
Analysis of the NAC Gene Family in Salix and the Identification of SpsNAC005 Gene Contributing to Salt and Drought Tolerance



Abstract
:1. Introduction
2. Materials and Methods
2.1. Salix NAC Gene Family Analysis and Functional Prediction of SpsNAC005 Gene
2.1.1. Bioinformatics Analysis of Genes in Salix
2.1.2. Evolutionary Tree Construction of Genes in Salix and Arabidopsis
2.1.3. Subfamilies Analysis and Functional Prediction of SpsNAC005
2.2. Experimental Materials and Vector Construction
2.3. Agrobacterium-Mediated Transformation of P. hopeiensis Leaf Discs and PCR Assay
2.4. Tissue Expression Specificity of SpsNAC005 in Salix psammophila and Relative Expression Level of SpsNAC005 in Overexpression Lines
2.5. Determination of Salt and Drought Stress and Related Indices in Overexpression Lines
2.5.1. Transplanting before Stress and Determination of Related Growth Indices
2.5.2. Salt and Drought Treatments and Determination of SOD, POD, MDA, and Pro
2.5.3. Expression Analysis of the SOS1, MPK6, HKT1, P5CS1, and PRODH1 Genes under Drought and Salt Stress
2.6. Statistical Analysis


3. Results
3.1. Identification of NAC Genes Obtained by Cloning in Salix psammophila
3.2. Classification of Salix NAC Gene Family Members and Phylogenetic Tree Analysis
3.3. Functional Prediction of SpsNAC005
3.4. Tissue-Specific Expression of the SpsNAC005 Gene in Salix psammophila
3.5. PCR Identification and Expression Analysis of Transgenic P. hopeiensis
3.6. Phenotypic Analysis of Transgenic P. hopeiensis Lines
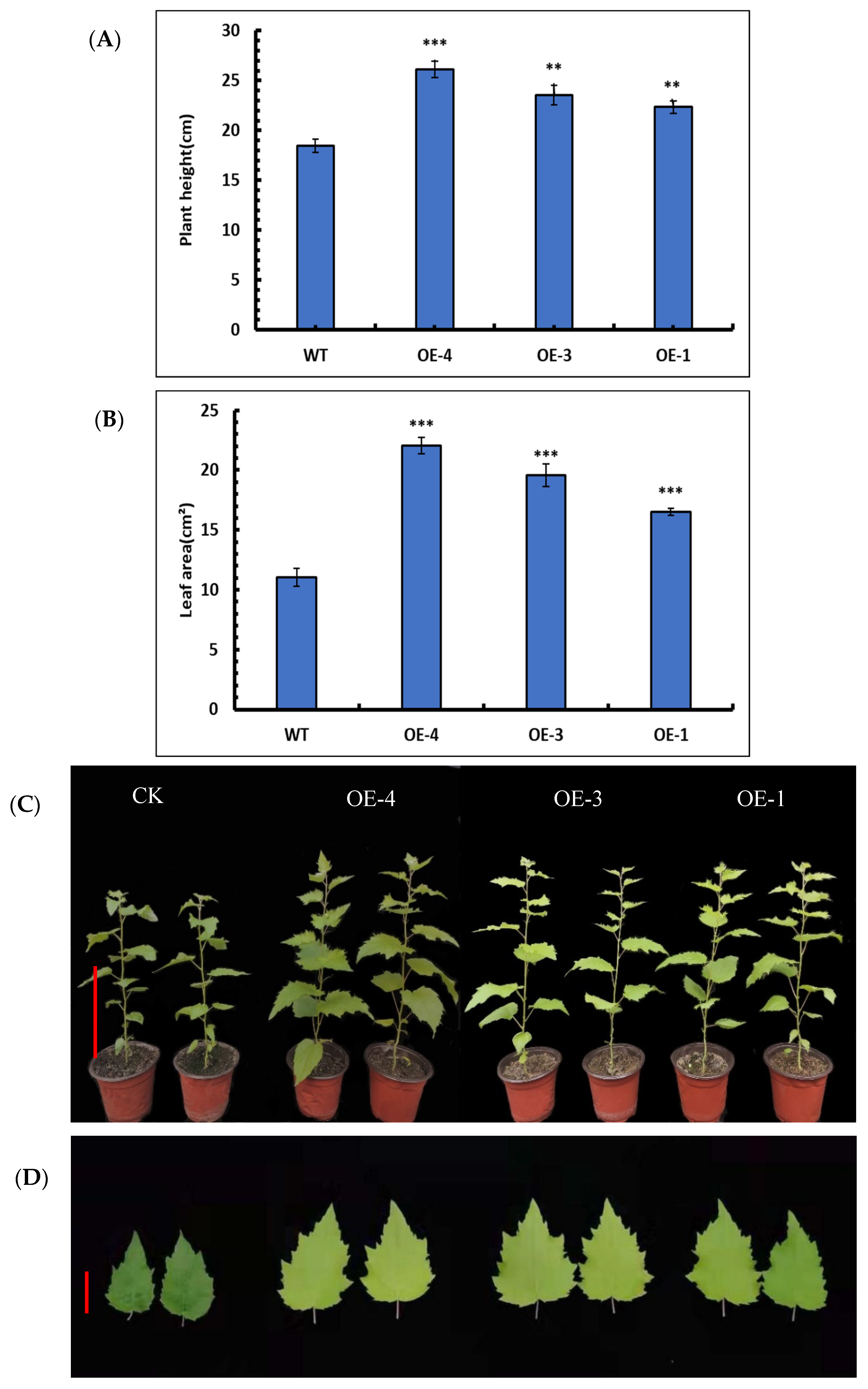
3.6.1. Changes in Plant Height in SpsNAC005 Overexpression Lines
3.6.2. Leaf Area Changes in SpsNAC005 Overexpression Lines
3.7. Morphological Changes in Transgenic P. hopeiensis under Stress Treatment
3.8. The Fresh Weight and Dry Weight of P. hopeiensis under Stress Treatment
3.9. Response of Antioxidant System and Osmotic Regulation of Transgenic P. hopeiensis to Stress
3.10. Up-Regulated Expression of SOS1, MPK6, HKT1, P5CS1, and PRODH1 Genes in Transgenic P. hopeiensis under Stress
4. Discussion
4.1. SpsNAC005 Gene Promotes the Growth and Development of Plants
4.2. SpsNAC005 Gene Enhances Stress Tolerance and Antioxidant Capacity of Transgenic P. hopeiensis
4.3. SpsNAC005 Gene Regulates Tolerance to Stress through Na+ Transporters Pathway
4.4. SpsNAC005 Gene Responds to Stress by Regulating Proline Synthesis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| qRT-PCR | Quantitative real-time PCR |
| cDNA | Complementary DNA |
References
- Christianson, J.A.; Dennis, E.S.; Llewellyn, D.J.; Wilson, I.W. ATAF NAC transcription factors: Regulators of plant stress signaling. Plant. Signal. Behav. 2010, 5, 428–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souer, E.; van Houwelingen, A.; Kloos, D.; Mol, J.; Koes, R. The No Apical Meristem Gene of Petunia Is Required for Pattern Formation in Embryos and Flowers and Is Expressed at Meristem and Primordia Boundaries. Cell 1996, 85, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Aida, M.; Ishida, T.; Fukaki, H.; Fujisawa, H.; Tasaka, M. Genes involved in organ separation in Arabidopsis: An analysis of the cup-shaped cotyledon mutant. Plant. Cell 1997, 9, 841–857. [Google Scholar] [CrossRef] [Green Version]
- Ernst, H.A.; Olsen, A.N.; Skriver, K.; Larsen, S.; Leggio, L.L. Structure of the conserved domain of ANAC, a member of the NAC family of transcription factors. EMBO Rep. 2004, 5, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, Q.; Xiong, L.; Lou, Z. A structural view of the conserved domain of rice stress-responsive NAC1. Protein Cell 2011, 2, 55–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, A.N.; Ernst, H.A.; Leggio, L.L.; Skriver, K. NAC transcription factors: Structurally distinct, functionally diverse. Trends Plant. Sci. 2005, 10, 79–87. [Google Scholar] [CrossRef]
- Li, W.; Han, L.; Qian, Y.Q.; Sun, Z.Y. Characteristics and Functions of NAC Transcription Factors in Plants. Chin. J. Appl. Environ. Biol. 2011, 17, 596–606. Available online: https://en.cnki.com.cn/Article_en/CJFDTotal-YYHS201104030.htm (accessed on 13 December 2021).
- Ooka, H.; Satoh, K.; Doi, K.; Nagata, T.; Otomo, Y.; Murakami, K.; Matsubara, K.; Osato, N.; Kawai, J.; Carninci, P.; et al. Comprehensive Analysis of NAC Family Genes in Oryza sativa and Arabidopsis thaliana. DNA Res. 2003, 10, 239–247. [Google Scholar] [CrossRef]
- Hu, R.; Qi, G.; Kong, Y.; Kong, D.; Gao, Q.; Zhou, G. Comprehensive Analysis of NAC Domain Transcription Factor Gene Family in Populus trichocarpa. BMC Plant. Biol. 2010, 10, 145. [Google Scholar] [CrossRef] [Green Version]
- Gong, L.; Zhang, H.; Liu, X.; Gan, X.; Nie, F.; Yang, W.; Zhang, L.; Chen, Y.; Song, Y.; Zhang, H. Ectopic expression of HaNAC1, an ATAF transcription factor from Haloxylon ammodendron, improves growth and drought tolerance in transgenic Arabidopsis. Plant. Physiol. Biochem. 2020, 151, 535–544. [Google Scholar] [CrossRef]
- Mahmood, K.; El-Kereamy, A.; Kim, S.-H.; Nambara, E.; Rothstein, S.J. ANAC032 Positively Regulates Age-Dependent and Stress-Induced Senescence in Arabidopsis thaliana. Plant. Cell Physiol. 2016, 57, 2029–2046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmood, K.; Xu, Z.; El-Kereamy, A.; Casaretto, J.A.; Rothstein, S.J. The Arabidopsis Transcription Factor ANAC032 Represses Anthocyanin Biosynthesis in Response to High Sucrose and Oxidative and Abiotic Stresses. Front. Plant Sci. 2016, 7, 1548. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Fan, C.H. Cloning and Expression Analysis of CaNAC23 Gene in Capsicum annuum L. Mol. Plant Breed. 2020, 18, 3854–3861. [Google Scholar]
- Zhang, L.Q.; Jia, X.H.; Zhao, J.W.; Liu, Y.X.; Hasiagula. Cloning and Expression Analysis of MfNAC37 from Medicago falcata under Salt Stress. Chin. J. Grassl. 2019, 41, 10–14+166. [Google Scholar] [CrossRef]
- Jia, D.; Jiang, Q.; van Nocker, S.; Gong, X.; Ma, F. An apple (Malus domestica) NAC transcription factor enhances drought tolerance in transgenic apple plants. Plant Physiol. Biochem. 2019, 139, 504–512. [Google Scholar] [CrossRef]
- Hu, H.; Dai, M.; Yao, J.; Xiao, B.; Li, X.; Zhang, Q.; Xiong, L. Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proc. Natl. Acad. Sci. USA 2006, 103, 12987–12992. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Zhang, J.; Li, J.; Sun, P.; Zhang, Y.; Xin, X.; Lu, M.; Hu, J. Genome-wide transcriptomic analysis of a desert willow, Salix psammophila, reveals the function of hub genes SpMDP1 and SpWRKY33 in drought tolerance. BMC Plant Biol. 2019, 19, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Li, W.X.; Liu, Z.X.; Yan, W.; Zhang, X.J.; Miu, S.G.; Huo, J.F.; Wang, Y.S. Study on Good Family Breeding of Salix Psammophila. J. Desert Res. 2008, 28, 679–684. [Google Scholar]
- Gu, M.H.; Xie, Z.H. Soil moisture characteristics of eight types of shelter forest in Kubuqi desert. Pratacult. Sci. 2017, 34, 2437–2444. [Google Scholar]
- Luo, W.X.; Liu, G.Q.; Li, J.J. Cultivation Techniques for Main Tree Species in Northwest China. China For. Press. 2007, 8, 682–687. [Google Scholar]
- Yu, X.Y. Cloning of NAC in Salix psammophila, Bioinformatics Analysis and Construction of Expression. Master’s Thesis, Inner Mongolia agricultural University, Hohhot, China, 2020. [Google Scholar]
- Horsch, R.B.; Fry, J.E.; Hoffmann, N.L.; Wallroth, M.; Eichholtz, D.; Rogers, S.G.; Fraley, R.T. A Simple and General Method for Transferring Genes into Plants. Science 1985, 227, 1229–1231. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.-S.; Li, F.; Xu, C.-J.; Zhang, S.-L.; Fu, C.-X. An efficient macro-method of genomic DNA isolation from Actinidia chinensis leaves. Hereditas 2004, 26, 529–531. [Google Scholar] [PubMed]
- Yang, H.F.; Bo, G.F. Cloning and expression analysis of SpsNAC042 gene in Salix psammophila. J. Northwest For. Univ. 2021, 36, 11–17+27. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Cao, Z.Z. Digital camera and Auto CAD based method for measuring the leaf area of landscape plants. Grassl. Turf. 2008, 3, 25–28. [Google Scholar]
- Cai, X.; Jiang, Z.; Tang, L.; Zhang, S.; Li, X.; Wang, H.; Liu, C.; Chi, J.; Zhang, X.; Zhang, J. Genome-wide characterization of carotenoid oxygenase gene family in three cotton species and functional identification of GaNCED3 in drought and salt stress. J. Appl. Genet. 2021, 62, 527–543. [Google Scholar] [CrossRef]
- Ning, H.L. Field Experiment and Statistical Methods; Science Press: Beijing, China, 2012; Available online: http://ss.zhizhen.com/detail_38502727e7500f26a81075d227d4798ebf0734766bdfb21d1921b0a3ea25510134114c969f2eae5c34597dee38cfeccc2f9303951adbee66eb7ea3d7539ba9795c5481485155be4acdc25c707daf213c? (accessed on 4 March 2022).
- Puranik, S.; Sahu, P.P.; Srivastava, P.S.; Prasad, M. NAC proteins: Regulation and role in stress tolerance. Trends Plant Sci. 2012, 17, 369–381. [Google Scholar] [CrossRef]
- Shen, H.; Yin, Y.; Chen, F.; Xu, Y.; Dixon, R.A. A Bioinformatic Analysis of NAC Genes for Plant Cell Wall Development in Relation to Lignocellulosic Bioenergy Production. Bioenerg. Res. 2009, 2, 217–232. [Google Scholar] [CrossRef]
- Hibara, K.; Takada, S.; Tasaka, M. CUC1 gene activates the expression of SAM-related genes to induce adventitious shoot formation. Plant J. 2003, 36, 687–696. [Google Scholar] [CrossRef]
- Zhong, R.; Richardson, E.A.; Ye, Z.-H. Two NAC domain transcription factors, SND1 and NST1, function redundantly in regulation of secondary wall synthesis in fibers of Arabidopsis. Planta 2007, 225, 1603–1611. [Google Scholar] [CrossRef]
- Mitsuda, N.; Iwase, A.; Yamamoto, H.; Yoshida, M.; Seki, M.; Shinozaki, K.; Ohme-Takagi, M. NAC Transcription Factors, NST1 and NST3, Are Key Regulators of the Formation of Secondary Walls in Woody Tissues of Arabidopsis. Plant Cell 2007, 19, 270–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Avci, U.; Grant, E.H.; Haigler, C.H.; Beers, E.P. XND1, a member of the NAC domain family in Arabidopsis thaliana, negatively regulates lignocellulose synthesis and programmed cell death in xylem. Plant J. 2008, 53, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Tran, L.-S.P.; Nakashima, K.; Sakuma, Y.; Simpson, S.D.; Fujita, Y.; Maruyama, K.; Fujita, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Isolation and Functional Analysis of Arabidopsis Stress-Inducible NAC Transcription Factors That Bind to a Drought-Responsive cis-Element in the early responsive to dehydration stress 1 Promoter[W]. Plant Cell 2004, 16, 2481–2498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Gan, S. AtNAP, a NAC family transcription factor, has an important role in leaf senescence. Plant J. 2006, 46, 601–612. [Google Scholar] [CrossRef]
- Yang, Z.-T.; Lu, S.-J.; Wang, M.-J.; Bi, D.-L.; Sun, L.; Zhou, S.-F.; Song, Z.-T.; Liu, J.-X. A plasma membrane-tethered transcription factor, NAC062/ANAC062/NTL6, mediates the unfolded protein response in Arabidopsis. Plant J. 2014, 79, 1033–1043. [Google Scholar] [CrossRef]
- Jung, J.-H.; Park, C.-M. Auxin modulation of salt stress signaling in Arabidopsis seed germination. Plant Signal. Behav. 2011, 6, 1198–1200. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Sun, J.; Wu, Y. Arabidopsis ATAF1 enhances the tolerance to salt stress and ABA in transgenic rice. J. Plant Res. 2016, 129, 955–962. [Google Scholar] [CrossRef]
- Yang, H.F.; Yu, X.W.; Jin, N.; He, Y.J.; Li, A.Y.; Zhang, X.; Wang, Y.Z.; Gao, Y.Y.; Wang, L. Cloning and bioinformatics analysis of SpsNAC005 gene in Salix psammophila. J. Inn. Mong. Agric Univ. 2021, 42, 65–71. [Google Scholar]
- Sun, L.; Zhang, P.; Wang, R.; Wan, J.; Ju, Q.; Rothstein, S.J.; Xu, J. The SNAC-A Transcription Factor ANAC032 Reprograms Metabolism in Arabidopsis. Plant Cell Physiol. 2019, 60, 999–1010. [Google Scholar] [CrossRef]
- Wang, S. Cloning and Expression Analysis of Solanum Lycopersicum NAC Transcription Factors. Master’s Thesis, Tianjin University, Tianjin, China, 2014. [Google Scholar]
- Shao, H.; Wang, H.; Tang, X. NAC transcription factors in plant multiple abiotic stress responses: Progress and prospects. Front. Plant Sci. 2015, 6, 902. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.S.; Kim, Y.S.; Baek, K.H.; Jung, H.; Ha, S.-H.; Do Choi, Y.; Kim, M.; Reuzeau, C.; Kim, J.-K. Root-Specific Expression of OsNAC10 Improves Drought Tolerance and Grain Yield in Rice under Field Drought Conditions. Plant Physiol. 2010, 153, 185–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.; Huang, A.; Li, J.; Gao, L.; Feng, Y.; Pemberton, E.; Chen, C. OsNAC45 plays complex roles by mediating POD activity and the expression of development-related genes under various abiotic stresses in rice root. Plant Growth Regul. 2018, 84, 519–531. [Google Scholar] [CrossRef]
- Rong, Y.P.; Tang, B.; Li, P.; Zhang, J.Q.; Chen, Q.F.; Zhu, L.W.; Deng, J.; Huang, J. Identification and Expression of NAC Transcription Factor FtNAC17 in Tartary Buckwheat. Biotechnol. Bull. 2021, 37, 174. [Google Scholar] [CrossRef]
- Zhang, L.; Xiong, H.H.; Cao, Q.; Zhao, J.L.; Zhang, H.G. Drought Resistance of Larch NAC Gene by Transient Genetic Transformation. Bull. Bot. Res. 2020, 40, 394–400. [Google Scholar]
- Zhou, L.D.; Yao, Z.P. Cloning and characterization of the HaNAC20 gene of P. sylvestris. Acta Agric. Boreali-Occident. Sin. 2021, 30, 1556–1564. [Google Scholar]
- Wang, X.Y.; Zhang, X.M. Function analysis of poplar transcription factor PsnNAC030 gene. J. Northeast. For. Univ. 2021, 49, 1–8. [Google Scholar]
- Yao, W.; Zhou, B.; Zhang, X.; Zhao, K.; Cheng, Z.; Jiang, T. Transcriptome analysis of transcription factor genes under multiple abiotic stresses in Populus simonii × P.nigra. Gene 2019, 707, 189–197. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, N.K. Cloning and functional analysis of drought-related gene IbNAC72 in sweet potato. Acta Agron. Sin. 2020, 46, 1649–1658. [Google Scholar]
- Liu, W.P. Mining of Drought-Resistant Related Genes in Maize and Functional Identification of Drought-Resistant Genes. Ph.D. Thesis, Northeastern Agricultural University, Harbin, China, 2020. [Google Scholar]
- Wang, R.; Zhang, Y.; Wang, C.; Wang, Y.-C.; Wang, L.-Q. ThNAC12 from Tamarix hispida directly regulates ThPIP2;5 to enhance salt tolerance by modulating reactive oxygen species. Plant Physiol. Biochem. 2021, 163, 27–35. [Google Scholar] [CrossRef]
- Foster, K.J.; Miklavcic, S.J. A Comprehensive Biophysical Model of Ion and Water Transport in Plant Roots. II. Clarifying the Roles of SOS1 in the Salt-Stress Response in Arabidopsis. Front. Plant Sci. 2019, 10, 1121. [Google Scholar] [CrossRef]
- Shi, H.; Lee, B.; Wu, S.-J.; Zhu, J.-K. Overexpression of a plasma membrane Na+/H+ antiporter gene improves salt tolerance in Arabidopsis thaliana. Nat. Biotechnol. 2003, 21, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Sun, J.; Cao, P.; Ren, L.; Liu, C.; Chen, S.; Chen, F.; Jiang, J. Variation in tissue Na+ content and the activity of SOS1 genes among two species and two related genera of Chrysanthemum. BMC Plant Biol. 2016, 16, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, N.S.; Tuteja, R.; Tuteja, N. Signaling through MAP kinase networks in plants. Arch. Biochem. Biophys. 2006, 452, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, Y. Activation of Salicylic Acid–Induced Protein Kinase, a Mitogen-Activated Protein Kinase, Induces Multiple Defense Responses in Tobacco. Plant Cell 2001, 13, 1877–1889. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Nie, J.; Cao, C.; Jin, Y.; Yan, M.; Wang, F.; Liu, J.; Xiao, Y.; Liang, Y.; Zhang, W. Phosphatidic acid mediates salt stress response by regulation of MPK6 in Arabidopsis thaliana. New Phytol. 2010, 188, 762–773. [Google Scholar] [CrossRef]
- Rubio, F.; Gassmann, W.; Schroeder, J.I. Sodium-Driven Potassium Uptake by the Plant Potassium Transporter HKT1 and Mutations Conferring Salt Tolerance. Science 1995, 270, 1660–1663. [Google Scholar] [CrossRef]
- Waters, S.; Gilliham, M.; Hrmova, M. Plant High-Affinity Potassium (HKT) Transporters Involved in Salinity Tolerance: Structural Insights to Probe Differences in Ion Selectivity. Int. J. Mol. Sci. 2013, 14, 7660–7680. [Google Scholar] [CrossRef] [Green Version]
- Van Bezouw, R.F.H.M.; Janssen, E.M.; Ashrafuzzaman, M.; Ghahramanzadeh, R.; Kilian, B.; Graner, A.; Visser, R.G.F.; van der Linden, C.G. Shoot sodium exclusion in salt stressed barley (Hordeum vulgare L.) is determined by allele specific increased expression of HKT1;5. J. Plant Physiol. 2019, 241, 153029. [Google Scholar] [CrossRef]
- Suzuki, K.; Yamaji, N.; Costa, A.; Okuma, E.; Kobayashi, N.I.; Kashiwagi, T.; Katsuhara, M.; Wang, C.; Tanoi, K.; Murata, Y.; et al. OsHKT1;4-mediated Na+ transport in stems contributes to Na+ exclusion from leaf blades of rice at the reproductive growth stage upon salt stress. BMC Plant Biol. 2016, 16, 22. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Gao, Z. Functional Identification of Grape Potassium Ion Transporter VviHKT1;7 Under Salt Stress. Sci. Agric. Sinina 2021, 54, 1952–1963. [Google Scholar]
- Delauney, A.; Hu, C.; Kishor, P.; Verma, D. Cloning of ornithine delta-aminotransferase cDNA from Vigna aconitifolia by trans-complementation in Escherichia coli and regulation of proline biosynthesis. J. Biol. Chem. 1993, 268, 18673–18678. [Google Scholar] [CrossRef]
- Kishor, P.B.K.; Sangam, S.; Amrutha, R.N.; Laxmi, P.S.; Naidu, K.R.; Rao, K.R.S.S.; Rao, S.; Reddy, K.J.; Theriappan, P.; Sreenivasulu, N. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: Its implications in plant growth and abiotic stress tolerance. Curr. Sci. 2005, 88, 424–438. [Google Scholar]
- Funck, D.; Eckard, S.; Müller, G. Non-redundant functions of two proline dehydrogenase isoforms in Arabidopsis. BMC Plant Biol. 2010, 10, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanjo, T.; Kobayashi, M.; Yoshiba, Y.; Kakubari, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Antisense suppression of proline degradation improves tolerance to freezing and salinity in Arabidopsis thaliana. FEBS Lett. 1999, 461, 205–210. [Google Scholar] [CrossRef] [Green Version]
- Yoshiba, Y.; Kiyosue, T.; Katagiri, T.; Ueda, H.; Mizoguchi, T.; Yamaguchi-Shinozaki, K.; Wada, K.; Harada, Y.; Shinozaki, K. Correlation between the induction of a gene for Δ1-pyrroline-5-carboxylate synthetase and the accumulation of proline in Arabidopsis thaliana under osmotic stress. Plant J. 1995, 7, 751–760. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene-ID | Amino Acids | Molecular Weight (kD) | Theoretical pI | Instability Index | Aliphatic Index | Hydrophobicity | Subcellular Localization |
|---|---|---|---|---|---|---|---|
| SpsNAC005 | 308 | 35.24 | 6.02 | 43.68 | 60.91 | Hydrophilic | Nucleolus, Nucleus, Nucleoplasm |
| SpsNAC034 | 598 | 65.82 | 5.32 | 68.83 | 71.92 | Hydrophilic | Nucleolus, Nucleus, Nucleoplasm |
| SpsNAC041 | 294 | 33.09 | 6.46 | 46.12 | 65.34 | Hydrophilic | Cytoplasm, Cytosol |
| SpsNAC042 | 299 | 34.19 | 6.45 | 51.39 | 68.09 | Hydrophilic | Cytoplasm, Cytosol |
| SpsNAC120 | 343 | 38.30 | 8.56 | 40.74 | 67.53 | Hydrophilic | Nucleolus, Nucleus, Nucleoplasm |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Fan, L.; Yu, X.; Zhang, X.; Hao, P.; Wei, D.; Zhang, G. Analysis of the NAC Gene Family in Salix and the Identification of SpsNAC005 Gene Contributing to Salt and Drought Tolerance. Forests 2022, 13, 971. https://doi.org/10.3390/f13070971
Yang H, Fan L, Yu X, Zhang X, Hao P, Wei D, Zhang G. Analysis of the NAC Gene Family in Salix and the Identification of SpsNAC005 Gene Contributing to Salt and Drought Tolerance. Forests. 2022; 13(7):971. https://doi.org/10.3390/f13070971
Chicago/Turabian StyleYang, Haifeng, Lijiao Fan, Xingwang Yu, Xinqian Zhang, Pu Hao, Dongshan Wei, and Guosheng Zhang. 2022. "Analysis of the NAC Gene Family in Salix and the Identification of SpsNAC005 Gene Contributing to Salt and Drought Tolerance" Forests 13, no. 7: 971. https://doi.org/10.3390/f13070971
APA StyleYang, H., Fan, L., Yu, X., Zhang, X., Hao, P., Wei, D., & Zhang, G. (2022). Analysis of the NAC Gene Family in Salix and the Identification of SpsNAC005 Gene Contributing to Salt and Drought Tolerance. Forests, 13(7), 971. https://doi.org/10.3390/f13070971