
Auxiliary Seed Treatment Is Necessary to Increase Recruitment of a Critically Endangered Species, Abies beshanzuensis (Pinaceae)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species and Sample Collection
2.2. Thousand-Grain Weight
2.3. Seed Viability Test
2.4. Seed Germination Experiment
2.5. Fine Root Identification and Molecular Analysis
2.6. Statistical Analyses
3. Results
3.1. Seed Mass and Viability
3.2. Seed Germination Percentage
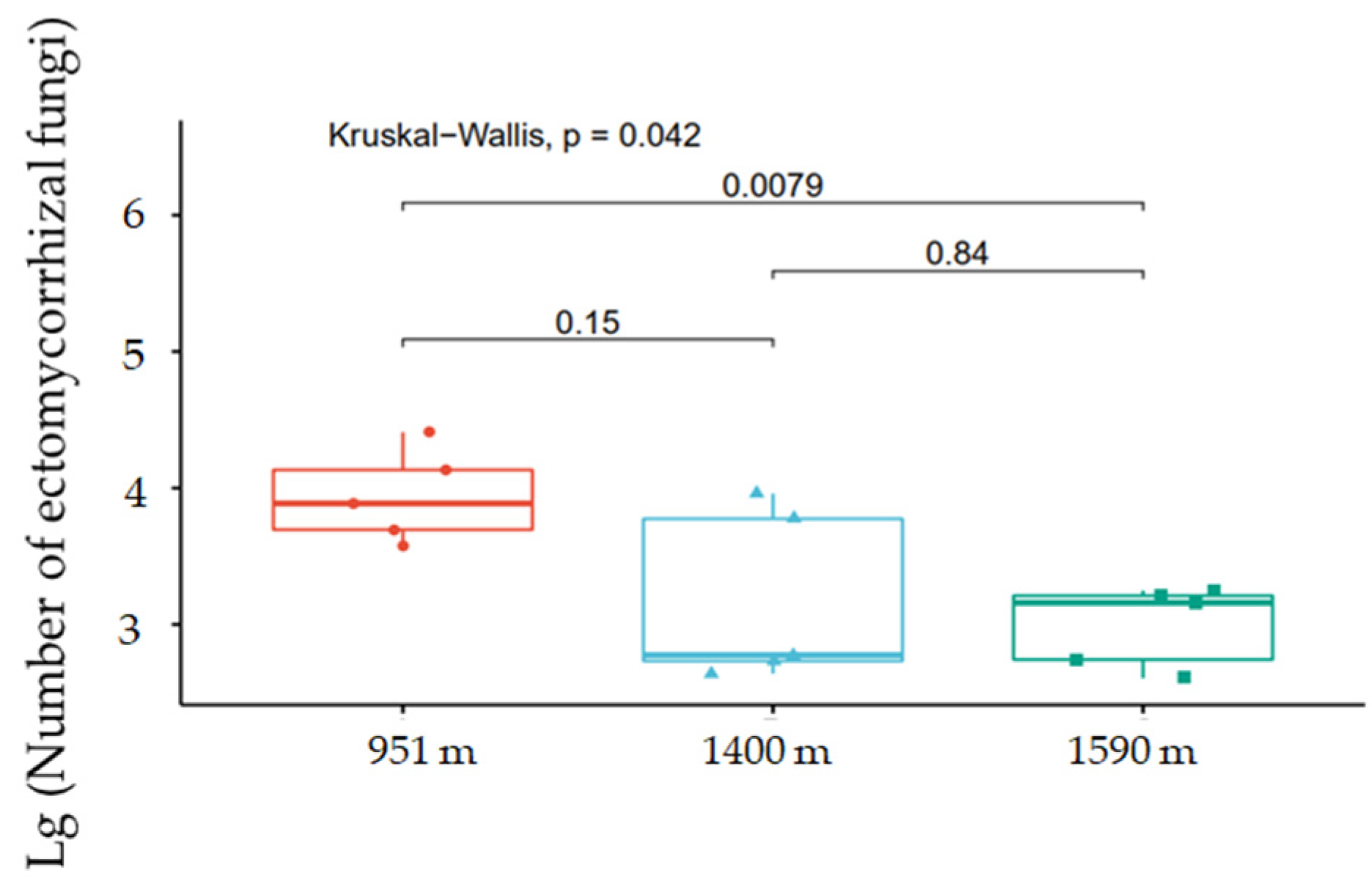
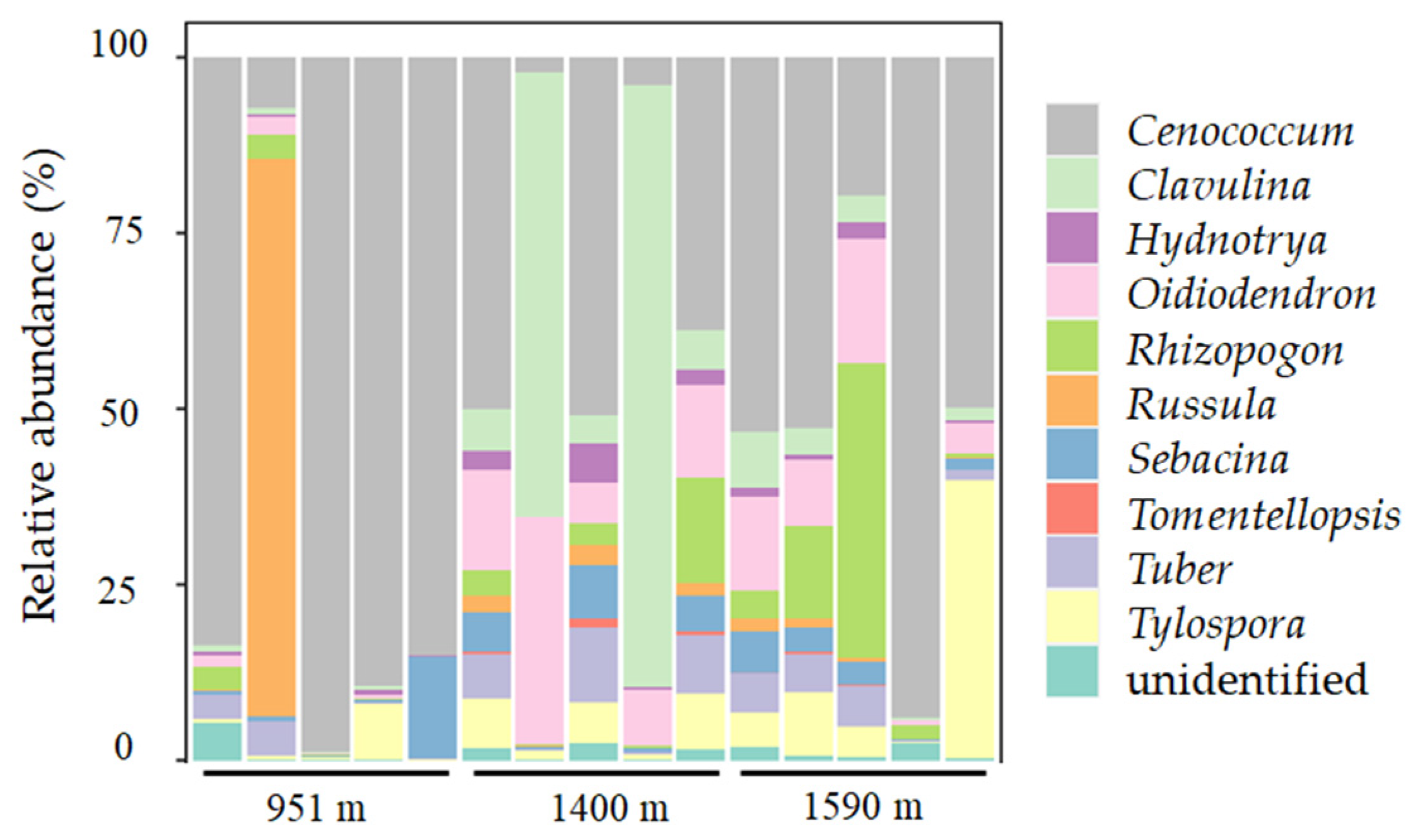
3.3. ECM Fungi Present on Fine Roots of A. beshanzuensis
4. Discussion
4.1. Relationships between Seed Mass and Germination Percentage
4.2. Effects of Warming on Seed Germination in A. beshanzuensis
4.3. ECM Root-Tip Communities
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Volis, S. How to conserve threatened Chinese plant species with extremely small populations? Plant Divers. 2016, 38, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Wade, E.M.; Nadarajan, J.; Yang, X.; Ballesteros, D.; Sun, W.; Pritchard, H.W. Plant species with extremely small populations (PSESP) in China: A seed and spore biology perspective. Plant Divers. 2016, 38, 209–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Cai, L.; Liu, D.; Chen, G.; Gratzfeld, J.; Sun, W. China’s conservation program on Plant Species with Extremely Small Populations (PSESP): Progress and perspectives. Biol. Conserv. 2020, 244, 108535. [Google Scholar] [CrossRef]
- Sha, D.; Yanni, W.; Kunlin, W.; Lin, F.; Lin, L.; Songjun, Z. Breeding characteristics and artificial propagation of 14 species of Wild Plant with Extremely Small Populations (WPESP) in China. Biodivers. Sci. 2020, 28, 385–400. [Google Scholar] [CrossRef]
- Ma, Y.; Chen, G.; Grumbine, R.E.; Dao, Z.; Sun, W.; Guo, H. Conserving plant species with extremely small populations (PSESP) in China. Biodivers. Conserv. 2013, 22, 803–809. [Google Scholar] [CrossRef]
- Fenner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Donohue, K.; De Casas, R.R.; Burghardt, L.; Kovach, K.; Willis, C.G. Germination, Postgermination Adaptation, and Species Ecological Ranges. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 293–319. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: London, UK, 2014. [Google Scholar]
- Koutecká, E.; Lepš, J. Effect of Light and Moisture Conditions and Seed Age on Germination of Three Closely Related Myosotis Species. Folia Geobot. 2009, 44, 109–130. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. (Eds.) Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA; Berlin/Heidelberg, Germany; Dordrecht, The Netherlands; London, UK, 2013; p. 392. [Google Scholar]
- Cochrane, A.; Hoyle, G.L.; Yates, C.J.; Wood, J.; Nicotra, A.B. Predicting the impact of increasing temperatures on seed germination among populations of Western Australian Banksia (Proteaceae). Seed Sci. Res. 2014, 24, 195–205. [Google Scholar] [CrossRef]
- Orrù, M.; Mattana, E.; Pritchard, H.W.; Bacchetta, G. Thermal thresholds as predictors of seed dormancy release and germination timing: Altitude-related risks from climate warming for the wild grapevine Vitis vinifera subsp. sylvestris. Ann. Bot. 2012, 110, 1651–1660. [Google Scholar] [CrossRef] [Green Version]
- Long, R.L.; Gorecki, M.J.; Renton, M.; Scott, J.K.; Colville, L.; Goggin, D.E.; Commander, L.E.; Westcott, D.A.; Cherry, H.; Finch-Savage, W.E. The ecophysiology of seed persistence: A mechanistic view of the journey to germination or demise. Biol. Rev. Camb. Philos. Soc. 2015, 90, 31–59. [Google Scholar] [CrossRef]
- Liu, Y.; El-Kassaby, Y.A. Timing of seed germination correlated with temperature-based environmental conditions during seed development in conifers. Seed Sci. Res. 2014, 25, 29–45. [Google Scholar] [CrossRef]
- Eriksson, O. Seed size variation and its effect on germination and seedling performance in the clonal herb Convallaria majalis. Acta Oecologica 1999, 20, 61–66. [Google Scholar] [CrossRef]
- Kidson, R.; Westoby, M. Seed mass and seedling dimensions in relation to seedling establishment. Oecologia 2000, 125, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Parker, W.C.; Noland, T.L.; Morneault, A.E. The Effects of Seed Mass on Germination, Seedling Emergence, and Early Seedling Growth of Eastern White Pine (Pinus strobus L.). New For. 2006, 32, 33–49. [Google Scholar] [CrossRef]
- Bu, H.; Chen, X.; Xu, X.; Liu, K.; Jia, P.; Du, G. Seed mass and germination in an alpine meadow on the eastern Tsinghai–Tibet plateau. Plant Ecol. 2007, 191, 127–149. [Google Scholar] [CrossRef]
- Hantsch, L.; Bruelheide, H.; Erfmeier, A. High phenotypic variation of seed traits, germination characteristics and genetic diversity of an invasive annual weed. Seed Sci. Res. 2013, 23, 27–40. [Google Scholar] [CrossRef]
- Liu, X.T. Predicting the Suitable Distribution Area of Abies beshanzuensis: To Explor the Potential Dispersion of the Plant Species with Extremely Small Populations. Master’s Thesis, Zhejiang Normal University, Jinhua, China, 2019. [Google Scholar]
- Ai, J.G. Studies on the Endangerment Mechanism and Conservation Strategies for Abies beshanzuensis. Master’s Thesis, Zhejiang University, Hangzhou, China, 2005. [Google Scholar]
- Li, X.-X.; Tao, C.; Wang, Q.-C.; Cui, G.-F. Characteristics of geographic distribution of four critically endangered species of Abies in subtropical China and its relationship with climate. Chin. J. Plant Ecol. 2012, 36, 1154–1164. [Google Scholar] [CrossRef]
- Liu, R.; Wang, C.J.; He, J.; Zhang, Z.X. Analysis of geographical distribution of Abies in China under climate change. Bull. Bot. Res. 2018, 38, 37–46. [Google Scholar]
- He, F. Price of prosperity: Economic development and biological conservation in China. J. Appl. Ecol. 2009, 46, 511–515. [Google Scholar] [CrossRef]
- Rao, L.B. Study on Conservation Genetics of Five Endangered Abies Species by Marker Technology of Biochip. Ph.D. Thesis, Chinese Academy of Forestry, Beijing, China, 2009. [Google Scholar]
- Wu, Y.G.; Rao, L.B.; Chen, D.L.; Ye, Z.L. Artificial seedling raising of Abies beshanzuensis seed. J. Anhui Agric. Sci. 2010, 38, 12038–12039. [Google Scholar] [CrossRef]
- Tang, R.Q.; Li, X.K.; Ou, Z.L.; Li, R.T. The fruiting characteristics and reproductive capacity of seeds of Abies yuanbaoshanenis. Bull. Bot. Res. 2001, 21, 404–408. [Google Scholar]
- Lai, J.S.; Li, Q.M.; Xie, Z.Q. Seed germination characteristic of the endangered plant Abies chensiensis. Acta Phytoecol. Sin. 2003, 27, 661–666. [Google Scholar] [CrossRef] [Green Version]
- Treseder, K.K. A meta-analysis of mycorrhizal responses to nitrogen, phosphorus, and atmospheric CO2 in field studies. New Phytol. 2004, 164, 347–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Classen, A.T.; Sundqvist, M.K.; Henning, J.A.; Newman, G.S.; Moore, J.A.M.; Cregger, M.A.; Moorhead, L.C.; Patterson, C.M. Direct and indirect effects of climate change on soil microbial and soil microbial-plant interactions: What lies ahead? Ecosphere 2015, 6, 1–21. [Google Scholar] [CrossRef]
- Dalong, M.; Luhe, W.; Guoting, Y.; LiQiang, M.; Chun, L. Growth response of Pinus densiflora seedlings inoculated with three indigenous ectomycorrhizal fungi in combination. Braz. J. Microbiol. 2011, 42, 1197–1204. [Google Scholar] [CrossRef] [Green Version]
- Correia, M.; Heleno, R.; Vargas, P.; Rodríguez-Echeverría, S. Should I stay or should I go? Mycorrhizal plants are more likely to invest in long-distance seed dispersal than non-mycorrhizal plants. Ecol. Lett. 2018, 21, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Hayward, J.; Horton, T.R.; Pauchard, A.; Nuñez, M.A. A single ectomycorrhizal fungal species can enable a Pinus invasion. Ecology 2015, 96, 1438–1444. [Google Scholar] [CrossRef]
- Murata, M.; Kanetani, S.; Nara, K. Ectomycorrhizal fungal communities in endangered Pinus amamiana forests. PLoS ONE 2017, 12, e0189957. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Wang, S.; Wei, X.; Jiang, M. Sensitivity of seed germination to temperature of a relict tree species from different origins along latitudinal and altitudinal gradients: Implications for response to climate change. Trees 2019, 33, 1435–1445. [Google Scholar] [CrossRef]
- Teste, F.P.; Karst, J.; Jones, M.D.; Simard, S.W.; Durall, D.M. Methods to control ectomycorrhizal colonization: Effectiveness of chemical and physical barriers. Mycorrhiza 2006, 17, 51–65. [Google Scholar] [CrossRef]
- Veach, A.M.; Yip, D.; Engle, N.L.; Yang, Z.K.; Bible, A.; Morrell-Falvey, J.; Tschaplinski, T.J.; Kalluri, U.C.; Schadt, C.W. Modification of plant cell wall chemistry impacts metabolome and microbiome composition in Populus PdKOR1 RNAi plants. Plant Soil 2018, 429, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Clemmensen, K.E.; Bahr, A.; Ovaskainen, O.; Dahlberg, A.; Ekblad, A.; Wallander, H.; Stenlid, J.; Finlay, R.D.; Wardle, D.A.; Lindahl, B.D. Roots and Associated Fungi Drive Long-Term Carbon Sequestration in Boreal Forest. Science 2013, 339, 1615–1618. [Google Scholar] [CrossRef] [PubMed]
- Bernareggi, G.; Carbognani, M.; Mondoni, A.; Petraglia, A. Seed dormancy and germination changes of snowbed species under climate warming: The role of pre- and post-dispersal temperatures. Ann. Bot. 2016, 118, 529–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walck, J.L.; Hidayati, S.N.; Dixon, K.W.; Thompson, K.; Poschlod, P. Climate change and plant regeneration from seed. Glob. Chang. Biol. 2010, 17, 2145–2161. [Google Scholar] [CrossRef]
- Baird, R.; Stokes, C.E.; Frampton, J.; Smith, B.; Watson, C.; Pilgrim, C.; Scruggs, M. Diversity and density of the EM fungal community present in high elevation Fraser fir forests of Great Smoky Mountains National Park. N. Am. Fungi 2014, 9, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Teste, F.P.; Simard, S.W.; Durall, D.M.; Guy, R.D.; Jones, M.; Schoonmaker, A.L. Access to mycorrhizal networks and roots of trees: Importance for seedling survival and resource transfer. Ecology 2009, 90, 2808–2822. [Google Scholar] [CrossRef]
- Veach, A.M.; Stokes, C.E.; Knoepp, J.; Jumpponen, A.; Baird, R. Fungal communities and functional guilds shift along an elevational gradient in the southern appalachian mountains. Microbial Ecology 2017, 76, 156–168. [Google Scholar] [CrossRef]
- Fernandez, C.; Nguyen, N.H.; Stefanski, A.; Han, Y.; Hobbie, S.E.; Montgomery, R.A.; Reich, P.; Kennedy, P.G. Ectomycorrhizal fungal response to warming is linked to poor host performance at the boreal-temperate ecotone. Glob. Chang. Biol. 2016, 23, 1598–1609. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; May, T.; Smith, M.E. Ectomycorrhizal lifestyle in fungi: Global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 2009, 20, 217–263. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Pec, G.J.; Simard, S.W.; Cahill, J.F.; Karst, J. The effects of ectomycorrhizal fungal networks on seedling establishment are contingent on species and severity of overstorey mortality. Mycorrhiza 2020, 30, 173–183. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, R.; Liu, Y.; Zhang, J.; Xing, H.; Jiang, S.; Liu, Y. Auxiliary Seed Treatment Is Necessary to Increase Recruitment of a Critically Endangered Species, Abies beshanzuensis (Pinaceae). Forests 2022, 13, 961. https://doi.org/10.3390/f13060961
Hu R, Liu Y, Zhang J, Xing H, Jiang S, Liu Y. Auxiliary Seed Treatment Is Necessary to Increase Recruitment of a Critically Endangered Species, Abies beshanzuensis (Pinaceae). Forests. 2022; 13(6):961. https://doi.org/10.3390/f13060961
Chicago/Turabian StyleHu, Rui, Yajing Liu, Jiaxin Zhang, Hua Xing, Sha Jiang, and Yu Liu. 2022. "Auxiliary Seed Treatment Is Necessary to Increase Recruitment of a Critically Endangered Species, Abies beshanzuensis (Pinaceae)" Forests 13, no. 6: 961. https://doi.org/10.3390/f13060961
APA StyleHu, R., Liu, Y., Zhang, J., Xing, H., Jiang, S., & Liu, Y. (2022). Auxiliary Seed Treatment Is Necessary to Increase Recruitment of a Critically Endangered Species, Abies beshanzuensis (Pinaceae). Forests, 13(6), 961. https://doi.org/10.3390/f13060961