
Microbial Residue Distribution in Microaggregates Decreases with Stand Age in Subtropical Plantations

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
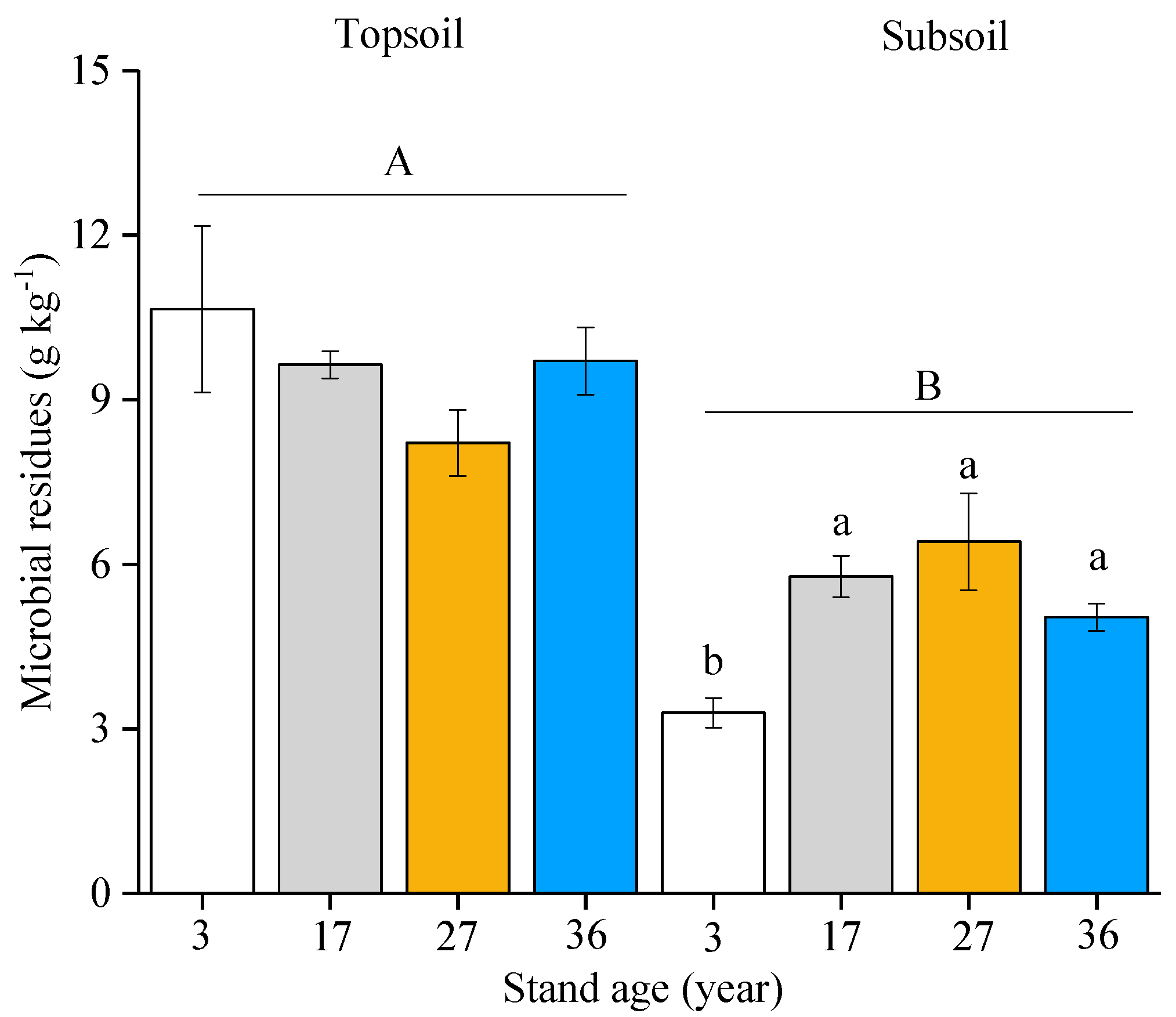
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Angst, G.; Mueller, K.E.; Nierop, K.G.; Simpson, M.J. Plant-or microbial-derived? A review on the molecular composition of stabilized soil organic matter. Soil Biol. Biochem. 2021, 156, 108189. [Google Scholar] [CrossRef]
- Ni, X.; Liao, S.; Tan, S.; Peng, Y.; Wang, D.; Yue, K.; Wu, F.Z.; Yang, Y. The vertical distribution and control of microbial necromass carbon in forest soils. Glob. Ecol. Biogeogr. 2020, 29, 1829–1839. [Google Scholar] [CrossRef]
- Klink, S.; Keller, A.B.; Wild, A.J.; Baumert, V.L.; Gube, M.; Lehndorff, E.; Meyer, N.; Mueller, C.W.; Phillips, R.P.; Pausch, J. Stable isotopes reveal that fungal residues contribute more to mineral-associated organic matter pools than plant residues. Soil Biol. Biochem. 2022, 168, 108634. [Google Scholar] [CrossRef]
- Liang, C.; Amelung, W.; Lehmann, J.; Kästner, M. Quantitative assessment of microbial necromass contribution to soil organic matter. Glob. Change Biol. 2019, 25, 3578–3698. [Google Scholar] [CrossRef] [PubMed]
- Six, H.; Bossuyt, S.; Degryze, K.; Denef, K. A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Li, D.; Niu, S.; Luo, Y. Global patterns of the dynamics of soil carbon and nitrogen stocks following afforestation: A meta-analysis. New Phytol. 2012, 195, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Hu, H.; Sun, W.; Zhu, J.; Liu, G.; Zhou, W.; Zhang, Q.; Shi, P.; Liu, X.; Wu, X.; et al. Effects of national ecological restoration projects on carbon sequestration in China from 2001 to 2010. Proc. Natl. Acad. Sci. USA 2018, 115, 4039–4044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.C.; Liang, M.J.; Wang, S.L. Carbon stock density in planted versus natural Pinus massoniana forests in sub-tropical. China Ann. For. Sci. 2016, 73, 461–472. [Google Scholar]
- State Forestry Administration. General Situation of Forest Resources in China—The 8th National Forest Inventory; State Forestry Administration: Beijing, China, 2014. [Google Scholar]
- Wu, H.; Xiang, W.; Chen, L.; Ouyang, S.; Xiao, W.; Li, S.; Forrester, D.I.; Lei, P.F.; Zeng, Y.L.; Deng, X.W.; et al. Soil phosphorus bioavailability and recycling increased with stand age in Chinese fir plantations. Ecosystems 2020, 23, 973–988. [Google Scholar] [CrossRef]
- Zeng, Y.; Wu, H.; Ouyang, S.; Chen, L.; Fang, X.; Peng, C.; Liu, S.R.; Xiao, W.F.; Xiang, W. Ecosystem service multifunctionality of Chinese fir plantations differing in stand age and implications for sustainable management. Sci. Total Environ. 2021, 788, 147791. [Google Scholar] [CrossRef]
- Chen, G.S.; Yang, Z.J.; Gao, R.; Xie, J.S.; Guo, J.F.; Huang, Z.Q.; Yang, Y.S. Carbon storage in a chronosequence of Chinese fir plantations in southern China. For. Ecol. Manag. 2013, 300, 68–76. [Google Scholar] [CrossRef]
- Selvaraj, S.; Duraisamy, V.; Huang, Z.; Guo, F.; Ma, X. Influence of long-term successive rotations and stand age of Chinese fir (Cunninghamia lanceolata) plantations on soil properties. Geoderma 2017, 306, 127–134. [Google Scholar] [CrossRef]
- He, X.; Huang, Y.; Zhang, Q.; Ye, S.; Wang, S. Distribution of organic carbon fractions in soil aggregates in Chinese fir plantations with different stand ages. Ecol. Process. 2021, 10, 49. [Google Scholar] [CrossRef]
- Jing, Y.; Ding, X.; Zhao, X.; Tian, P.; Xiao, F.; Wang, Q. Non-additive effects of nitrogen and phosphorus fertilization on microbial biomass and residue distribution in a subtropical plantation. Eur. J. Soil Biol. 2022, 108, 103376. [Google Scholar] [CrossRef]
- Xia, Q.; Chen, L.; Xiang, W.; Ouyang, S.; Wu, H.; Lei, P.; Xiao, W.; Li, S.; Zeng, L.; Kuzyakov, Y. Increase of soil nitrogen availability and recycling with stand age of Chinese-fir plantations. For. Ecol. Manag. 2021, 480, 118643. [Google Scholar] [CrossRef]
- Lu, R. Methods of Soil Agricultural Chemistry Analysis; Chinese Agricultural Science and Technology Press: Beijing, China, 2000; pp. 24–26. [Google Scholar]
- Bardgett, R.D.; Hobbs, P.J.; Frostegard, A. Changes in soil fungal:bacterial biomass ratios following reductions in the intensity of management of an upland grassland. Biol. Fertil. Soils 1996, 22, 261–264. [Google Scholar] [CrossRef]
- Wang, Q.; Gao, W.; Bol, R.; Xiao, Q.; Wu, L.; Zhang, W. Microbial regulation of net N mineralization is driven by C, N, P content and stoichiometry. Eur. J. Soil Sci. 2022, 73, e13257. [Google Scholar] [CrossRef]
- Zhang, X.; Amelung, W. Gas chromatographic determination of muramic acid, glucosamine, mannosamine, and galactosamine in soils. Soil Biol. Biochem. 1996, 28, 1201–1206. [Google Scholar] [CrossRef]
- Engelking, B.; Flessa, H.; Joergensen, R.G. Shifts in amino sugar and ergosterol contents after addition of sucrose and cellulose to soil. Soil Biol. Biochem. 2007, 39, 2111–2118. [Google Scholar] [CrossRef]
- Schermelleh-Engel, K.; Moosbrugger, H.; Müller, H. Evaluating the fit of structural equation models: Tests of significance and descriptive goodness-of-fit measures. Methods Psychol. Res. 2003, 8, 23–74. [Google Scholar]
- Jing, Y.L.; Liu, S.R.; Yin, Y.; Deng, J.F.; Liu, Y.Y.; Yan, P.C.; Gou, K.K. Effects of N-fixing tree species (alnus sibirica) on amino sugars in the soils of a larix kaempferi plantation in eastern liaoning province, China. Acta Ecol. Sin. 2018, 38, 2838–2845. [Google Scholar]
- Yang, L.; Chen, S.; Li, Y.; Wang, Q.; Zhong, X.; Yang, Z.; Lin, C.; Yang, Y. Conversion of natural evergreen broadleaved forests decreases soil organic carbon but increases the relative contribution of microbial residue in subtropical China. Forests 2019, 10, 468. [Google Scholar] [CrossRef] [Green Version]
- Jing, Y.; Tian, P.; Wang, Q.; Li, W.; Sun, Z.; Yang, H. Effects of root dominate over aboveground litter on soil microbial biomass in global forest ecosystems. For. Ecosyst. 2021, 8, 38. [Google Scholar] [CrossRef]
- Hu, M.; Zou, B.; Huang, Z.; Wang, S.; Su, X.; Ding, X.; Zheng, G.C.; Chen, H.Y. Fine root biomass and necromass dynamics of Chinese fir plantations and natural secondary forests in subtropical China. For. Ecol. Manag. 2021, 496, 119413. [Google Scholar] [CrossRef]
- Gunina, A.; Kuzyakov, Y. From energy to (soil organic) matter. Glob. Change Biol. 2022, 28, 2169–2182. [Google Scholar] [CrossRef]
- He, H.; Zhang, W.; Zhang, X.; Xie, H.; Zhuang, J. Temporal responses of soil microorganisms to substrate addition as indicated by amino sugar differentiation. Soil Biol. Biochem. 2011, 43, 1155–1161. [Google Scholar] [CrossRef]
- Indorf, C.; Dyckmans, J.; Joergensen, R.G. Short-term changes in amino sugarspecific d 13 C values after application of C4 and C3 sucrose. Soil Biol. Biochem. 2015, 91, 92–98. [Google Scholar] [CrossRef]
- Zhou, L.; Shalom, A.D.D.; Wu, P.; Li, S.; Jia, Y.; Ma, X. Litterfall production and nutrient return in different-aged Chinese fir (Cunninghamia lanceolata) plantations in South China. J. For. Res. 2015, 26, 79–89. [Google Scholar] [CrossRef]
- Sattar, A.; Naveed, M.; Ali, M.; Zahir, Z.A.; Nadeem, S.M.; Yaseen, M.; Meena, V.S.; Farooq, M.; Singh, R.; Rahman, M.; et al. Perspectives of potassium solubilizing microbes in sustainable food production system: A review. Appl. Soil Ecol. 2019, 133, 146–159. [Google Scholar] [CrossRef]
- Ma, Q.; Scanlan, C.; Bell, R.; Brennan, R. The dynamics of potassium uptake and use, leaf gas exchange and root growth throughout plant phenological development and its effects on seed yield in wheat (triticum aestivum) on a low-k sandy soil. Plant Soil 2013, 373, 373–384. [Google Scholar] [CrossRef]
- Li, P.; Wu, M.; Kang, G.; Zhu, B.; Li, H.; Hu, F.; Jiao, J. Soil quality response to organic amendments on dryland red soil in subtropical China. Geoderma 2020, 373, 114416. [Google Scholar] [CrossRef]
- Huang, Y.; Liang, C.; Duan, X.; Chen, H.; Li, D. Variation of microbial residue contribution to soil organic carbon sequestration following land use change in a subtropical karst region. Geoderma 2019, 353, 340–346. [Google Scholar] [CrossRef]
- Watteau, F.; Villemin, G.; Burtin, G.; Jocteur-Monrozier, L. Root impact on the stability and types of micro-aggregates in silty soil under maize. Eur. J. Soil Sci. 2006, 57, 247–257. [Google Scholar] [CrossRef]
- Zhu, X.M.; Zhang, Z.L.; Wang, Q.T.; Peñuelas, J.; Sardans, J.; Lambers, H.; Li, N.; Liu, Q.; Yin, H.J.; Liu, Z.F. More soil organic carbon is sequestered through the mycelium-pathway than through the root-pathway under nitrogen enrichment in an alpine forest. Glob. Change Biol. 2022, 28, 4947–4961. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Stand Age (year) | Depth (cm) | SOC (g kg−1) | C/N | C/P | AK (mg kg−1) | pH | EC (mg kg−1) | Soil Aggregate Distribution (%) | Bacteria (mg kg−1) | Fungi (mg kg−1) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LMA | SMA | MA | ||||||||||
| 3 | Topsoil | 20.4 ± 0.6 | 11.9 ± 0.4 | 73.5 ± 1.5a | 56.3 ± 2.0 | 4.84 ± 0.01 | 311.0 ± 2.5 | 48.2 ± 0.5 | 41.7 ± 0.6 | 10.1 ± 0.9 | 6.5 ± 0.9 | 0.4 ± 0.0 |
| 17 | 20.1 ± 1.4 | 12.1 ± 0.5 | 36.5 ± 1.7b | 38.1 ± 7.0 | 5.16 ± 0.11 | 255.5 ± 15.2 | 44.9 ± 0.2 | 42.5 ± 2.3 | 12.6 ± 0.4 | 7.8 ± 0.5 | 0.4 ± 0.0 | |
| 27 | 19.5 ± 1.9 | 11.7 ± 1.1 | 34.5 ± 4.4b | 52.1 ± 5.2 | 4.86 ± 0.05 | 349.0 ± 3.5 | 48.3 ± 3.1 | 43.0 ± 0.5 | 8.7 ± 0.8 | 7.6 ± 0.9 | 0.6 ± 0.1 | |
| 36 | 20.9 ± 1.1 | 11.7 ± 0.5 | 40.1 ± 5.0b | 49.2 ± 6.6 | 4.65 ± 0.04 | 320.0 ± 29.2 | 52.2 ± 0.7 | 39.6 ± 0.6 | 8.2 ± 0.4 | 7.2 ± 0.8 | 0.5 ± 0.1 | |
| 3 | Subsoil | 8.5 ± 1.3 | 8.0 ± 1.0b | 16.7 ± 3.8 | 29.8 ± 2.5 | 4.30 ± 0.00 | 246.3 ± 21.1 | 51.3 ± 1.6 | 40.0 ± 1.7 | 8.7 ± 0.4 | 4.5 ± 0.7 | 0.4 ± 0.1 |
| 17 | 14.4 ± 2.2 | 10.6 ± 0.4a | 25.7 ± 8.8 | 28.0 ± 3.03 | 4.26 ± 0.05 | 221.0 ± 7.47 | 64.1 ± 3.3 | 28.3 ± 3.5 | 7.7 ± 0.4 | 6.4 ± 0.8 | 0.4 ± 0.1 | |
| 27 | 11.5 ± 1.6 | 6.7 ± 0.7b | 17.4 ± 3.8 | 38.7 ± 4.4 | 4.26 ± 0.03 | 270.5 ± 8.6 | 51.2 ± 1.2 | 41.7 ± 1.2 | 7.2 ± 0.2 | 6.1 ± 1.1 | 0.0 ± 0.1 | |
| 36 | 10.7 ± 0.8 | 8.5 ± 0.4b | 18.3 ± 1.4 | 28.8 ± 1.3 | 4.3 ± 0.04 | 249.9 ± 10.9 | 49.2 ± 1.2 | 43.3 ± 0.5 | 7.5 ± 0.8 | 7.6 ± 0.8 | 0.0 ± 0.0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, Y.; Zhao, X.; Liu, S.; Tian, P.; Sun, Z.; Chen, L.; Wang, Q. Microbial Residue Distribution in Microaggregates Decreases with Stand Age in Subtropical Plantations. Forests 2022, 13, 1145. https://doi.org/10.3390/f13071145
Jing Y, Zhao X, Liu S, Tian P, Sun Z, Chen L, Wang Q. Microbial Residue Distribution in Microaggregates Decreases with Stand Age in Subtropical Plantations. Forests. 2022; 13(7):1145. https://doi.org/10.3390/f13071145
Chicago/Turabian StyleJing, Yanli, Xuechao Zhao, Shengen Liu, Peng Tian, Zhaolin Sun, Longchi Chen, and Qingkui Wang. 2022. "Microbial Residue Distribution in Microaggregates Decreases with Stand Age in Subtropical Plantations" Forests 13, no. 7: 1145. https://doi.org/10.3390/f13071145
APA StyleJing, Y., Zhao, X., Liu, S., Tian, P., Sun, Z., Chen, L., & Wang, Q. (2022). Microbial Residue Distribution in Microaggregates Decreases with Stand Age in Subtropical Plantations. Forests, 13(7), 1145. https://doi.org/10.3390/f13071145