
Impact of Selected Environmental Factors on Variation in Leaf and Branch Traits on Endangered Karst Woody Plants of Southwest China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
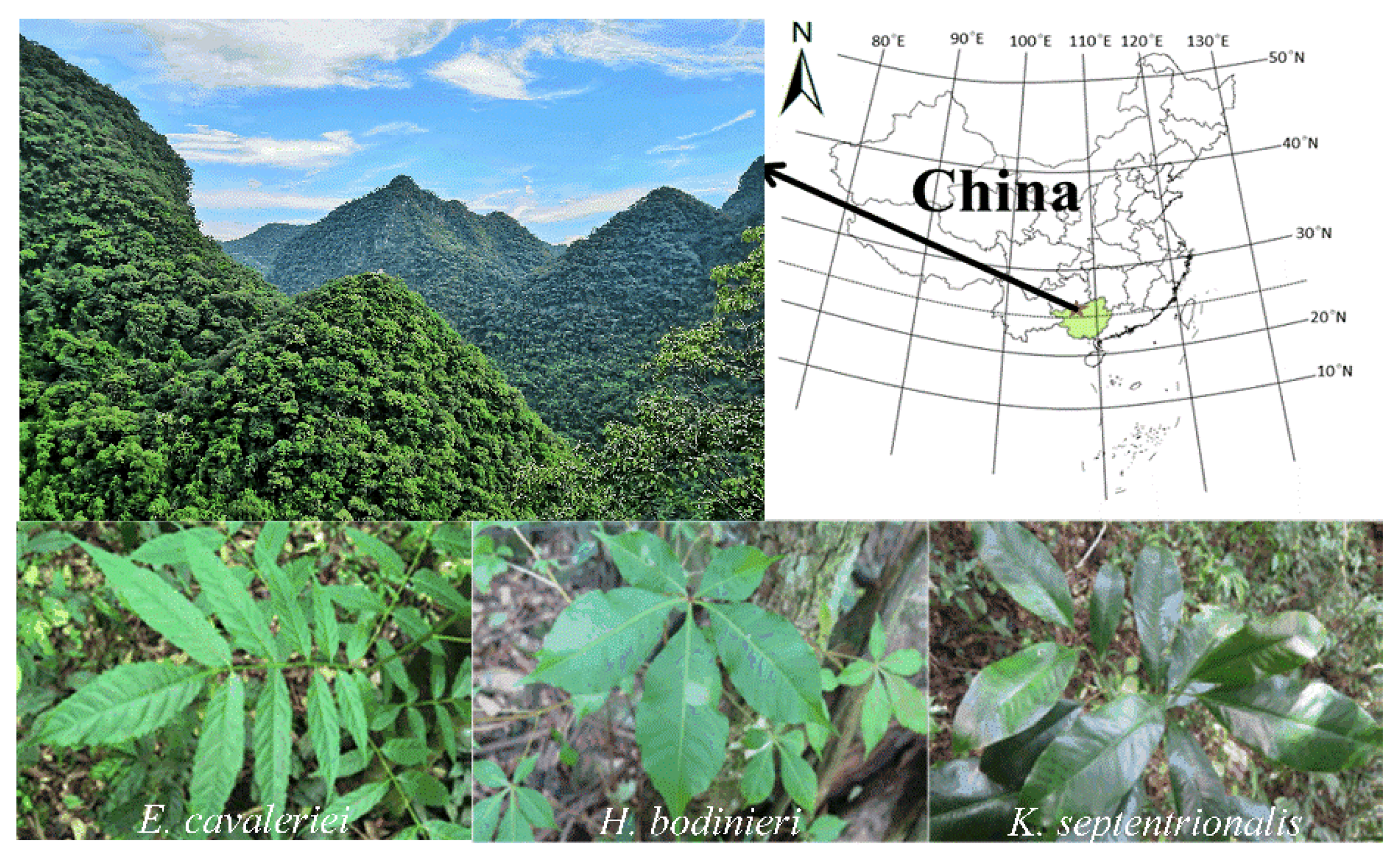
2.1. Research Site
2.2. Field Survey
2.3. Measurement of Leaf and Branch Traits
2.4. Soil Sampling and Measurement
2.5. Statistical Analysis
3. Results
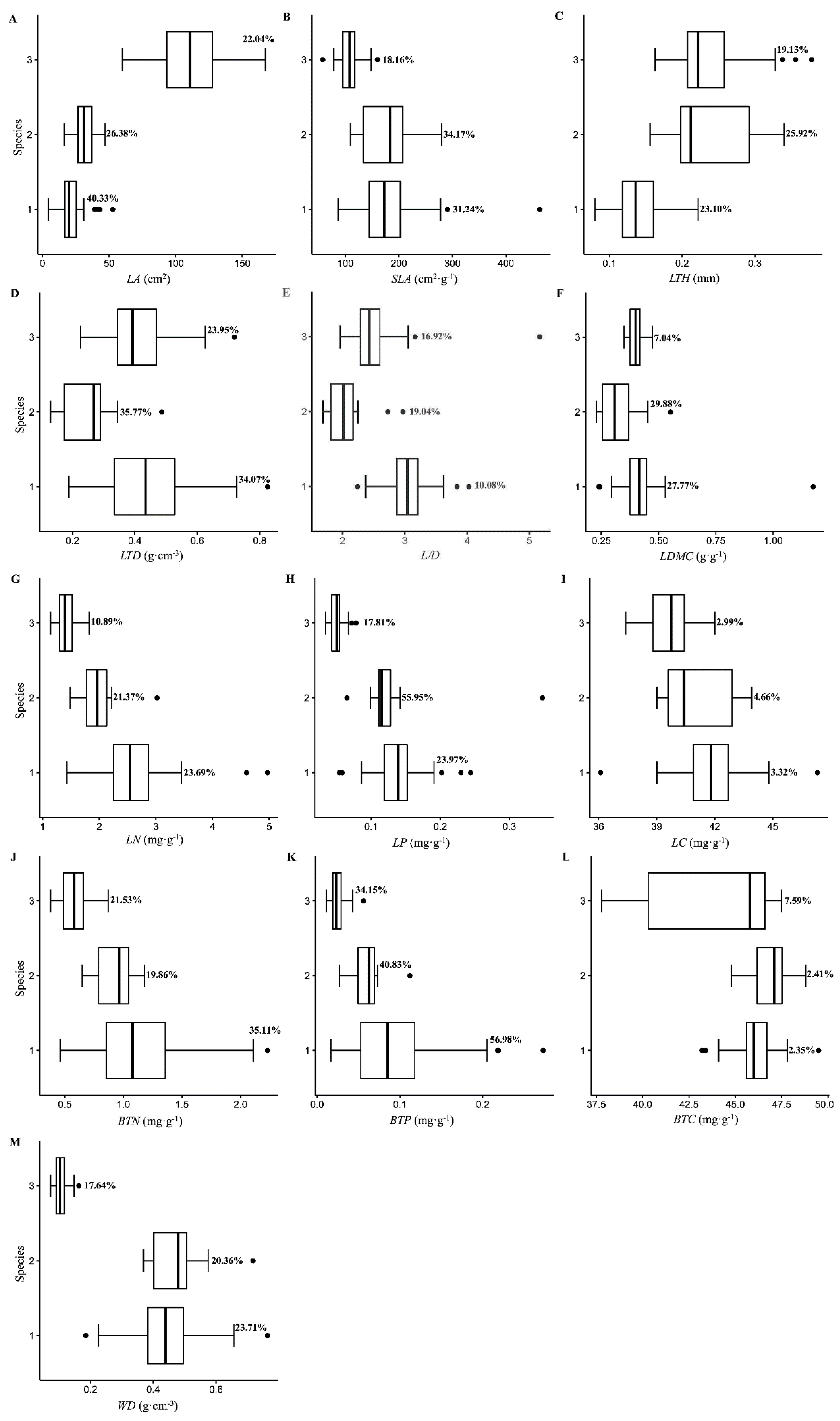
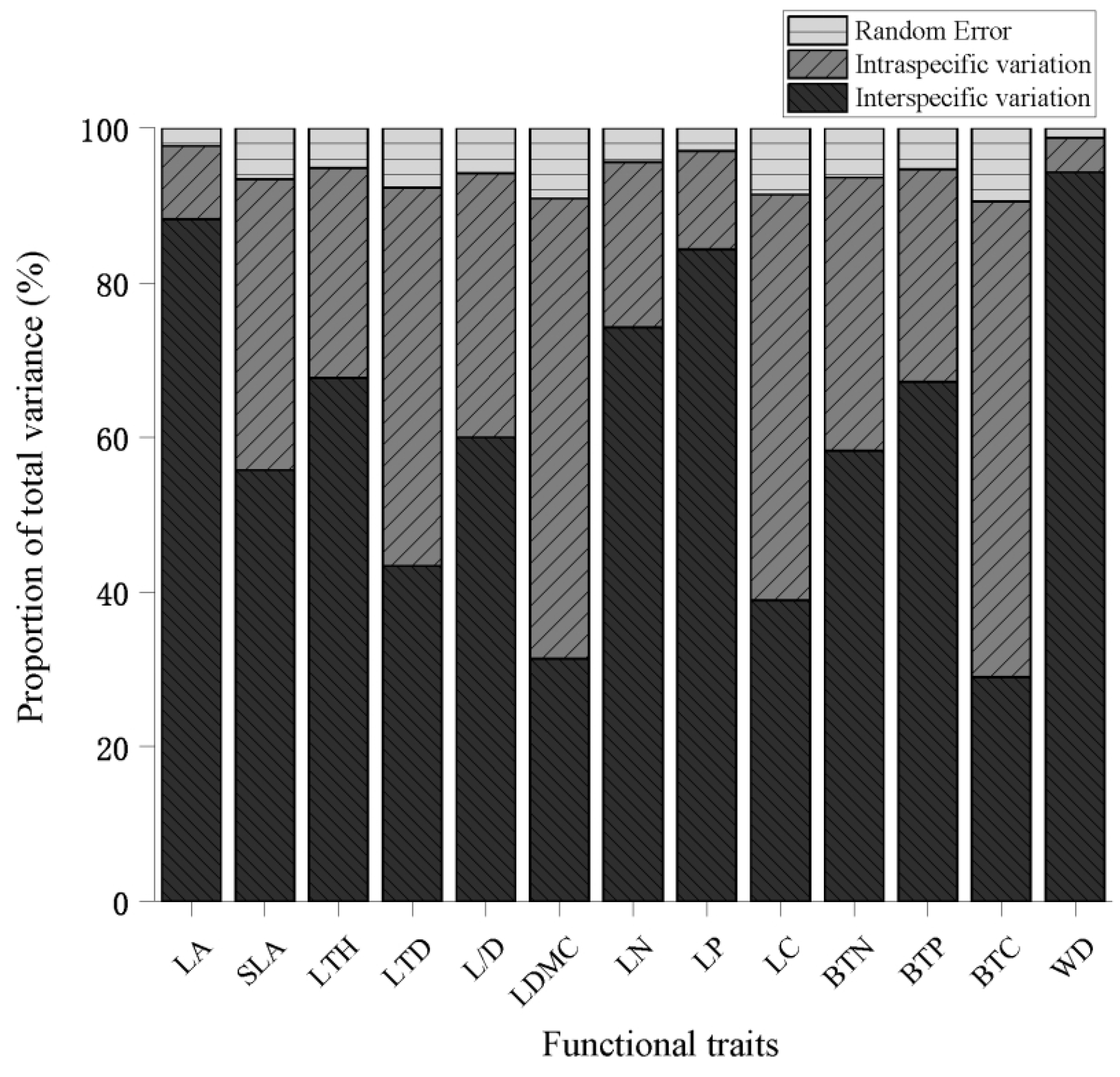
3.1. Variation in the Leaf and Branch Traits of Karst Endangered Plants
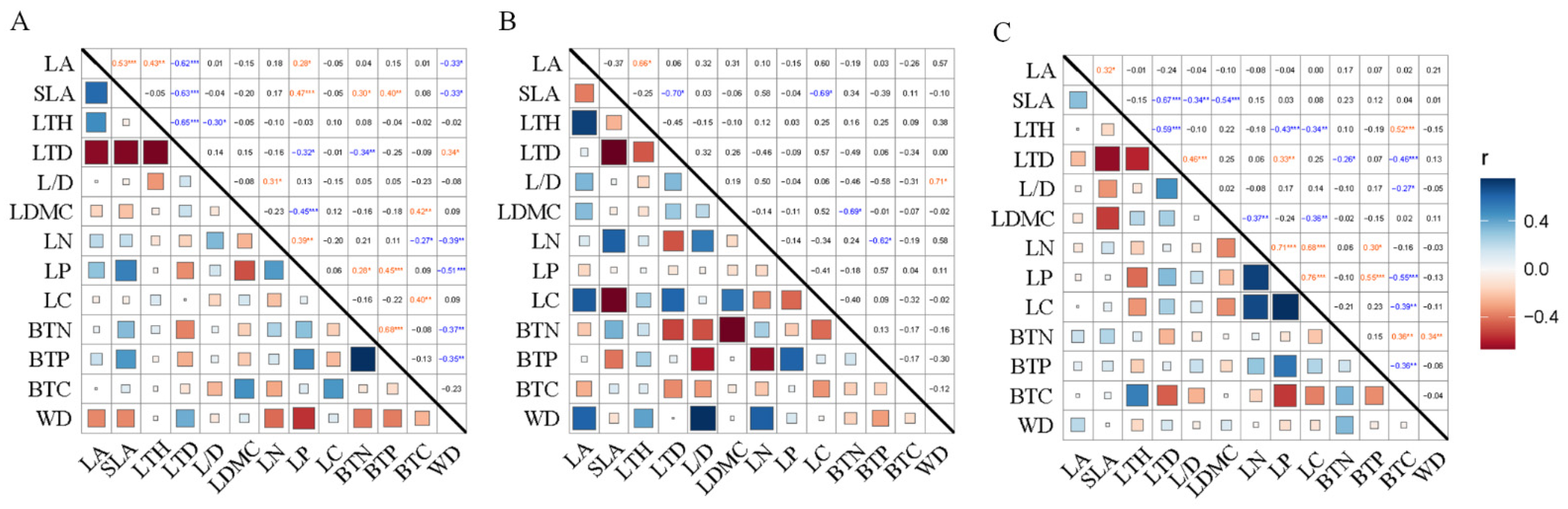
3.2. Trade-Off in the Plant Traits of Karst Endangered Plants
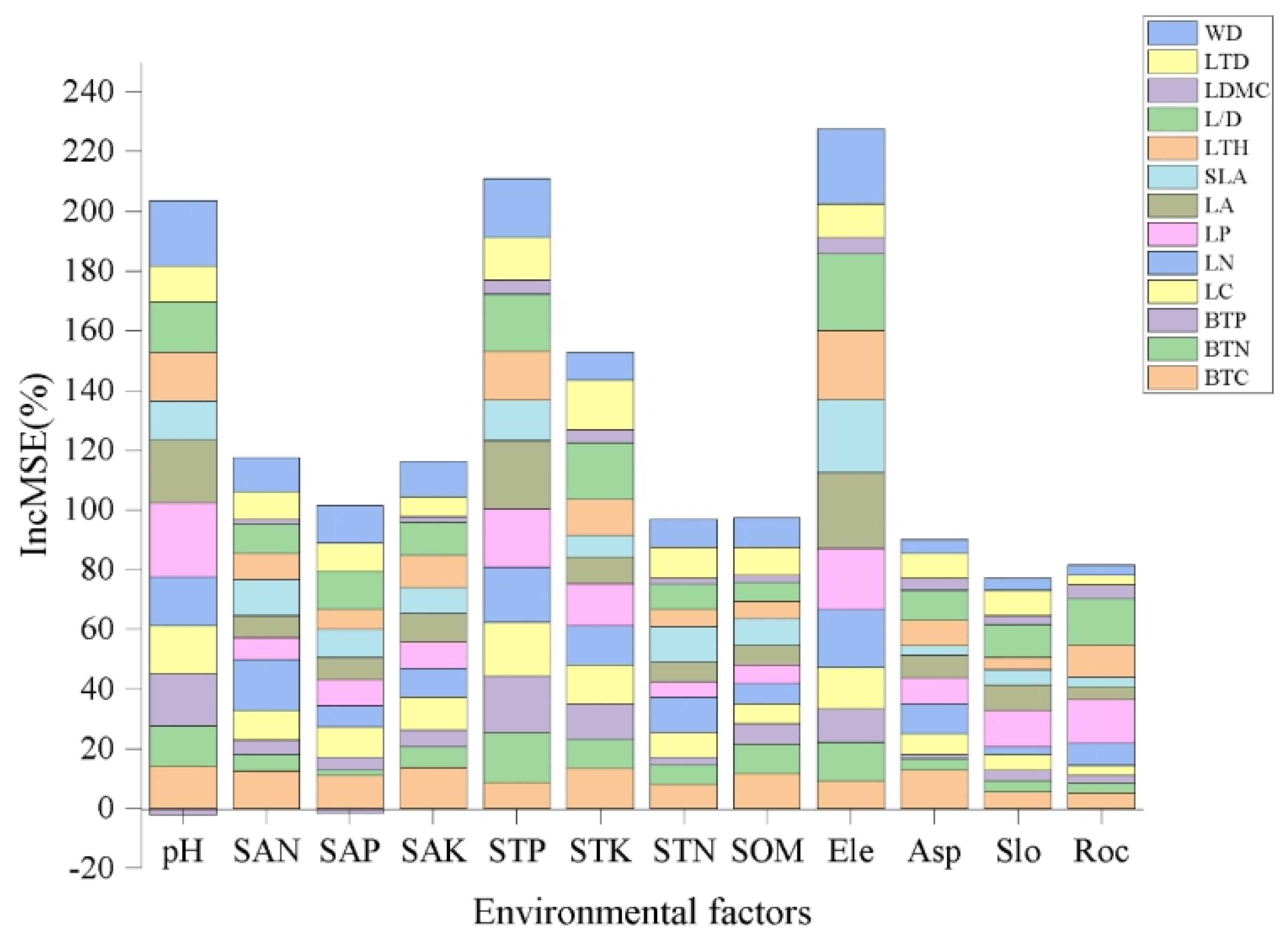
3.3. Relationship between the Karst Environment and Variation in Leaf and Branch Traits
4. Discussion
4.1. Variation in Leaf and Branch Traits in Karst Endangered Plants
4.2. Trade-Offs between Branch and Leaf Traits in Karst Endangered Plants
4.3. Relationship between Endangered Plant Traits and Environmental Factors in the Karst Region
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Meng, T.; Jian, N.I.; Wang, G. Plant functional traits, environments and ecosystem functioning. Acta Phytoecol. Sin. 2007, 31, 150–165. [Google Scholar]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Albert, C.H.; Thuiller, W.; Yoccoz, N.G.; Soudant, A.; Boucher, F.; Saccone, P.; Lavorel, S. Intraspecific functional variability: Extent, structure and sources of variation. J. Ecol. 2010, 98, 604–613. [Google Scholar] [CrossRef]
- Chave, J.; Coomes, D.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a worldwide wood economics spectrum. Ecol. Lett. 2009, 12, 351–366. [Google Scholar] [CrossRef]
- Diaz, S.; Kattge, J.; Cornelissen, J.H.C.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Prentice, I.C.; et al. The global spectrum of plant form and function. Nature 2016, 529, 167. [Google Scholar] [CrossRef]
- Laughlin, D.C. The intrinsic dimensionality of plant traits and its relevance to community assembly. J. Ecol. 2014, 102, 186–193. [Google Scholar] [CrossRef]
- Messier, J.; Lechowicz, M.J.; McGill, B.J.; Violle, C.; Enquist, B.J. Interspecific integration of trait dimensions at local scales: The plant phenotype as an integrated network. J. Ecol. 2017, 105, 1775–1790. [Google Scholar] [CrossRef] [Green Version]
- Verheijen, L.M.; Aerts, R.; Boenisch, G.; Kattge, J.; Van Bodegom, P.M. Variation in trait trade-offs allows differentiation among predefined plant functional types: Implications for predictive ecology. New Phytol. 2016, 209, 563–575. [Google Scholar] [CrossRef]
- Wyka, T.P.; Oleksyn, J.; Zytkowiak, R.; Karolewski, P.; Jagodzinski, A.M.; Reich, P.B. Responses of leaf structure and photosynthetic properties to intra-canopy light gradients: A common garden test with four broadleaf deciduous angiosperm and seven evergreen conifer tree species. Oecologia 2012, 170, 11–24. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Wang, S.; Liu, X.; Nie, M.; Zhou, S.; Hautier, Y. Intra- and interspecific variability of specific leaf area mitigate the reduction of community stability in response to warming and nitrogen addition. Oikos 2022, e09207. [Google Scholar] [CrossRef]
- Pichon, N.A.; Cappelli, S.L.; Allan, E. Intraspecific trait changes have large impacts on community functional composition but do not affect ecosystem function. J. Ecol. 2022, 110, 644–658. [Google Scholar] [CrossRef]
- Gewin, V. Beyond neutrality—Ecology finds its niche. PLoS Biol. 2006, 4, e278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornwell, W.K.; Schwilk, D.W.; Ackerly, D.D. A trait-based test for habitat filtering: Convex hull volume. Ecology 2006, 87, 1465–1471. [Google Scholar] [CrossRef]
- Grime, J.P. Trait convergence and trait divergence in herbaceous plant communities: Mechanisms and consequences. J. Veg. Sci. 2006, 17, 255–260. [Google Scholar] [CrossRef]
- Bhaskar, R.; Dawson, T.E.; Balvanera, P. Community assembly and functional diversity along succession post-management. Funct. Ecol. 2014, 28, 1256–1265. [Google Scholar] [CrossRef]
- Nie, Y.; Chen, H.; Wang, K.; Yang, J. Water source utilization by woody plants growing on dolomite outcrops and nearby soils during dry seasons in karst region of Southwest China. J. Hydrol. 2012, 420, 264–274. [Google Scholar] [CrossRef]
- Zou, Z.; Zeng, F.; Wang, K.; Zeng, Z.; Zhao, L.; Du, H.; Zhang, F.; Zhang, H. Emergy and Economic Evaluation of Seven Typical Agroforestry Planting Patterns in the Karst Region of Southwest China. Forests 2019, 10, 138. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Hu, F.; Zeng, F.; Wang, K.; Peng, W.; Zhang, H.; Zeng, Z.; Zhang, F.; Song, T. Spatial distribution of tree species in evergreen-deciduous broadleaf karst forests in southwest China. Sci. Rep. 2017, 7, 15664. [Google Scholar] [CrossRef] [Green Version]
- Tan, Q.; Wang, W.; Chen, H.; Qin, Z.; Zheng, S.; Zhang, H.; Du, H.; Song, T. Coupling relationships between plant community and soil characteristics in canyon karst region in Southwest China. Bangladesh J. Bot. 2017, 46, 1117–1127. [Google Scholar]
- Peng, Y.; He, Q.; Tan, C.; Shen, W.; Qin, W.; He, F. Quantitative analysis of stand spatial structure of a rare species Kmeria septentrionalis in Guangxi. Chin. J. Ecol. 2016, 35, 363–369. [Google Scholar]
- Song, Y.-B.; Shen-Tu, X.-L.; Dong, M. Intraspecific Variation of Samara Dispersal Traits in the Endangered Tropical Tree Hopea hainanensis (Dipterocarpaceae). Front. Plant Sci. 2020, 11, 599764. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Zeng, F.; Wang, K.; Zeng, Z.; Tang, H.; Zhang, H. Evaluation and Tradeoff Analysis of Ecosystem Service for Typical Land-Use Patterns in the Karst Region of Southwest China. Forests 2020, 11, 451. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Du, H.; Song, T.; Peng, W.; Su, L.; Zhang, H.; Zeng, Z.; Wang, K.; Tang, W.; Wei, L.; et al. Drivers of tree survival in an evergreen-deciduous broadleaf karst forest in southwest China. For. Ecol. Manag. 2021, 499, 119598. [Google Scholar] [CrossRef]
- Su, L.; Du, H.; Zeng, F.; Peng, W.; Rizwan, M.; Nunez-Delgado, A.; Zhou, Y.; Song, T.; Wang, H. Soil and fine roots ecological stoichiometry in different vegetation restoration stages in a karst area, southwest China. J. Environ. Manag. 2019, 252, 109694. [Google Scholar] [CrossRef]
- Wang, M.; Rong, L.; Li, T.; Wang, Q.; Ye, T. Studies on Leaf Functional Traits of Nine Woody Lianas in the Karst Area of Central Guizhou Province. J. Trop. Subtrop. Bot. 2021, 29, 455–464. [Google Scholar]
- Zhang, J.; Li, X.; Han, W.; You, Y.; Jiao, J. Population structure and distribution pattern of Handeliodendron bodinieri in Mulun Natural Reserve, Guangxi. Ecol. Sci. 2009, 28, 495–502. [Google Scholar]
- Liao, M.; Shu, Z.; Wei, X.; Jin, T. Study on Tissue Culture Technique of Eurycorymbus cavaleriei. Seed 2005, 24, 9–11,18. [Google Scholar]
- Lin, Q.; Duan, L.; Yuan, Q. Taxonomic Notes on the Genus Kmeria (Pierre) Dandy (Magnoliaceae). J. Wuhan Bot. Res. 2005, 23, 236–238. [Google Scholar]
- Du, H.; Peng, W.X.; Song, T.Q.; Zeng, F.P.; Wang, K.L.; Song, M.; Zhang, H. Spatial pattern of woody plants and their environmental interpretation in the karst forest of southwest China. Plant Biosyst. 2015, 149, 121–130. [Google Scholar] [CrossRef]
- Bao, S. Soil and Agricultural Chemistry Analysis, 3rd ed.; Agricultural Press: Beijing, China, 2000. [Google Scholar]
- Aerts, R.; Chapin, F.S. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. Adv. Ecol. Res. 1999, 30, 1–67. [Google Scholar]
- Li, J.; Xu, W.; Xiong, G.; Wang, Y.; Zhao, C.; Lu, Z.; Li, Y.; Xie, Z. Leaf nitrogen and phosphorus concentration and the empirical regulations in dominant woody plants of shrublands across southern China. Chin. J. Plant Ecol. 2017, 41, 31–42. [Google Scholar]
- Zhang, X.; Mu, Z.; Lin, M.; Jiang, M.; Gong, J.; You, W. Functional traits of leaves of four dominant plants on Langqi Island, Fuzhou, and factors influencing these traits. Chin. J. Appl. Environ. Biol. 2020, 26, 667–673. [Google Scholar]
- Liu, R.; Liang, S.; Long, W.; Jiang, Y. Variations in Leaf Functional Traits Across Ecological Scales in Riparian Plant Communities of the Lijiang River, Guilin, Southwest China. Trop. Conserv. Sci. 2018, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Paz-Dyderska, S.; Dyderski, M.K.; Szwaczka, P.; Brzezicha, M.; Bigos, K.; Jagodzinski, A.M. Leaf Traits and Aboveground Biomass Variability of Forest Understory Herbaceous Plant Species. Ecosystems 2020, 23, 555–569. [Google Scholar] [CrossRef] [Green Version]
- Rosell, J.A.; Gleason, S.; Mendez-Alonzo, R.; Chang, Y.; Westoby, M. Bark functional ecology: Evidence for tradeoffs, functional coordination, and environment producing bark diversity. New Phytol. 2014, 201, 486–497. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yu, S. Potential tradeoffs between intraspecific and interspecific trait variations along an environmental gradient in a subtropical forest. J. For. Res. 2018, 29, 1731–1740. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, X.; Ma, J.; Liang, S.; Huang, J.; Liu, R.; Pan, Y. Interspecific and intraspecific variation in functional traits of subtropical evergreen and deciduous broadleaved mixed forests in Karst topography, Guilin, Southwest China. Trop. Conserv. Sci. 2016, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.; Huang, Y.; Ding, Y.; Zang, R. Interspecific and intraspecific variation in functional traits of subtropical evergreen and deciduous broad-leaved mixed forests. Biodivers. Sci. 2016, 24, 262–270. [Google Scholar] [CrossRef] [Green Version]
- Auger, S.; Shipley, B. Inter-specific and intra-specific trait variation along short environmental gradients in an old-growth temperate forest. J. Veg. Sci. 2013, 24, 419–428. [Google Scholar] [CrossRef]
- Liu, R.; Bai, J.; Bao, H.; Nong, J.; Zhao, J.; Jiang, Y.; Liang, S.; Li, Y. Variation and correlation in functional traits of main woody plants in the Cyclobalanopsis glauca community in the karst hills of Guilin, southwest China. Chin. J. Plant Ecol. 2020, 44, 828–841. [Google Scholar] [CrossRef]
- Kerkhoff, A.J.; Fagan, W.F.; Elser, J.J.; Enquist, B.J. Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants. Am. Nat. 2006, 168, E103–E122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, I.J.; Ackerly, D.D.; Bongers, F.; Harms, K.E.; Ibarra-Manriquez, G.; Martinez-Ramos, M.; Mazer, S.J.; Muller-Landau, H.C.; Paz, H.; Pitman, N.C.A.; et al. Relationships among ecologically important dimensions of plant trait variation in seven Neotropical forests. Ann. Bot. 2007, 99, 1003–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baraloto, C.; Paine, C.E.T.; Poorter, L.; Beauchene, J.; Bonal, D.; Domenach, A.-M.; Herault, B.; Patino, S.; Roggy, J.-C.; Chave, J. Decoupled leaf and stem economics in rain forest trees. Ecol. Lett. 2010, 13, 1338–1347. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Zhao, Y.; Yang, X.; Shi, Q.; Zhou, L.; Zhang, Q.; Arshad, A.; Yan, E. Geostatistical analysis of spatial variations in leaf traits of woody plants in Tiantong, Zhejiang Province. Chin. J. Plant Ecol. 2016, 40, 48–59. [Google Scholar]
- Cao, J.; Liu, J.; Yuan, Q.; Xu, D.; Fan, H.; Chen, H.; Tan, B.; Liu, L.; Ye, D.; Ni, J. Traits of shrubs in forests and bushes reveal different life strategies. Chin. J. Plant Ecol. 2020, 44, 715–729. [Google Scholar] [CrossRef]
- Zhou, J.; Cieraad, E.; van Bodegom, P.M. Global analysis of trait-trait relationships within and between species. New Phytol. 2022, 233, 1643–1656. [Google Scholar] [CrossRef]
- Penuelas, J.; Fernandez-Martinez, M.; Ciais, P.; Jou, D.; Piao, S.; Obersteiner, M.; Vicca, S.; Janssens, I.A.; Sardans, J. The bioelements, the elementome, and the biogeochemical. Ecology 2019, 100, e02652. [Google Scholar] [CrossRef]
- Werner, F.A.; Homeier, J. Is tropical montane forest heterogeneity promoted by a resource-driven feedback cycle? Evidence from nutrient relations, herbivory and litter decomposition along a topographical gradient. Funct. Ecol. 2015, 29, 430–440. [Google Scholar] [CrossRef]
- Schreeg, L.A.; Santiago, L.S.; Wright, S.J.; Turner, B.L. Stem, root, and older leaf N:P ratios are more responsive indicators of soil nutrient availability than new foliage. Ecology 2014, 95, 2062–2068. [Google Scholar] [CrossRef] [Green Version]
- Courbaud, B.; Vieilledent, G.; Kunstler, G. Intra-specific variability and the competition-colonisation trade-off: Coexistence, abundance and stability patterns. Theor. Ecol. 2012, 5, 61–71. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Adler, P.B.; Godoy, O.; James, E.C.; Fuller, S.; Levine, J.M. Community assembly, coexistence and the environmental filtering metaphor. Funct. Ecol. 2015, 29, 592–599. [Google Scholar] [CrossRef]
- Zeng, Z.; Wang, K.; Liu, X.; Zeng, F.; Song, T.; Peng, W.; Zhang, H.; Du, H. Stoichiometric characteristics of live fresh leaves and leaf litter from typical plant communities in a karst region of northwestern Guangxi, China. Acta Ecol. Sin. 2016, 36, 1907–1914. [Google Scholar]
- Both, S.; Riutta, T.; Paine, C.E.T.; Elias, D.M.O.; Cruz, R.S.; Jain, A.; Johnson, D.; Kritzler, U.H.; Kuntz, M.; Majalap-Lee, N.; et al. Logging and soil nutrients independently explain plant trait expression in tropical forests. New Phytol. 2019, 221, 1853–1865. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Zuo, J.; He, Z.; Wang, Y.; Liu, J.; Liu, W.; Cornelissen, J.H.C. Traits including leaf dry matter content and leaf pH dominate over forest soil pH as drivers of litter decomposition among 60 species. Funct. Ecol. 2019, 33, 1798–1810. [Google Scholar] [CrossRef]
- Siefert, A.; Violle, C.; Chalmandrier, L.; Albert, C.H.; Taudiere, A.; Fajardo, A.; Aarssen, L.W.; Baraloto, C.; Carlucci, M.B.; Cianciaruso, M.V.; et al. A global meta-analysis of the relative extent of intraspecific trait variation in plant communities. Ecol. Lett. 2015, 18, 1406–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Swenson, N.G.; Joseph Wright, S.; Zhang, L.; Song, K.; Du, Y.; Zhang, J.; Mi, X.; Ren, H.; Ma, K. Covariation in Plant Functional Traits and Soil Fertility within Two Species-Rich Forests. PLoS ONE 2012, 7, e34767. [Google Scholar] [CrossRef]
- Reich, P.B. The world-wide ‘fast-slow’ plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Wilson, P.J.; Thompson, K.; Hodgson, J.G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytol. 1999, 143, 155–162. [Google Scholar] [CrossRef]
- Sungpalee, W.; Itoh, A.; Kanzaki, M.; Sri-ngernyuang, K.; Noguchi, H.; Mizuno, T.; Teejuntuk, S.; Hara, M.; Chai-udom, K.; Ohkubo, T.; et al. Intra- and interspecific variation in wood density and fine-scale spatial distribution of stand-level wood density in a northern Thai tropical montane forest. J. Trop. Ecol. 2009, 25, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Violle, C.; Enquist, B.J.; McGill, B.J.; Jiang, L.; Albert, C.H.; Hulshof, C.; Jung, V.; Messier, J. The return of the variance: Intraspecific variability in community ecology. Trends Ecol. Evol. 2012, 27, 244–252. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Mean ± SD | Minimum | Maximum | Coefficient of Variation (%) |
|---|---|---|---|---|
| Leaf area (cm2) | 64.440 ± 47.842 | 4.496 | 167.336 | 74.24 |
| Specific leaf area (cm2·g−1) | 147.120 ± 56.114 | 57.940 | 463.125 | 38.14 |
| Leaf thickness (mm) | 0.193 ± 0.064 | 0.080 | 0.378 | 32.92 |
| Leaf tissue density (g·cm−3) | 0.416 ± 0.134 | 0.130 | 0.825 | 32.23 |
| Leaf length–width ratio | 2.712 ± 0.490 | 1.688 | 5.164 | 18.07 |
| Leaf dry matter content (g·g−1) | 0.402 ± 0.087 | 0.229 | 1.175 | 21.73 |
| Leaf nitrogen concentration (mg·g−1) | 1.992 ± 0.723 | 1.140 | 4.970 | 36.28 |
| Leaf phosphorus concentration (mg·g−1) | 0.098 ± 0.055 | 0.034 | 0.348 | 56.20 |
| Leaf carbon concentration (mg·g−1) | 40.690 ± 1.678 | 36.100 | 44.800 | 4.12 |
| Branch nitrogen concentration (mg·g−1) | 0.875 ± 0.399 | 0.380 | 2.230 | 45.64 |
| Branch phosphorus concentration (mg·g−1) | 0.061 ± 0.052 | 0.011 | 0.273 | 85.06 |
| Branch carbon concentration (mg·g−1) | 45.016 ± 2.697 | 37.800 | 49.500 | 5.99 |
| Wood density (g·cm−3) | 0.288 ± 0.189 | 0.073 | 0.764 | 65.70 |
| Traits | Enviornmental Factors | Wald χ2 | df | Sig. | R2 | p |
|---|---|---|---|---|---|---|
| LA | pH | 20.366 | 1 | 0.000 | 0.92 | <0.05 |
| SAN | 10.007 | 1 | 0.002 | |||
| STP | 9.702 | 1 | 0.002 | |||
| STK | 10.813 | 1 | 0.001 | |||
| Ele | 19.379 | 1 | 0.000 | |||
| Asp | 39.77 | 6 | 0.000 | |||
| Slo | 14.432 | 1 | 0.000 | |||
| SLA | STN | 4.005 | 1 | 0.045 | 0.42 | <0.05 |
| STK | 5.47 | 1 | 0.019 | |||
| Ele | 5.914 | 1 | 0.015 | |||
| LTH | SAN | 5.363 | 1 | 0.021 | 0.62 | <0.05 |
| Asp | 18.636 | 6 | 0.005 | |||
| LTD | SAN | 6.483 | 1 | 0.011 | 0.28 | <0.05 |
| STN | 6.755 | 1 | 0.009 | |||
| STK | 8.663 | 1 | 0.003 | |||
| L/D | SAN | 13.914 | 1 | 0.000 | 0.50 | <0.05 |
| STN | 10.535 | 1 | 0.001 | |||
| STK | 7.857 | 1 | 0.005 | |||
| SOM | 4.304 | 1 | 0.038 | |||
| Asp | 22.074 | 6 | 0.001 | |||
| Slo | 8.346 | 1 | 0.004 | |||
| LDMC | — | — | — | — | 0.11 | NS |
| LN | pH | 5.243 | 1 | 0.022 | 0.66 | <0.05 |
| SAN | 12.446 | 1 | 0.000 | |||
| STN | 1.639 | 1 | 0.200 | |||
| STK | 4.972 | 1 | 0.026 | |||
| Asp | 16.392 | 6 | 0.012 | |||
| LP | SAP | 9.774 | 1 | 0.002 | 0.73 | <0.05 |
| STN | 4.124 | 1 | 0.042 | |||
| STP | 20.765 | 1 | 0.000 | |||
| Ele | 13.888 | 1 | 0.000 | |||
| Asp | 17.929 | 6 | 0.006 | |||
| Roc | 16.881 | 1 | 0.000 | |||
| LC | — | — | — | — | 0.49 | <0.05 |
| BTN | — | — | — | — | 0.47 | <0.05 |
| BTP | STP | 7.963 | 1 | 0.005 | 0.49 | <0.05 |
| BTC | — | — | — | — | 0.17 | NS |
| WD | Ele | 13.737 | 1 | 0.000 | 0.79 | <0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Zeng, F.; Zeng, Z.; Du, H.; Su, L.; Zhang, L.; Lu, M.; Zhang, H. Impact of Selected Environmental Factors on Variation in Leaf and Branch Traits on Endangered Karst Woody Plants of Southwest China. Forests 2022, 13, 1080. https://doi.org/10.3390/f13071080
Zhang C, Zeng F, Zeng Z, Du H, Su L, Zhang L, Lu M, Zhang H. Impact of Selected Environmental Factors on Variation in Leaf and Branch Traits on Endangered Karst Woody Plants of Southwest China. Forests. 2022; 13(7):1080. https://doi.org/10.3390/f13071080
Chicago/Turabian StyleZhang, Chen, Fuping Zeng, Zhaoxia Zeng, Hu Du, Liang Su, Lijin Zhang, Mengzhen Lu, and Hao Zhang. 2022. "Impact of Selected Environmental Factors on Variation in Leaf and Branch Traits on Endangered Karst Woody Plants of Southwest China" Forests 13, no. 7: 1080. https://doi.org/10.3390/f13071080
APA StyleZhang, C., Zeng, F., Zeng, Z., Du, H., Su, L., Zhang, L., Lu, M., & Zhang, H. (2022). Impact of Selected Environmental Factors on Variation in Leaf and Branch Traits on Endangered Karst Woody Plants of Southwest China. Forests, 13(7), 1080. https://doi.org/10.3390/f13071080