responses indicate shifts in the competitive ability of European beech and Norway spruce under recent climate warming in East-Central Europe. Dendrochronologia 2019, 54, 37–48. [Google Scholar] [CrossRef]
- Bailey, R. Ecoregions: The Ecosystem Geography of the Oceans and Continents; Springer: New York, NY, USA, 2014. [Google Scholar]
- Orians, G.; Schoen, J. North Pacific Temperate Rainforests: Ecology and Conservation; University of Washington Press: Seattle, WA, USA, 2017. [Google Scholar]
- Coops, N.C.; Waring, R.H.; Schroeder, T.A. Combining a generic process-based productivity model and a statistical classification method to predict the presence and absence of tree species in the Pacific Northwest, USA. Ecol. Model 2009, 220, 1787–1796. [Google Scholar] [CrossRef]
- Lambers, H.; Pons, T.L.; Chapin, F.S., III. Plant Physiological Ecology, 2nd ed.; Springer: New York, NY, USA, 2008. [Google Scholar]
- Karnosky, D.F.; Zak, D.R.; Pregitzer, K.S.; Awmack, C.S.; Bockheim, J.G.; Dickson, R.E.; Hendrey, G.R.; Host, G.E.; King, J.S.; Kopper, B.J.; et al. Tropospheric O3 moderates responses of temperate hardwood forests to elevated CO2: A synthesis of molecular to ecosystem results from the Aspen FACE project. Funct. Ecol. 2003, 17, 289–304. [Google Scholar] [CrossRef] [Green Version]
- Weemstra, M.; Eilmann, B.; Sass-Klaassen, U.G.W.; Sterck, F.J. Summer droughts limit tree growth across 10 temperate species on a productive forest site. For. Ecol. Manag. 2013, 306, 142–149. [Google Scholar] [CrossRef]
- Eckhart, T.; Potzelsberger, E.; Koeck, R.; Thom, D.; Lair, G.J.; van Loo, M.; Hasenauer, H. Forest stand productivity derived from site conditions: An assessment of old Douglas-fir stands (Pseudotsuga menziesii (Mirb.) Franco var. menziesii) in Central Europe. Ann. For. Sci. 2019, 76, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonelli, G.; Denneler, B.; Bergeron, Y. Climate sensitivity of trembling aspen radial growth along a productivity gradient in northeastern British Columbia, Canada. Can. J. For. Res. 2008, 38, 1211–1222. [Google Scholar] [CrossRef]
- Machmuller, M.B.; Kramer, M.G.; Cyle, T.K.; Hill, N.; Hancock, D.; Thompson, A. Emerging land use practices rapidly increase soil organic matter. Nat. Commun. 2015, 6, 6995. [Google Scholar] [CrossRef] [PubMed]
- Bhadha, J.; Capasso, J.; Khatiwada, R.; Swanson, S.; Laborde, C. Raising Soil Organic Matter Content to Improve Water Holding Capacity; SL447; Institute of Food and Agricultural Sciences: Gainesville, FL, USA, 2017. [Google Scholar]
- Gao, L.; Gou, X.; Deng, Y.; Wang, Z.; Gu, F.; Wang, F. Increased growth of Qinghai spruce in northwestern China during the recent warming hiatus. Agric. For. Meteorol. 2018, 260–261, 9–16. [Google Scholar] [CrossRef]
- Venegas-González, A.; Juñent, F.R.; Gutiérrez, A.G.; Filho, M.T. Recent radial growth decline in response to increased drought conditions in the northernmost Nothofagus populations from South America. For. Ecol. Manag. 2018, 409, 94–104. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Fries, A.; Lindgren, D.; Cheng, C.Y.; Ruotsalainen, S.; Lindgren, K.; Elfving, B.; Karlmats, U. The effect of temperature on site index in western Canada and Scandinavia estimated from IUFRO Pinus contorta provenance experiments. Can. J. For. Res. 2000, 30, 921–929. [Google Scholar] [CrossRef]
- Rodríguez-Ramírez, E.C.; Luna-Vega, I.; Rozas, V. Tree-Ring Research of Mexican Beech (Fagus GrandifoliaSubsp.Mexicana) A Relict Tree Endemic to Eastern Mexico. Tree Ring Res. 2018, 74, 94–107. [Google Scholar] [CrossRef]
- Ganey, J.L.; Vojta, S.C. Tree mortality in drought-stressed mixed-conifer and ponderosa pine forests, Arizona, USA. For. Ecol. Manag. 2011, 261, 162–168. [Google Scholar] [CrossRef]
- Restaino, C.; Young, D.J.N.; Estes, B.; Gross, S.; Wuenschel, A.; Meyer, M.; Safford, H. Forest structure and climate mediate drought-induced tree mortality in forests of the Sierra Nevada, USA. Ecol. Appl. 2019, 29, e01902. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, L.D.; Anderegg, W.R.; Abatzoglou, J.; Hausladen, A.M.; Berry, J.A. Drought characteristics’ role in widespread aspen forest mortality across Colorado, USA. Glob. Change Biol. 2013, 19, 1526–1537. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Forests. Biogeoclimatic Zones of British Columbia; 1:12000000 Coloured Map; Ministry of Forests, Research Branch: Victoria, BC, Canada, 1988.
- Gavin, D.G.; Hu, F.S. Spatial variation of climatic and non-climatic controls on species distribution: The range limit of Tsuga heterophylla. J. Biogeogr. 2006, 33, 1384–1396. [Google Scholar] [CrossRef]
- Gavin, D.G. The coastal-disjunct mesic flora in the inland Pacific Northwest of USA and Canada: Refugia, dispersal and disequilibrium. Divers. Distrib. 2009, 15, 972–982. [Google Scholar] [CrossRef]
- Asselin, H.; Payette, S.; Fortin, M.-J.; Vallée, S. The northern limit of Pinus banksiana Lamb. in Canada: Explaining the difference between the eastern and western distributions. J. Biogeogr. 2003, 30, 1709–1718. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, J.F.; Chapin, F.S. Non-equilibrium succession dynamics indicate continued northern migration of lodgepole pine. Glob. Change Biol. 2003, 9, 1401–1409. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
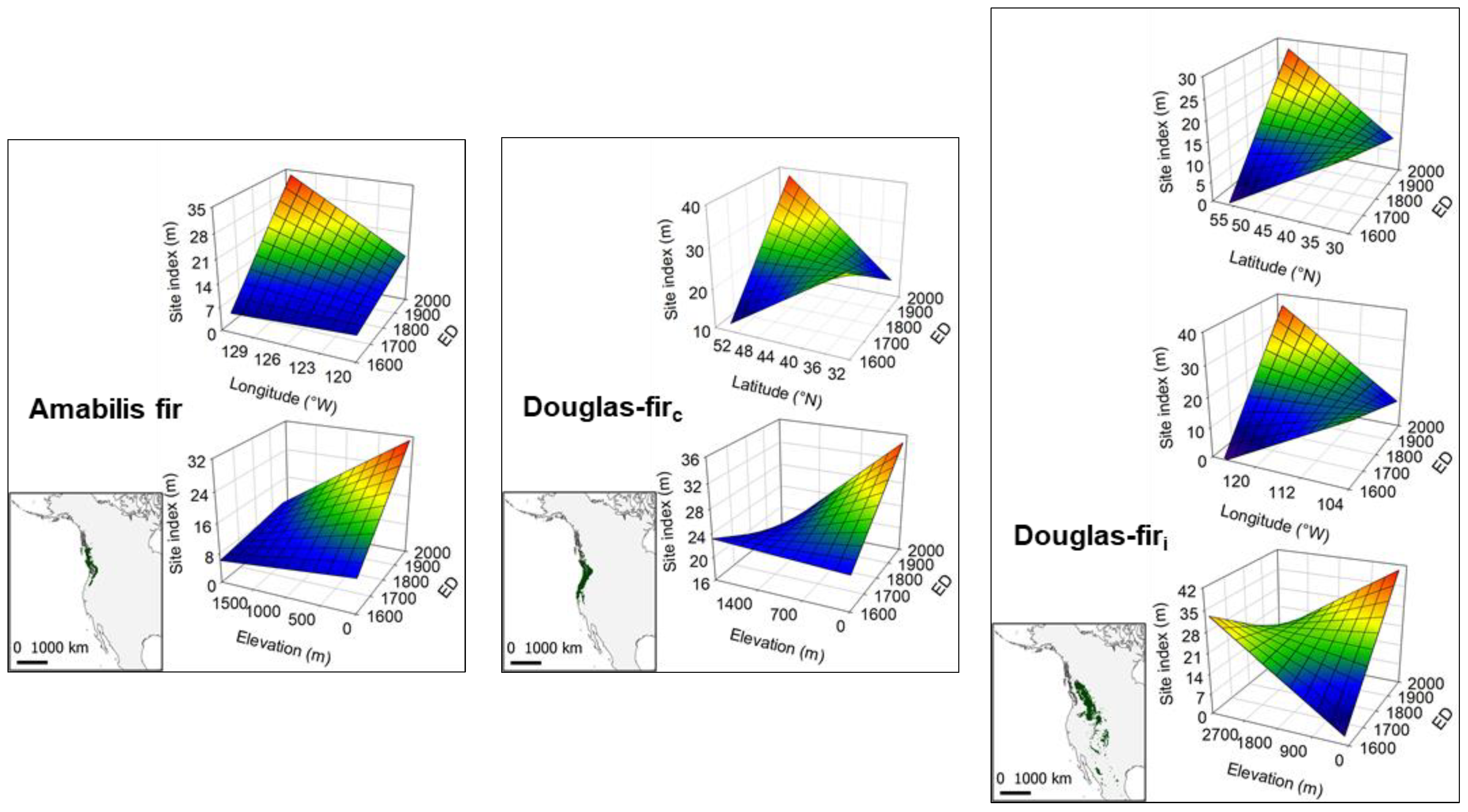{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | N | Latitude | Longitude (W) | Elevation (m) | ED | SI (m) |
|---|---|---|---|---|---|---|
| Alaska-cedar | 445 | 44°24′–58°50′ | 121°30′–136°23′ | 8–1781 | 1601–1940 | 2.78–29.5 |
| Amabilis fir | 791 | 42°56′–55°49′ | 120°37′–130°21′ | 1–1920 | 1604–1968 | 2.02–48.52 |
| Black cottonwood | 99 | 43°33′–61°19′ | 109°05′–153°51′ | 30–2048 | 1783–1965 | 8.83–41.60 |
| Douglas-fir | 13,987 | 31°24′–55°02′ | 104°19′–127°26′ | 5–3477 | 1600–1966 | 2.44–53.34 |
| Engelmann spruce | 3899 | 32°42′–56°59′ | 104°56′–128°23′ | 610–3687 | 1634–1964 | 2.85–32.24 |
| Grand fir | 1345 | 39°06′–50°14′ | 113°47′–125°24′ | 25–2103 | 1739–1966 | 6.10–59.89 |
| Lodgepole pine | 6784 | 34°07′–62°08′ | 104°58′–136°14′ | 11–3612 | 1604–1966 | 2.13–32.92 |
| Mountain hemlock | 1825 | 37°09′–61°06′ | 114°36′–150°36′ | 7–3150 | 1600–1960 | 2.01–27.01 |
| Ponderosa pine | 6838 | 31°47′–51°12′ | 99°31′–124°03′ | 91–3036 | 1625–1967 | 4.07–49.53 |
| Red alder | 135 | 40°45′–57°59′ | 121°09′–135°28′ | 5–1072 | 1781–1963 | 14.54–42.31 |
| Sitka spruce | 922 | 40°34′–61°07′ | 121°47′–153°11′ | 1–1255 | 1603–1962 | 2.31–57.91 |
| Subalpine fir | 5296 | 33°36′–61°12′ | 105°06′–133°04′ | 100–3627 | 1624–1967 | 1.82–40.39 |
| Western hemlock | 3479 | 37°55′–61°04′ | 114°29′–148°04′ | 1–2896 | 1601–1967 | 2.33–43.28 |
| Western larch | 1006 | 44°18′–50°50′ | 112°51′–121°16′ | 455–2286 | 1617–1964 | 7.2–34.14 |
| Western redcedar | 1424 | 43°35′–58°20′ | 113°44′–135°06′ | 1–1783 | 1601–1962 | 3.52–36.86 |
| Western white pine | 165 | 36°18′–52°13′ | 113°50′–124°28′ | 91–3018 | 1659–1963 | 5.57–36.58 |
| Total | 48,440 | 31°24′–62°08′ | 99°31′–153°51′ | 1–3687 | 1600–1968 | 1.82–59.89 |
| Species | AN_T (°C) | JJA_T (°C) | PDSI_AN | PDSI_JJA | ED |
|---|---|---|---|---|---|
| Alaska-cedar | 0.195 ** | 0.266 ** | 0.296 ** | 0.164 * | −0.255 ** |
| Amabilis fir | 0.554 ** | 0.552 ** | −0.115 * | −0.020 | 0.478 ** |
| Black cottonwood | 0.366 ** | 0.299 * | 0.309 * | 0.286 * | 0.274 * |
| Douglas-firc | −0.044 ** | −0.239 ** | 0.183 ** | 0.143 ** | 0.182 ** |
| Douglas-firi | 0.307 ** | 0.225 ** | 0.086 ** | 0.108 ** | 0.175 ** |
| Engelmann spruce | 0.459 ** | 0.452 ** | −0.026 | 0.006 | 0.157 ** |
| Grand firc | 0.052 | 0.085 | 0.138 | 0.120 | 0.214 * |
| Grand firi | 0.172 ** | −0.011 | 0.084 * | 0.090 * | 0.040 |
| Lodgepole pine | 0.375 ** | 0.376 ** | 0.047 ** | 0.042 ** | 0.336 ** |
| Mountain hemlockc | 0.121 ** | 0.186 ** | 0.056 | −0.142 ** | 0.254 ** |
| Mountain hemlocki | 0.401 ** | 0.401 ** | 0.118 | 0.164 | 0.323 ** |
| Ponderosa pine | 0.207 ** | −0.145 ** | 0.099 ** | 0.094 ** | 0.008 |
| Red alder | 0.062 | −0.057 | 0.048 | 0.022 | 0.161 |
| Sitka spruce | 0.437 ** | 0.417 ** | −0.030 | −0.105 * | 0.433 ** |
| Subalpine fir | 0.394 ** | 0.336 ** | −0.044 * | 5 × 10−4 | 0.306 ** |
| Western larch | 0.270 ** | 0.231 ** | 0.049 | 0.040 | 0.203 ** |
| Western hemlockc | 0.528 ** | 0.450 ** | −0.038 | 0.025 | 0.487 ** |
| Western hemlocki | 0.271 ** | 0.272 ** | −0.010 | 0.077 | 0.390 ** |
| Western redcedarc | 0.308 ** | 0.423 ** | −0.191 ** | −0.070 | 0.463 ** |
| Western redcedari | 0.103 * | 0.127 * | −0.104 * | −0.073 | 0.286 ** |
| Western white pinec | 0.195 | 0.224 | 0.171 | 0.173 | 0.135 |
| Western white pinei | 0.210 | 0.255 | −0.029 | −0.029 | 0.313 * |
| Total | 0.508 ** | 0.342 ** | 0.071 ** | 0.084 ** | 0.235 ** |
| Fixed Effects | F-Value | p-Value | Random Effects | Variance | SD | Total Effects |
|---|---|---|---|---|---|---|
| R2 = 0.310 | R2 = 0.490 | |||||
| AN_T | 2125.83 | <0.001 | Species | 14.04 | 3.75 | |
| PDSI_AN | 141.04 | <0.001 | Residual | 40.11 | 6.33 | |
| Leaf form | 122.85 | <0.001 | ||||
| ED | 95.76 | <0.001 | ||||
| Groups | 24.27 | <0.001 | ||||
| Shade tolerance | 19.02 | <0.001 | ||||
| Slope | 7.75 | <0.001 | ||||
| C/N | 4.07 | 0.044 | ||||
| Groups*ED | 38.62 | <0.001 | ||||
| Leaf form*ED | 16.35 | <0.001 | ||||
| Shade tolerance*ED | 14.13 | <0.001 | ||||
| Slope*ED | 7.52 | <0.001 | ||||
| C/N*ED | 5.27 | 0.022 |
| Species | Lat. × ED | Lon. × ED | Ele. × ED | Slope × ED | C/N × ED | R2 |
|---|---|---|---|---|---|---|
| Alaska-cedar | 1.14 | 1.61 | 0.05 | 0.83 | 0.57 | 0.221 |
| Amabilis fir | 2.47 | 10.23 * | 34.04 ** | 1.11 | 0.35 | 0.606 |
| Black cottonwood | 2.35 | 0.27 | 3 × 10−3 | 0.36 | 8.70 * | 0.495 |
| Douglas-firc | 103.15 ** | 3.20 | 5.84 | 0.52 | 10.80 * | 0.335 |
| Douglas-firi | 356.44 ** | 119.94 ** | 31.82 ** | 4.71 * | 21.20 ** | 0.466 |
| Engelmann spruce | 29.42 ** | 1.19 | 29.17 ** | 2.44 | 0.06 | 0.378 |
| Grand firc | 1.49 | 1.84 | 0.03 | 0.68 | 0.01 | 0.235 |
| Grand firi | 1.79 | 3.66 | 2.37 | 3.91 | 0.13 | 0.449 |
| Lodgepole pine | 204.74 ** | 104.44 ** | 3.23 | 10.14 ** | 8.23 * | 0.403 |
| Mountain hemlockc | 30.85 ** | 21.26 ** | 19.84 ** | 3.61 | 1.24 | 0.355 |
| Mountain hemlocki | 0.61 | 0.41 | 0.87 | 0.46 | 0.19 | 0.239 |
| Ponderosa pine | 26.61 ** | 8.10 * | 65.24 ** | 0.39 | 29.29 ** | 0.613 |
| Red alder | 0.64 | 0.40 | 2.15 | 3.13 | 4 × 10−3 | 0.223 |
| Sitka spruce | 10.12 * | 0.10 | 13.13 ** | 0.08 | 0.08 | 0.379 |
| Subalpine fir | 87.52 ** | 42.98 ** | 0.30 | 1.74 | 0.02 | 0.352 |
| Western larch | 25.81 ** | 1.12 | 0.27 | 1.52 | 1.36 | 0.243 |
| Western hemlockc | 8.12 * | 13.83 ** | 133.63 ** | 5.48 * | 0.41 | 0.446 |
| Western hemlocki | 1.64 | 6.89 * | 8.67 * | 1.05 | 0.35 | 0.267 |
| Western redcedarc | 32.41 ** | 4.48 | 2.59 | 0.04 | 0.37 | 0.352 |
| Western redcedari | 0.03 | 0.02 | 1.60 | 1.40 | 0.06 | 0.242 |
| Western white pinec | 3.23 | 1.47 | 0.44 | 0.46 | 2.72 | 0.309 |
| Western white pinei | 0.02 | 0.62 | 0.47 | 0.05 | 0.60 | 0.317 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Messaoud, Y.; Reid, A.; Tchebakova, N.M.; Goldman, J.A.; Hofgaard, A. The Historical Complexity of Tree Height Growth Dynamic Associated with Climate Change in Western North America. Forests 2022, 13, 738. https://doi.org/10.3390/f13050738
Messaoud Y, Reid A, Tchebakova NM, Goldman JA, Hofgaard A. The Historical Complexity of Tree Height Growth Dynamic Associated with Climate Change in Western North America. Forests. 2022; 13(5):738. https://doi.org/10.3390/f13050738
Chicago/Turabian StyleMessaoud, Yassine, Anya Reid, Nadezhda M. Tchebakova, Jack A. Goldman, and Annika Hofgaard. 2022. "The Historical Complexity of Tree Height Growth Dynamic Associated with Climate Change in Western North America" Forests 13, no. 5: 738. https://doi.org/10.3390/f13050738
APA StyleMessaoud, Y., Reid, A., Tchebakova, N. M., Goldman, J. A., & Hofgaard, A. (2022). The Historical Complexity of Tree Height Growth Dynamic Associated with Climate Change in Western North America. Forests, 13(5), 738. https://doi.org/10.3390/f13050738