
AFLP-Based Genetic Structure of Lithuanian Populations of Small Balsam (Impatiens parviflora DC.) in Relation to Habitat Characteristics

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
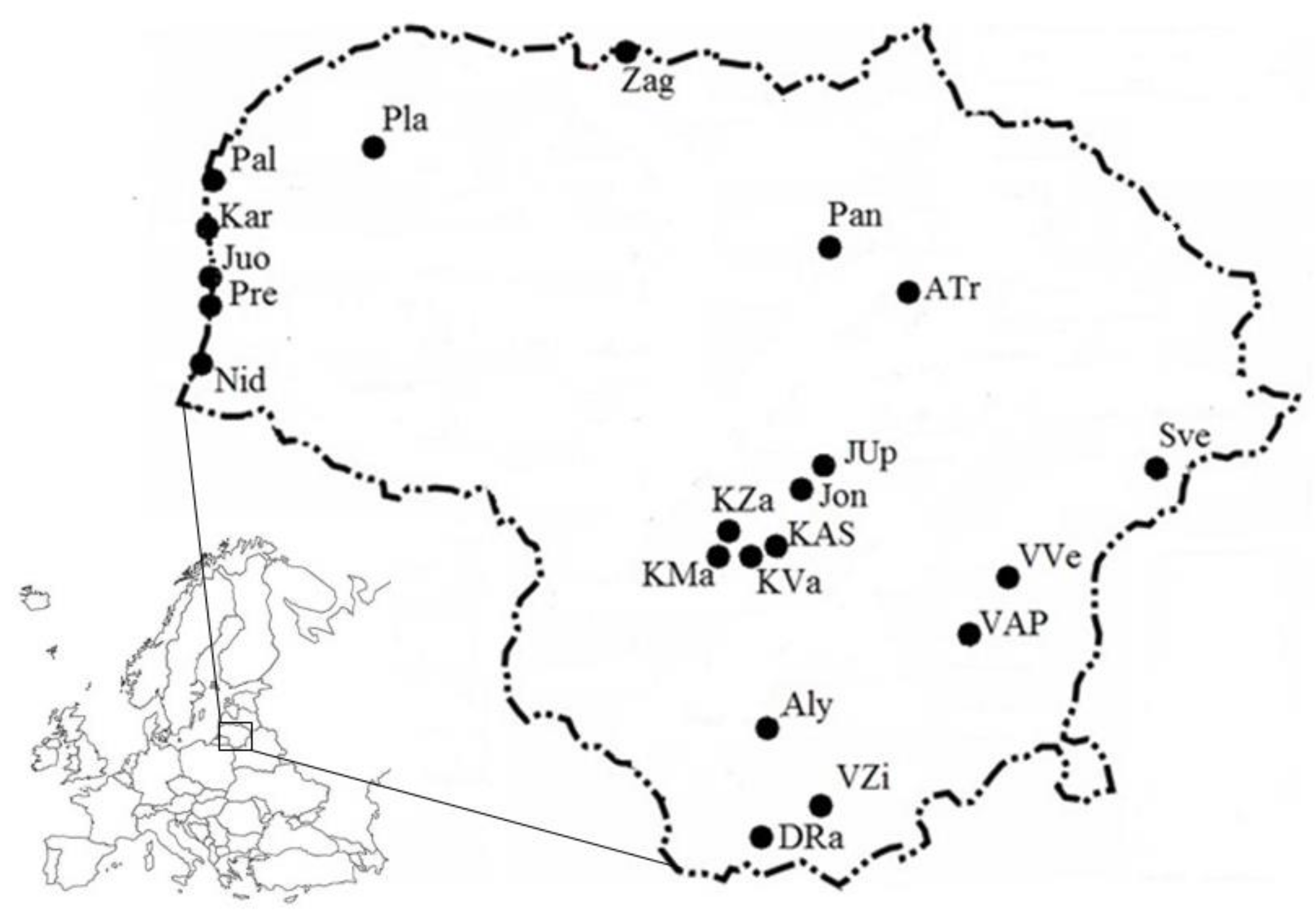
2.1. Study Area
2.2. Molecular Analysis
2.3. Abiotic and Biotic Features of the Sites
2.4. Data Analysis
3. Results
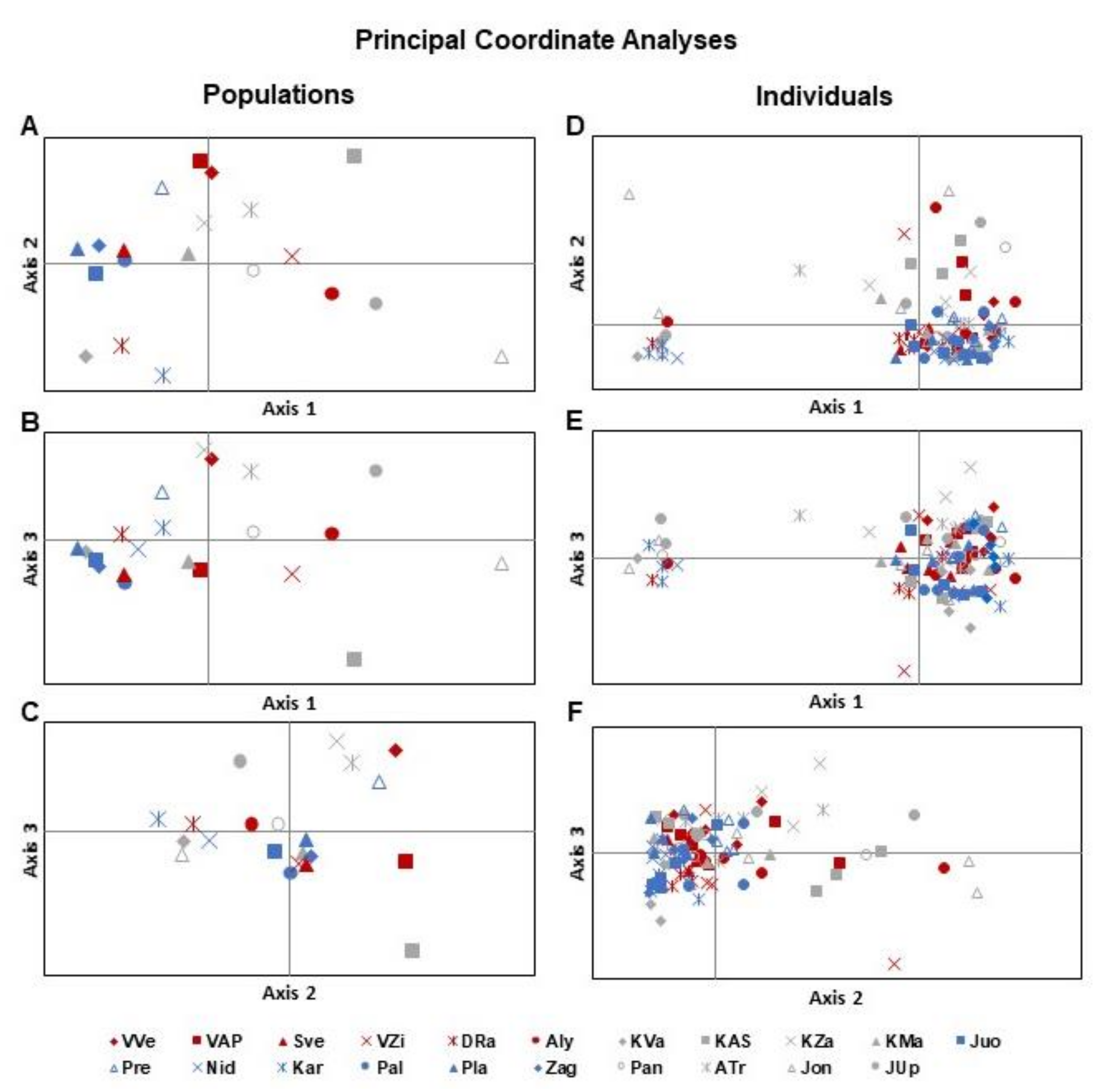
3.1. Analysis of Genetic Diversity
3.2. Herbaceous Species Composition
3.3. Relations between Molecular and Environmental Variables
4. Discussion
4.1. Molecular Features of Impatiens parviflora
4.2. Peculiarities of Herbaceous Plant Assemblages
4.3. Importance of Genetical and Ecological Variables at Sites with Impatiens parviflora
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegert, M.J. Environmental Sciences in the Twenty-First Century. Front. Environ. Sci. 2016, 4, 16. [Google Scholar] [CrossRef]
- Langmaier, M.; Lapin, K. A Systematic review of the impact of invasive alien plants on forest regeneration in European temperate forests. Front. Plant Sci. 2020, 11, 524969. [Google Scholar] [CrossRef]
- Xie, P.; Liu, T.; Chen, H.; Su, Z. Community structure and soil mineral concentration in relation to plant invasion in a subtropical urban and rural ecotone. Forests 2021, 12, 185. [Google Scholar] [CrossRef]
- Wagner, V.; Chytrý, M.; Jiménez-Alfaro, B.; Pergl, J.; Hennekens, S.; Biurrun, I.; Knollová, I.; Berg, C.; Vassilev, K.; Rodwell, J.S.; et al. Alien plant invasions in European woodlands. Divers. Distrib. 2017, 23, 969–981. [Google Scholar] [CrossRef]
- Biodiversity Strategy for 2030. European Commission, Brussels. 2020. Available online: https://ec.europa.eu/environment/strategy/biodiversity-strategy-2030_en (accessed on 31 January 2022).
- New EU Forest Strategy for 2030. European Commission, Brussels. 2021. Available online: https://ec.europa.eu/environment/strategy/forest-strategy_en (accessed on 31 January 2022).
- The Ministry of Environment of the Republic of Lithuania. List of Invasive Species in Lithuania, Approved by the Minister of Environment of the Republic of Lithuania, Order No. D1-433 of 27 August 2004. Register Of Legal Acts, 20 July 2015, No. 11487. 2015. Available online: https://www.e-tar.lt/portal/lt/legalAct/TAR.7B6390A69C91/asr (accessed on 28 December 2021).
- Marozas, V.; Cekstere, G.; Laivins, M.; Straigyte, L. Comparison of neophyte communities of Robinia pseudoacacia L. and Acer negundo L. in the eastern Baltic Sea region cities of Riga and Kaunas. Urban For. Urban Green. 2015, 14, 826–834. [Google Scholar] [CrossRef]
- Straigyte, L.; Cekstere, G.; Laivins, M.; Marozas, V. The spread, intensity and invasiveness of the Acer negundo in Riga and Kaunas. Dendrobiology 2015, 74, 157–168. [Google Scholar] [CrossRef]
- Krevš, A.; Darginavičienė, J.; Gylytė, B.; Grigutytė, R.; Jurkonienė, S.; Karitonas, R.; Kučinskienė, A.; Pakalnis, R.; Sadauskas, K.; Vitkus, R.; et al. Ecotoxicological effects evoked in hydrophytes by leachates of invasive Acer negundo and autochthonous Alnus glutinosa fallen off leaves during their microbial decomposition. Environ. Pollut. 2013, 173, 75–84. [Google Scholar] [CrossRef]
- Manusadžianas, L.; Darginavičienė, J.; Gylytė, B.; Jurkonienė, S.; Krevš, A.; Kučinskienė, A.; Mačkinaitė, V.; Pakalnis, R.; Sadauskas, K.; Sendžikaitė, J.; et al. Ecotoxicity effects triggered in aquatic organisms by invasive Acer negundo and native Alnus glutinosa leaf leachates obtained in the process of aerobic decomposition. Sci. Total Environ. 2014, 496, 35–44. [Google Scholar] [CrossRef]
- Krevš, A.; Kučinskienė, A.; Mačkinaitė, R.; Manusadžianas, L. Microbial colonization and decomposition of invasive and native leaf litter in the littoral zone of lakes of different trophic state. Limnologica 2017, 67, 54–63. [Google Scholar] [CrossRef]
- Ellenberg, H.; Weber, H.E.; Wirth, V.; Werner, W.; Paulien, D. Zeigerwerte von Pflanzen in Mitteleuropa. Scr. Geobot. 1992, 18, 1–258. [Google Scholar]
- Tanner, R. Datasheets: Impatiens parviflora (small balsam). CAB Europe—UK, Bakeham Lane, Egham, Surrey TW20 9TY, UK. 2008. Available online: http://www.cabi.org/isc/datasheet/28768 (accessed on 28 January 2022).
- Jarčuška, B.; Slezák, M.; Hrivnák, R.; Senko, D. Invasibility of alien Impatiens parviflora in temperate forest understories. Flora 2016, 224, 14–23. [Google Scholar] [CrossRef]
- Reczyńska, K.; Świerkosz, K.; Dajdok, Z. The spread of Impatiens parviflora DC. in Central European oak forests—Another stage of invasion? Acta Soc. Bot. Pol. Pol. 2015, 84, 401–411. [Google Scholar] [CrossRef]
- Maslo, S.; Šarić, Š. Small Balsam, Impatiens parviflora (Balsaminaceae): A new alien species to the flora of Bosnia and Herzegovina. Phytol. Balc. 2019, 25, 69–73. [Google Scholar]
- Coombe, D.E. Biological flora of the British Isles, Impatiens parviflora DC. J. Ecol. 1956, 44, 701–713. [Google Scholar] [CrossRef]
- Elias, P. Biological and ecological causes of invasion of Impatiens parviflora DC. into forest communities in Central Europe. AHR 1999, 1, 1–3. [Google Scholar]
- Trepl, L. Uber Impatiens parviflora DC. als Agriophyt in Mitteleuropa. In Dissertationes Botaniceae; Gantner, A.R., Ed.; Kommanditgesellschaft: Vaduz, Germany, 1984. (In German) [Google Scholar]
- Hejda, M. What is the impact of Impatiens parviflora on diversity and composition of herbal layer communities of temperate forests? PLoS ONE 2012, 7, e39571. [Google Scholar] [CrossRef]
- Chmura, D. Biology and Ecology of an Invasion of Impatiens parviflora DC. In Natural and Semi-Natural Habitats; Akademia Techniczno-Humanistyczna w Bielsku-Bialej: Bialsko-Biela, Poland, 2014; 216p. [Google Scholar]
- Kamieński, F. Nowy nabytek flory krajowej. Wszechświat 1884, 3, 517–518. (In Polish) [Google Scholar]
- Priede, A. Invasive Neophytes in the Flora of Latvia: Distribution and Dynamics. Ph.D. Thesis, University of Latvia, Riga, Latvia, 2009; p. 26. [Google Scholar]
- Matulionis, P. Latin-Lithuanian-Russian-Polish Dictionary of Lithuania Plants; M. Kuktos Print: Vilnius, Lithuania, 1907; 112p. [Google Scholar]
- Wisnewski, P. Nienotowane stanowiska kilku rzadszych krajowych i zawleczonych roslin w Wilensczyznie. Acta Soc. Bot. Pol. 1934, 11, 207–209. (In Polish) [Google Scholar]
- Dagys, J.; Kuprevičius, J.; Minkevičius, A. Impatiens parviflora. In Manual for Indentification of Lithuanian Plants (Vadovas Lietuvos augalams pažinti); Kuprevičius, J., Ed.; Spindulys: Kaunas, Lithuania, 1934; 358p. (In Lithuanian) [Google Scholar]
- Vailionis, L.; Dagys, J. Lithuanian Dictionary of Botany (Lietuviškas Botanikos Žodynas); Varpas: Kaunas, Lithuania, 1938; Volume 1, 598p. (in Lithuanian) [Google Scholar]
- Natkevicaite-Ivanauskiene, M. Adventive flora of Lithuanian Soviet Republic. Proc. Inst. Biol. 1951, 1, 77–124. (In Lithuanian) [Google Scholar]
- Snarskis, P. A Guide of Identification of Lithuania Plants; Mintis: Vilnius, Lithuania, 1954; 530p. (In Lithuanian) [Google Scholar]
- Obidziński, T.; Symonides, E. The influence of the groundlayer structure on the invasion of small balsam (Impatiens parviflora DC.) to natural and degraded forests. Acta Soc. Bot. Pol. Pol. 2000, 69, 311–318. [Google Scholar] [CrossRef]
- Klimko, M.; Piskorz, R. The usefulness of retrospective research along the road-forest border zone for reconstruction of invasion of oak-hombeam forest communities by Impatiens parviflora DC. Ecological Questions. Int. J. Controv. Probl. Ecol. 2003, 3, 63–69. [Google Scholar]
- Priede, A. Distribution of some invasive alien plant species in riparian habitats in Latvia. Bot. Lith. 2009, 14, 137–150. [Google Scholar]
- Dobravolskaitė, R. Alien species Impatiens parviflora invasion into forest communities of Lithuania. Bot. Lith. 2012, 18, 3–12. [Google Scholar] [CrossRef]
- Kupcinskiene, E.; Zybartaite, L.; Janulioniene, R.; Zukauskiene, J.; Paulauskas, A. Molecular diversity of small balsam populations in relation to site characteristics. Cent. Eur. J. Biol. 2013, 8, 1048–1061. [Google Scholar] [CrossRef]
- Marozas, V.; Preikša, Ž.; Koike, T.; Watanabe, T. Impact of nitrogen pollution disturbances on forest vegetation and fungi near a fertilizer factory. Eur. J. For. Res. 2022, 22, 52–58. [Google Scholar]
- Dormontt, E.E.; Lowe, A.J.; Prentis, P.J. Is rapid adaptive evolution important in successful invasions? In Fifty Years of Invasion Ecology: The Legacy of Charles Elton; Richardson, D.M., Ed.; Wiley-Blackwell: Oxford, UK, 2011; pp. 175–193. [Google Scholar]
- Krokaitė, E.; Janulionienė, R.; Jocienė, L.; Rekašius, T.; Rajackaitė, G.; Paulauskas, A.; Marozas, V.; Kupcinskiene, E. Relating invasibility and invasiveness: Case study of Impatiens parviflora. Front. Ecol. Evol. 2022, 10, 1–17. [Google Scholar] [CrossRef]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; Vandelee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M.; et al. AFLP—a new technique for DNA-fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef]
- Powell, W.; Machray, G.C.; Provan, J. Polymorphism revealed by simple sequence repeats. Trends Plant Sci. 1996, 1, 215–222. [Google Scholar] [CrossRef]
- Crawford, L.A.; Koscinski, D.; Keyghobadi, N. A call for more transparent reporting of error rates: The quality of AFLP data in ecological and evolutionary research. Mol. Ecol. 2012, 21, 5911–5917. [Google Scholar] [CrossRef]
- Bensch, S.; Akesson, M. Ten years of AFLP in ecology and evolution: Why so few animals? Mol. Ecol. 2005, 14, 2899–2914. [Google Scholar] [CrossRef] [PubMed]
- Poczai, P.; Varga, I.; Laos, M.; Cseh, A.; Bell, N.; Valkonen, J.; Hyvönen, J. Advances in plant gene-targeted and functional markers: A review. Plant Methods 2013, 9, 6. [Google Scholar] [CrossRef] [PubMed]
- Amsellem, L.; Noyer, J.L.; Le Bourgeois, T.; Hossaert-Mckey, M. Comparison of genetic diversity of the invasive weed Rubus alceifolius Poir. (Rosaceae) in its native range and in areas of introduction, using amplified fragment length polymorphism (AFLP) markers. Mol. Ecol. 2000, 9, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Jahodová, Š.; Trybush, S.; Pyšek, P.; Wade, M.; Karp, A. Invasive species of Heracleum in Europe: An insight into genetic relationships and invasion history. Divers. Distrib. 2007, 13, 99–114. [Google Scholar] [CrossRef]
- Chun, Y.J.; Nason, J.D.; Moloney, K.A. Comparison of quantitative and molecular genetic variation of native vs. invasive populations of purple loosestrife (Lythrum salicaria L., Lythraceae). Mol. Ecol. 2009, 18, 3020–3035. [Google Scholar] [CrossRef] [PubMed]
- Jocienė, L.; Krokaitė, E.; Shakeneva, D.; Rekašius, T.; Stanys, V.; Šikšnianienė, J.B.; Žvingila, D.; Paulauskas, A.; Kupčinskienė, E. Relationship between genetic and environmental characteristics of Lithuanian populations of purple loosestrife (Lythrum salicaria). J. Environ. Eng. Landsc. Manag. 2022, 30, 81–93. [Google Scholar] [CrossRef]
- Casler, M.D.; Phillips, M.M.; Krohn, A.L. DNA polymorphisms reveal geographic races of reed canarygrass. Crop Sci. 2009, 49, 2139–2148. [Google Scholar] [CrossRef]
- Wu, H.; Qiang, S.; Peng, G. Genetic diversity in Veronica hederifolia (Plantaginaceae), an invasive weed in China, assessed using AFLP markers. In Annales Botanici Fennici; Finnish Zoological and Botanical Publishing Board: Helsinki, Finland, 2010; Volume 47, pp. 190–198. [Google Scholar]
- Bzdęga, K.; Janiak, A.; Tarłowska, S.; Kurowska, M.; Tokarska-Guzik, B.; Szarejko, I. Unexpected genetic diversity of Fallopia japonica from Central Europe revealed after AFLP analysis. Flora: Morphol. Distrib. Funct. Ecol. Plants 2012, 207, 636–645. [Google Scholar] [CrossRef]
- Wang, T.; Chen, G.; Zan, Q.; Wang, C.; Su, Y.J. AFLP genome scan to detect genetic structure and candidate loci under selection for local adaptation of the invasive weed Mikania micrantha. PLoS ONE 2012, 7, e41310. [Google Scholar] [CrossRef]
- Zhao, S.Y.; Sun, S.G.; Dai, C.; Gituru, R.W.; Chen, J.M.; Wang, Q.F. Genetic variation and structure in native and invasive Solidago canadensis populations. Weed Res. 2015, 55, 163–172. [Google Scholar] [CrossRef]
- Carr, J.; Korban, S.S. Evaluating genetic relationships in seed impatiens, Impatiens walleriana, using AFLP profiling. Plant Breed. 2004, 123, 577–581. [Google Scholar] [CrossRef]
- Carr, J.; Xu, M.; Dudley, J.W.; Korban, S.S. AFLP analysis of genetic variability in New Guinea impatiens. Theor. Appl. Genet. 2003, 106, 1509–1516. [Google Scholar] [CrossRef] [PubMed]
- Hatcher, P.E.; Wilkinson, M.J.; Albani, M.C.; Hebbern, C.A. Conserving marginal populations of the food plant (Impatiens noli-tangere) of an endangered moth (Eustroma reticulatum) in a changing climate. Biol. Conserv. 2004, 116, 305–317. [Google Scholar] [CrossRef]
- Von Wettberg, E.J. Do founder effects impact invasions? Genetic variation in North American Impatiens naturalized in England. In Proceedings of the ESA/SER Joint Meeting, Invasion: Genetics and Evolutionary Processes, San Jose, CA, USA, 5–10 August 2007. COS 51-5. [Google Scholar]
- Von Wettberg, E.J.; Remington, D.L.; Schmitt, J. Partitioning adaptive differentiation across a patchy landscape: Shade avoidance traits in Impatiens capensis. Evolution 2008, 62, 654–667. [Google Scholar] [CrossRef]
- Komosińska, E.; Wòdkiewicz, M.; Jarzyna, I.; Jarochowska, E.; Milanowski, R.; Chwedorzewska, K.; Wyszomirski, T. Some attempts to detect genetic differences between populations of small balsam (Impatiens parviflora DC.). Biodivers. Res. Conserv. 2006, 3–4, 245–247. [Google Scholar]
- Chmura, D.; Sierka, E. Relationships between invasive plant Impatiens parviflora and species richness of forest floor vegetation. Pol. J. Ecol. 2006, 54, 417–428. [Google Scholar]
- Vervoort, A.; Jacquemart, A.L. Habitat overlap of the invasive Impatiens parviflora DC. with its native congener I. noli-tangere L. Phytocoenologia 2012, 42, 249–257. [Google Scholar] [CrossRef]
- Florianova, A.; Münzbergová, Z. Invasive Impatiens parviflora has negative impact on native vegetation in oak-hornbeam forests. Flora 2017, 226, 10–16. [Google Scholar] [CrossRef]
- Zybartaite, L.; Zukauskiene, J.; Jodinskiene, M.; Janssens, S.B.; Paulauskas, A.; Kupcinskiene, E. RAPD analysis of genetic diversity among Lithuanian populations of Impatiens glandulifera. Zemdirb.-Agric. 2011, 98, 391–398. [Google Scholar]
- Kloss, L.; Fischer, M.; Durka, W. Land-use effects on genetic structure of a common grassland herb: A matter of scale. Basic Appl. Ecol. 2011, 12, 440–448. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Pflanzensoziologie, Grundzüge der Vegetationskunde, 3rd ed.; Springer: Vienna, Austria, 1964; p. 613. [Google Scholar]
- Marozas, V. Effect of the coniferous forest-grassland edge on ground vegetation in the mixed European forest zone, Lithuania. Dendrobiology 2014, 71, 15–22. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software Structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- McCune, B.; Mefford, M.J. PC-ORD. Multivariate Analysis of Ecological Data, Version 6.0 MjM Software; MjM Software Design: Gleneden Beach, OR, USA, 2011; 28p. [Google Scholar]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Frankham, R. Resolving the genetic paradox in invasive species. Heredity 2005, 94, 385. [Google Scholar] [CrossRef] [PubMed]
- Cross, H.B.; Lowe, A.J.; Gurgel, C.F.D. DNA barcoding of invasive species. In Fifty Years of Invasion Ecology: The Legacy of Charles Elton; Richardson, D.M., Ed.; Wiley-Blackwell: Oxford, UK, 2011; pp. 289–300. [Google Scholar]
- Hagenblad, J.; Hülskötter, J.; Acharya, K.P.; Brunet, J.; Chabrerie, O.; Cousins, S.A.O.; Dar, P.A.; Diekmann, M.; De Frenne, P.; Hermy, M.; et al. Low genetic diversity despite multiple introductions of the invasive plant species Impatiens glandulifera in Europe. BMC Genet. 2015, 16, 103. [Google Scholar] [CrossRef] [PubMed]
- Korpelainen, H.; Elshibli, S. Assessment of genetic relationships among native and introduced Himalayan balsam (Impatiens glandulifera) plants based on genome profiling. Ecol. Evol. 2021, 11, 13295–13304. [Google Scholar] [CrossRef]
- Jocienė, L. Genetic Diversity of Invasive Populations of Himalayan Balsam (Impatiens glandulifera Royle). Ph.D. Thesis, Vytautas Magnus University, Kaunas, Lithuania, 2015. [Google Scholar]
- Jakše, J.; Kindlhofer, K.; Javornik, B. Assessment of genetic variation and differentiation of hop genotypes by microsatellite and AFLP markers. Genome 2001, 44, 773–782. [Google Scholar] [CrossRef]
- Bandelj, D.; Jakse, J.; Javornik, B. Amplification of fluorescent-labelled microsatellite markers in olives by a novel, economic method. Acta Agric. Slov. 2004, 83, 323–329. [Google Scholar]
- Li, L.; Wanapu, C.; Huang, X.; Huang, T.; Li, Q.; Peng, Y.; Huang, G. Comparison of AFLP and SSR for genetic diversity analysis of Brassica napus hybrids. J. Agric. Sci. 2011, 3, 101–110. [Google Scholar] [CrossRef]
- Provan, J.; Love, H.M.; Maggs, C.A. Development of microsatellites for the invasive riparian plant Impatiens glandulifera (Himalayan balsam) using inter simple sequence repeat cloning. Mol. Ecol. Notes 2007, 7, 451–453. [Google Scholar] [CrossRef]
- Walker, N.F.; Hulme, P.E.; Hoelzel, A.R. Population genetics of an invasive riparian species, Impatiens glandulifera. Plant Ecol. 2009, 203, 243–252. [Google Scholar] [CrossRef]
- Skrede, I.; Borgen, L.; Brochmann, C. Genetic structuring in three closely related circumpolar plant species: AFLP versus microsatellite markers and high-arctic versus arctic–alpine distributions. Heredity 2009, 102, 293–302. [Google Scholar] [CrossRef]
- Kupcinskiene, E.; Zybartaite, L.; Paulauskas, A. Comparison of genetic diversity of three Impatiens species from Central Europe and Baltic region. Zemdirb.-Agric 2015, 102, 87–94. [Google Scholar] [CrossRef]
- Patamsytė, J.; Čėsnienė, T.; Naugžemys, D.; Kleizaitė, V.; Tunaitienė, V.; Rančelis, V.; Žvingila, D. Different habitats show similar genetic structure of Bunias orientalis L. (Brassicaceae) in Lithuania. Not. Bot. Horti Agrobot. Cluj Napoca 2013, 41, 396–403. [Google Scholar] [CrossRef]
- Vyšniauskienė, R.; Naugžemys, D.; Rudaitytė-Lukošienė, E.; Patamsytė, J.; Butkauskas, D.; Kupčinskienė, E.; Žvingila, D. Genetic diversity of Nuphar lutea in Lithuanian river populations. Aquat. Bot. 2020, 161, 103173. [Google Scholar] [CrossRef]
- Vilcinskas, R.; Jociene, L.; Rekasius, T.; Marozas, V.; Paulauskas, A.; Kupcinskiene, E. Genetic diversity of Lithuanian populations of Juniperus communis L. in relation to abiotic and biotic factors. Dendrobiology 2016, 76, 61–71. [Google Scholar] [CrossRef]
- Jørgensen, S.; Mauricio, R. Neutral genetic variation among wild North American populations of the weedy plant Arabidopsis thaliana is not geographically structured. Mol. Ecol. 2004, 13, 3403–3413. [Google Scholar] [CrossRef] [PubMed]
- Lithuanian Hydrometeorological Service under the Ministry of Environment. Available online: http://www.meteo.lt/en/web/guest/climate-regions-of-lithuania (accessed on 28 January 2022).
- Anderson, N.O.; Jocienė, L.; Krokaitė, E.; Rekašius, T.; Paulauskas, A.; Kupčinskienė, E. Genetic diversity of Phalaris arundinacea populations in relation to river regulation in the Merkys basin, Lithuania. River Res. Appl. 2018, 34, 300–309. [Google Scholar] [CrossRef]
- Wagner, V.; Vecera, M.; Jiménez-Alfaro, B.; Pergl, J.; Lenoir, J.; Svenning, J.C.; Pyšek, P.; Agrillo, E.; Biurrun, I.; Campos, J.A.; et al. Alien plant invasion hotspots and invasion debt in European woodlands. J. Veg. Sci. 2021, 32, e13014. [Google Scholar] [CrossRef]
- Lapin, K.; Oettel, J.; Steiner, H.; Langmaier, M.; Sustic, D.; Starlinger, F.; Kindermann, G.; Frank, G. Invasive alien plant species in unmanaged forest reserves, Austria. NeoBiota 2019, 48, 71–96. [Google Scholar] [CrossRef]
- Vojík, M.; Sádlo, J.; Petřík, P.; Pyšek, P.; Man, M.; Pergl, J. Two faces of parks: Sources of invasion and habitat for threatened native plants. Preslia 2020, 92, 353–373. [Google Scholar] [CrossRef]
- Jurová, J.; Matoušková, M.; Wajs-Bonikowska, A.; Kalemba, D.; Renčo, M.; Sedlák, V.; Gogalóvá, Z.; Poráčová, J.; Šalamún, P.; Grulóvá, D. Potential phytotoxic effect of essential oil of non-native species Impatiens parviflora DC. Plants 2019, 8, 241. [Google Scholar] [CrossRef] [PubMed]
- Käfer, J.; Witte, J.P.M. Cover-weighted averaging of indicator values in vegetation analyses. J. Veg. Sci. 2004, 15, 647–652. [Google Scholar] [CrossRef]
- Heinken, T.; Diekmann, M.; Liira, J.; Orczewska, A.; Schmidt, M.; Brunet, J.; Chytrý, M.; Chabrerie, O.; Decocq, G.; De Frenne, P.; et al. The European Forest Plant Species List (EuForPlant): Concept and applications. J. Veg. Sci. 2022, 33, e13132. [Google Scholar] [CrossRef]
- Krokaitė, E.; Shakeneva, D.; Juškaitytė, E.; Tomas, R.; Nemaniūtė-Gužienė, J.; Butkuvienė, J.; Patamsytė, J.; Rančelienė, V.; Vyšniauskienė, R.; Duchovskienė, L.; et al. Nitrogen concentration of the aquatic plant species in relation to land cover type and other variables of the environment. Zemdirb.-Agric. 2019, 106, 203–212. [Google Scholar] [CrossRef]
- Piao, S.; Liu, Q.; Chen, A.; Janssens, I.A.; Fu, Y.; Dai, J.; Lingli, L.; Xu, L.; Miaogen, S.; Zhu, X. Plant phenology and global climate change: Current progresses and challenges. Glob. Change Biol. 2019, 25, 1922–1940. [Google Scholar] [CrossRef]
- Kettenring, K.M.; Menuz, D.R.; Mock, K.E. The nativity and distribution of the cryptic invader Phalaris arundinacea (reed canarygrass) in riparian areas of the Columbia and Missouri River Basins. Wetlands 2019, 39, 55–66. [Google Scholar] [CrossRef]
- Bublyk, O.; Andreev, I.; Kalendar, R.; Spiridonova, K.; Kunakh, V. Efficiency of different PCR-based marker systems for assessment of Iris pumila genetic diversity. Biologia 2013, 68, 613–620. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of the Primer Pair | Sequence of the Primer 5′→3′ | Annealing T (°C) | Size of DNA Fragments (bp) | Total Number of DNA Fragments | Fragment Numbers Per Population | Mean Number of the Fragments Per Population |
|---|---|---|---|---|---|---|
| EcoRI-AAC-FAM MseI-CTG | GACTGCGTACCAATTCAAC GATGAGTCCTGAGTAACTC | 66 | 51–480 | 40 | 38–40 | 39.3 ± 0.3 * |
| EcoRI-ACG-VIC MseI-CAC | GACTGCGTACCAATTCACG GATGAGTCCTGAGTAACAC | 66 | 55–496 | 35 | 35 | 35.0 |
| EcoRI-ACC-NED MseI-CAC | GACTGCGTACCAATTCACC GATGAGTCCTGAGTAACAC | 66 | 50–490 | 26 | 26 | 26.0 |
| EcoRI-AGG-PET MseI-CAC | GACTGCGTACCAATTCAGG GATGAGTCCTGAGTAACAC | 66 | 80–496 | 13 | 12–13 | 13.0 ± 0.2 |
| EcoRI-AAC-FAM MseI-CTG | GACTGCGTACCAATTCACC GATGAGTCCTGAGTAACTG | 66 | 59–486 | 27 | 27 | 27.0 |
| EcoRI-ACG-VIC MseI-CAT | GACTGCGTACCAATTCACG GATGAGTCCTGAGTAACAT | 66 | 56–435 | 31 | 28–31 | 30.8 ± 0.7 |
| EcoRI-AAG-NED MseI-CAG | GACTGCGTACCAATTCAAG GATGAGTCCTGAGTAACAG | 66 | 51–490 | 21 | 21 | 21.0 |
| EcoRI-AGC-PET MseI-CAC | GACTGCGTACCAATTCAGC GATGAGTCCTGAGTAACAC | 66 | 54–483 | 30 | 30 | 30.0 |
| Population | PLP | h | I |
|---|---|---|---|
| Mean ± CI * | Mean ± CI * | ||
| VVe | 17.0 | 0.078 ± 0.012 | 0.111 ± 0.017 |
| VAP | 25.6 | 0.118 ± 0.014 | 0.167 ± 0.019 |
| Sve | 11.2 | 0.051 ± 0.010 | 0.072 ± 0.014 |
| VZi | 25.6 | 0.118 ± 0.014 | 0.167 ± 0.019 |
| DRa | 17.9 | 0.079 ± 0.012 | 0.113 ± 0.016 |
| Aly | 29.2 | 0.137 ± 0.014 | 0.193 ± 0.020 |
| KVa | 16.1 | 0.073 ± 0.011 | 0.103 ± 0.016 |
| KAS | 34.1 | 0.162 ± 0.015 | 0.227 ± 0.021 |
| KZa | 25.1 | 0.117 ± 0.014 | 0.164 ± 0.019 |
| KMa | 16.6 | 0.077 ± 0.012 | 0.109 ± 0.016 |
| Juo | 14.4 | 0.063 ± 0.011 | 0.090 ± 0.015 |
| Pre | 17.9 | 0.080 ± 0.012 | 0.114 ± 0.017 |
| Nid | 11.7 | 0.052 ± 0.010 | 0.074 ± 0.014 |
| Kar | 18.4 | 0.077 ± 0.011 | 0.111 ± 0.016 |
| Pal | 17.5 | 0.079 ± 0.012 | 0.113 ± 0.017 |
| Pla | 12.1 | 0.053 ± 0.010 | 0.076 ± 0.014 |
| Zag | 12.1 | 0.056 ± 0.010 | 0.079 ± 0.014 |
| Pan | 24.7 | 0.118 ± 0.014 | 0.166 ± 0.020 |
| ATr | 18.4 | 0.083 ± 0.012 | 0.118 ± 0.017 |
| JUp | 24.7 | 0.115 ± 0.014 | 0.162 ± 0.019 |
| Jon | 32.3 | 0.143 ± 0.014 | 0.204 ± 0.020 |
| Source | df | SS | MS | Est. Var. | % | Φ | p |
|---|---|---|---|---|---|---|---|
| A. | |||||||
| Among populations | 20 | 305.51 | 15 | 0.99 | 9 | ΦPT = 0.087 | 0.001 |
| Within populations | 84 | 869.60 | 10 | 10.35 | 91 | ||
| Total | 104 | 1175.11 | 11.34 | 100 | |||
| B. | |||||||
| Among population groups of different biotopes | 2 | 34.18 | 17 | 0.06 | 1 | ΦPT = 0.009 | 0.132 |
| Among populations within groups | 18 | 271.33 | 15 | 0.94 | 8 | ΦPR = 0.084 | 0.001 |
| Within populations | 84 | 869.60 | 10 | 10.35 | 91 | ΦRT = 0.089 | 0.001 |
| Total | 104 | 1175.11 | 11.36 | 100 | |||
| C. | |||||||
| Among population groups of different geographical zones | 2 | 33.66 | 17 | 0.05 | 0 | ΦPT = 0.004 | 0.142 |
| Among populations within groups | 18 | 271.84 | 15 | 0.95 | 8 | ΦPR = 0.084 | 0.001 |
| Within populations | 84 | 869.60 | 10 | 10.35 | 91 | ΦRT = 0.088 | 0.001 |
| Total | 104 | 1175.11 | 11.35 | 100 | |||
| D. | |||||||
| Among of population groups besides different road type | 2 | 28.96 | 14 | 0.00 | 0 | ΦPT = −0.002 | 0.679 |
| Among populations within groups | 18 | 276.55 | 15 | 1.00 | 9 | ΦPR = 0.088 | 0.001 |
| Within populations | 84 | 869.60 | 10 | 10.35 | 91 | ΦRT = 0.086 | 0.001 |
| Total | 104 | 1175.11 | 11.36 | 100 | |||
| covIP | L-WEIV | R-WEIV | N-WEIV | L-EIV | R-EIV | N-EIV | |
|---|---|---|---|---|---|---|---|
| Urban forest | |||||||
| Mean | 42.4 | 5.239 | 6.326 | 6.559 | 5.652 | 6.071 | 6.528 |
| CI | 12.0 | 0.199 | 0.346 | 0.203 | 0.180 | 0.465 | 0.325 |
| Riparian forest | |||||||
| Mean | 33.8 | 5.688 | 6.658 | 6.805 | 6.068 | 6.513 | 6.655 |
| CI | 18.5 | 0.219 | 0.308 | 0.241 | 0.225 | 0.439 | 0.394 |
| Agrarian scrubland | |||||||
| Mean | 21.3 | 5.640 | 6.620 | 6.837 | 5.844 | 6.590 | 6.895 |
| CI | 12.1 | 0.311 | 0.254 | 0.284 | 0.212 | 0.348 | 0.255 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jocienė, L.; Stravinskaitė, K.; Krokaitė, E.; Janulionienė, R.; Rekašius, T.; Paulauskas, A.; Marozas, V.; Kupčinskienė, E. AFLP-Based Genetic Structure of Lithuanian Populations of Small Balsam (Impatiens parviflora DC.) in Relation to Habitat Characteristics. Forests 2022, 13, 1228. https://doi.org/10.3390/f13081228
Jocienė L, Stravinskaitė K, Krokaitė E, Janulionienė R, Rekašius T, Paulauskas A, Marozas V, Kupčinskienė E. AFLP-Based Genetic Structure of Lithuanian Populations of Small Balsam (Impatiens parviflora DC.) in Relation to Habitat Characteristics. Forests. 2022; 13(8):1228. https://doi.org/10.3390/f13081228
Chicago/Turabian StyleJocienė, Lina, Kristė Stravinskaitė, Edvina Krokaitė, Rasa Janulionienė, Tomas Rekašius, Algimantas Paulauskas, Vitas Marozas, and Eugenija Kupčinskienė. 2022. "AFLP-Based Genetic Structure of Lithuanian Populations of Small Balsam (Impatiens parviflora DC.) in Relation to Habitat Characteristics" Forests 13, no. 8: 1228. https://doi.org/10.3390/f13081228
APA StyleJocienė, L., Stravinskaitė, K., Krokaitė, E., Janulionienė, R., Rekašius, T., Paulauskas, A., Marozas, V., & Kupčinskienė, E. (2022). AFLP-Based Genetic Structure of Lithuanian Populations of Small Balsam (Impatiens parviflora DC.) in Relation to Habitat Characteristics. Forests, 13(8), 1228. https://doi.org/10.3390/f13081228