
Study on the Genetic Variation of Triadica sebifera (Linnaeus) Small Populations Based on SSR Markers


Abstract
:1. Introduction
2. Materials and Methods
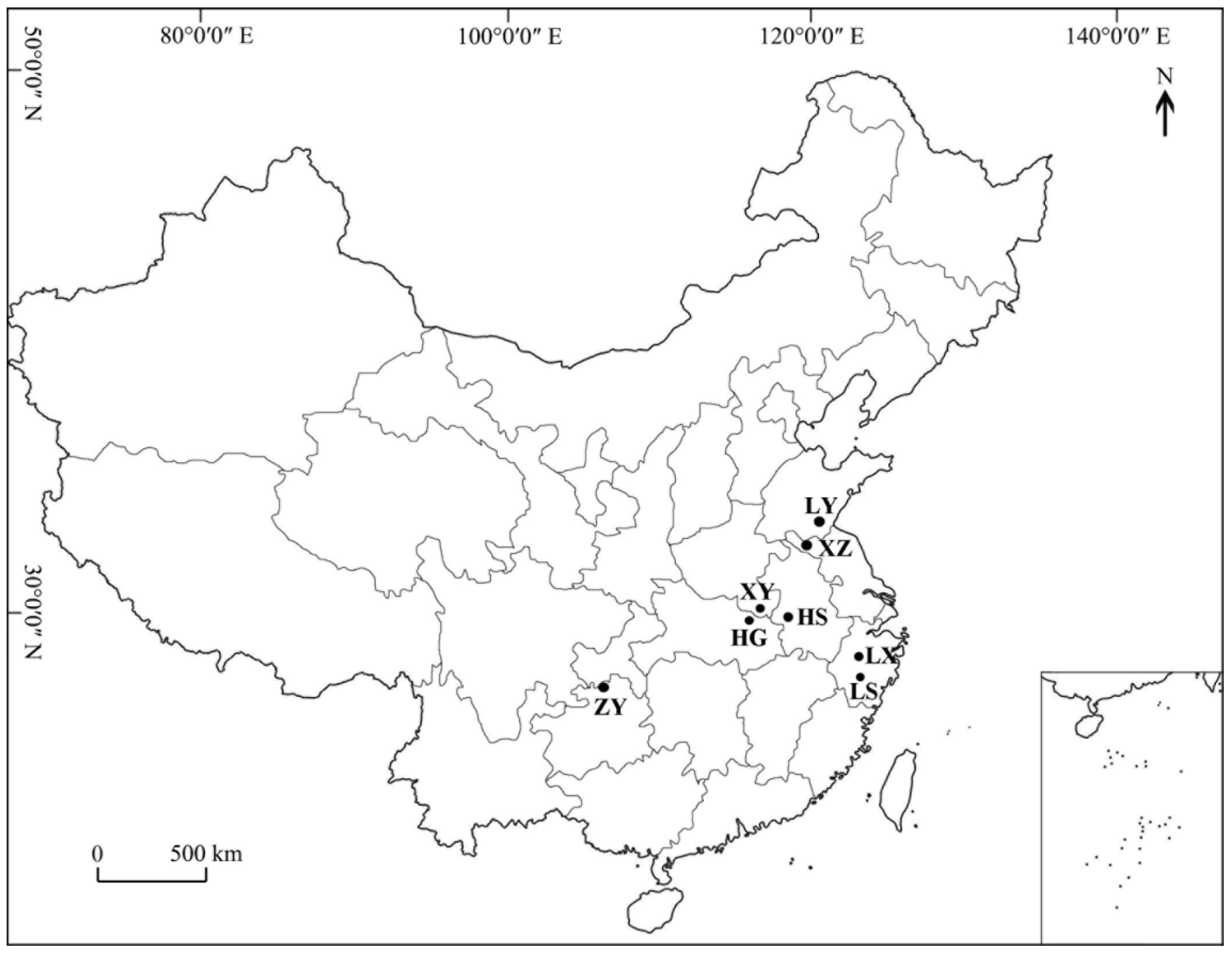
2.1. Sample Information
2.2. Experimental Method
2.3. Data Analysis
3. Results
3.1. Genetic Diversity
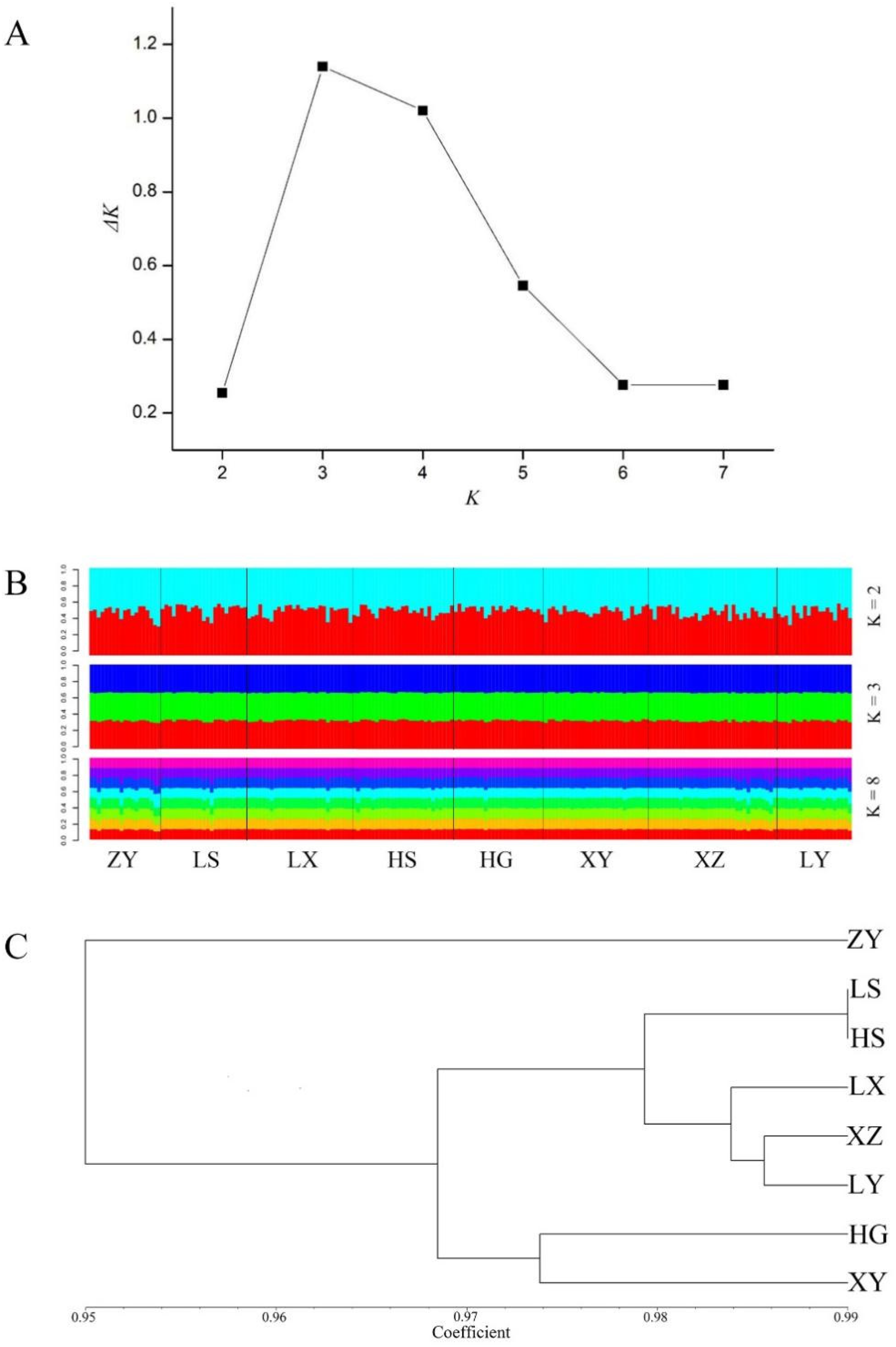
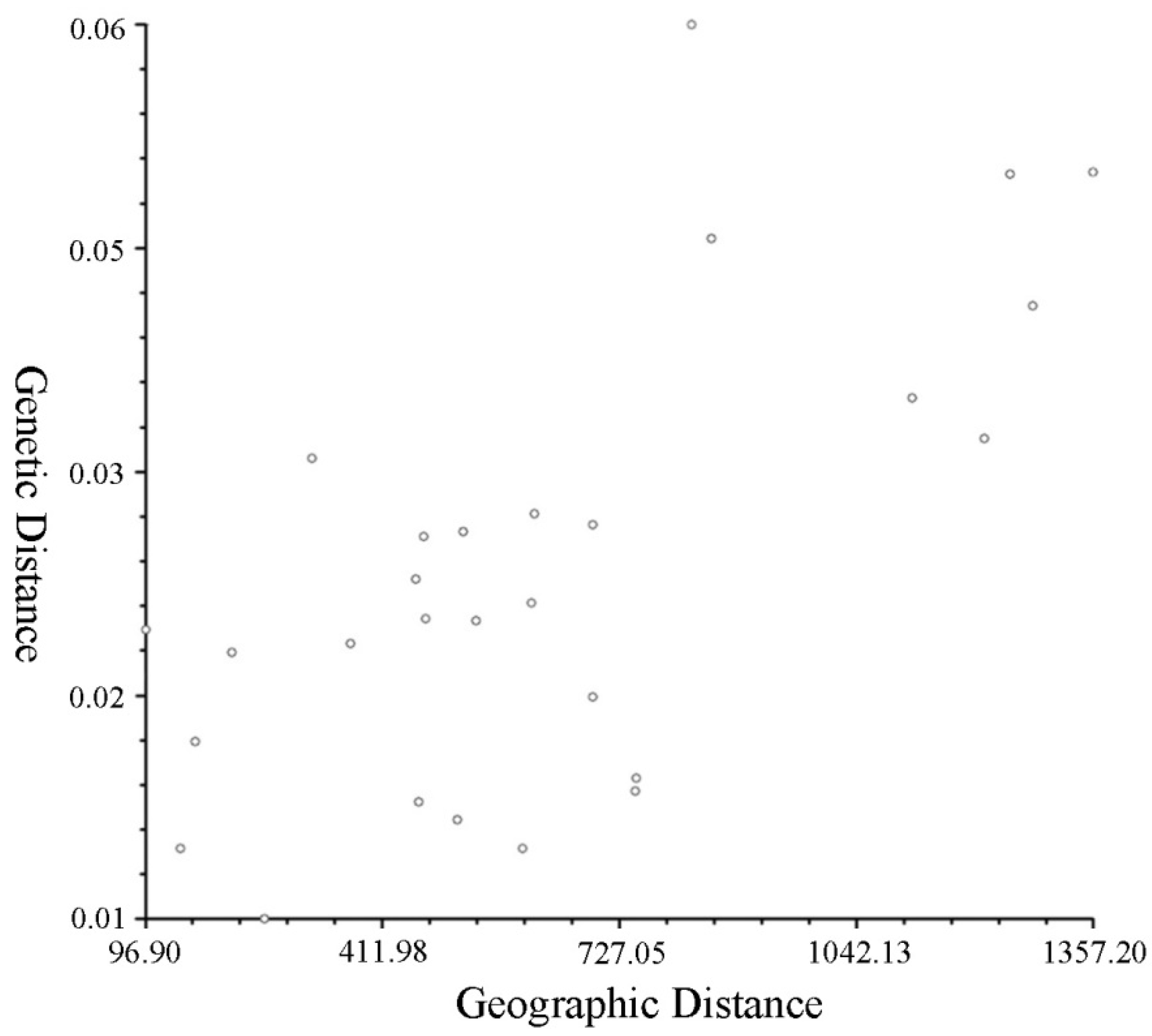
3.2. Genetic Differentiation and Genetic Structure
3.3. Rare Alleles
4. Discussion
4.1. Genetic Diversity
4.2. Genetic Differentiation and Genetic Structure
4.3. Genetic Resource Conservation Strategy of T. sebifera
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jin, D.J.; Huang, H.K.; Tang, R.Q.; Tong, Q.Y.; Shi, D.Y.; Hou, Z.S. Investigation and research on the resources of Triadica sebifera varieties in China. Guangxi Plants 1997, 4, 345–362. [Google Scholar]
- Li, D.L.; Huang, D.; Wang, J.; Jin, Y.Q. A review of research on Triadica sebifera. Jiangsu For. Technol. 2009, 36, 43–47. [Google Scholar]
- Zheng, K.; Zhao, L.; Lang, N.J.; Peng, M.J. Research progress on germplasm resources and cultivation of Triadica sebifera, a woody biomass energy tree. Biomass Chem. Eng. 2006, S1, 360–366. [Google Scholar]
- Wang, S.; Chen, Y.; Yang, Y.; Wu, W.; Liu, Y.; Fan, Q.; Zhou, R. Phylogenetic relationships and natural hybridization in Triadica inferred from nuclear and chloroplast DNA analyses. Biochem. Syst. Ecol. 2016, 64, 142–148. [Google Scholar] [CrossRef]
- DeWalt, S.J.; Siemann, E.; Rogers, W.E. Microsatellite markers for an invasive tetraploid tree, Chinese tallow (Triadica sebifera). Mol. Ecol. Notes 2006, 6, 505–507. [Google Scholar] [CrossRef]
- Gao, R.; Su, Z.; Yin, Y.; Sun, L.; Li, S. Germplasm, chemical constituents, biological activities, utilization, and control of Chinese tallow (Triadica sebifera (L.) Small). Biol. Invasions 2016, 18, 809–829. [Google Scholar] [CrossRef]
- Zhi, Y.; Taylor, M.C.; Campbell, P.M.; Warden, A.C.; Shrestha, P.; El Tahchy, A.; Rolland, V.; Vanhercke, T.; Petrie, J.R.; White, R.G.; et al. Comparative lipidomics and proteomics of lipid droplets in the mesocarp and seed tissues of Chinese tallow (Triadica sebifera). Front. Plant Sci. 2017, 8, 1339. [Google Scholar] [CrossRef]
- Feng, Y.; Luo, J.X.; Du, B.; Gu, Y.J.; Wang, H.L.; Xiang, Q. Study on the phenotypic diversity of seed traits in natural populations of Triadica sebifera in Sichuan-Chongqing area. Sichuan For. Technol. 2011, 32, 19–27. [Google Scholar] [CrossRef]
- Yang, J. Investigation on the Growth of Introduced Triadica sebifera in Rizhao City. Shandong For. Technol. 1996, 2, 17–18. [Google Scholar]
- Wang, H.; Yang, J. Analysis of Ecological Adaptation of Sapium sabiferum Introduced to Shandong Province. Chin. J. Ecol. 1997, 16, 17–19+27. [Google Scholar]
- Li, M.; Li, C.Z.; Wang, L.Y.; Jiang, L.J. Growth characteristics of Triadica sebifera from different provenances at seedling stage. Econ. For. Res. 2012, 30, 75–79. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, X.G.; Du, K.B.; Zhang, Y.Q.; Luo, Z.J.; Tu, B.K. Correlation analysis among several quantitative characters of Triadica sebifera. Econ. For. Res. 2010, 28, 61–66. [Google Scholar] [CrossRef]
- He, X.D.; Zheng, J.W.; Sun, C.; He, K.Y.; Wang, B.S. Construction of fingerprints for 33 varieties in Salicaceae. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2021, 45, 35–42. [Google Scholar] [CrossRef]
- Yuan, J.L.; Ma, J.X.; Zhong, Y.B.; Yue, J.J. SSR-based hybrid identification, genetic analyses and fingerprint development of hybridization progenies from sympodial bamboo (Bambusoideae, Poaceae). J. Nanjing For. Univ. (Nat. Sci. Ed.) 2021, 45, 10–18. [Google Scholar] [CrossRef]
- Li, F.; Gan, S.; Zhang, Z.; Weng, Q.; Xiang, D.; Li, M. Microsatellite-based Genotyping of the Commercial Eucalyptus Clones Cultivated in China. Silvae Genet. 2011, 60, 216. [Google Scholar] [CrossRef] [Green Version]
- DeWalt, S.J.; Siemann, E.; Rogers, W.E. Geographic distribution of genetic variation among native and introduced populations of Chinese tallow tree, Triadica sebifera (Euphorbiaceae). Am. J. Bot. 2011, 98, 1128–1138. [Google Scholar] [CrossRef]
- Cota-Sánchez, J.H.; Remarchuk, K.; Ubayasena, K. Ready-to-use DNA extracted with a CTAB method adapted for herbarium specimens and mucilaginous plant tissue. Plant Mol. Biol. Rep. 2006, 24, 161–167. [Google Scholar] [CrossRef]
- Palero, F.; González-Candelas, F.; Pascual, M. MICROSATELIGHT--pipeline to expedite microsatellite analysis. J. Hered. 2011, 102, 247–249. [Google Scholar] [CrossRef]
- Nei, M. Definition and Estimation of Fixation Indices. Evolution 1986, 40, 643–645. [Google Scholar] [CrossRef]
- Liu, K.J.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [Green Version]
- Goudet, J. FSTAT, A Program to Estimate and Test Gene Diversities and Fixation Indices, version 2.9.3. 2001. Available online: https://www.unil.ch/popgen/softwares/fstat.Htm (accessed on 18 October 2021).
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 64, 1567–1587. [Google Scholar] [CrossRef]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, N.A. Distruct: A program for the graphical display of population structure. Mol. Ecol Notes 2004, 4, 137–138. [Google Scholar] [CrossRef]
- Rohlf, F.J. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System, version 2.2, 2.1; Department of Ecoloy and Evolution, State University of New York: New York, NY, USA, 2005. [Google Scholar]
- Zhou, Q.; Mu, K.; Ni, Z.; Liu, X.; Li, Y.; Xu, L.-A. Analysis of genetic diversity of ancient Ginkgo populations using SSR markers. Ind. Crops Prod. 2020, 145, 111942. [Google Scholar] [CrossRef]
- Shi, X.; Wen, Q.; Cao, M.; Guo, X.; Xu, L.A. Genetic diversity and structure of natural Quercus variabilis population in China as revealed by microsatellites markers. Forests 2017, 8, 495. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Xu, H.; Xiao, F.; Sun, S.; Lou, Y.; Zou, Y.; Xu, X. Genetic diversity and paternity analyses in a 1.5th generation seed orchard of Chenshan red-heart Chinese fir. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2021, 45, 87–92. [Google Scholar] [CrossRef]
- Li, H.; Chen, L.; Liang, C.; Huang, M. Experimental study on provenance of Liriodendron species. For. Sci. Technol. Dev. 2005, 05, 13–16. [Google Scholar]
- Zhou, P.-Y.; Hui, L.-X.; Huang, S.-J.; Ni, Z.-X.; Yu, F.-X.; Xu, L.-A. Study on the Genetic Structure Based on Geographic Populations of the Endangered Tree Species: Liriodendron chinense. Forests 2021, 12, 917. [Google Scholar] [CrossRef]
- Feng, Y.H.; Li, H.G.; Yang, Z.Q.; Wu, D.S. Study on the genetic structure of natural populations of Pinus massoniana provenance. Guangxi Plant 2016, 36, 1275–1281, 1395. [Google Scholar]
- Qi, M.; Zhou, Q.; Ni, Z.; Wu, Y.; Han, X.; Xu, L. Genetic structure analysis of ancient Ginkgo biloba L. populations based on SSR markers. Chin. J. Ecol. 2019, 38, 2902–2910. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population Code | Location | Longitude (E) | Latitude (N) | Altitude (m a.s.l.) | Annual Rainfall (mm) | N |
|---|---|---|---|---|---|---|
| ZY | Zunyi, Guizhou | 27°41′57″ | 106°54′41″ | 209 | 1200 | 19 |
| HG | Huanggang, Hubei | 31°26′38″ | 114°24′06″ | 124 | 1400 | 24 |
| XY | Xinyang, Henan | 31°39′35″ | 115°23′35″ | 229 | 1200 | 28 |
| HS | Huangshan, Anhui | 30°01′16″ | 118°0′7″ | 224 | 2395 | 27 |
| LY | Linyi, Shandong | 35°15′40″ | 117°58′19″ | 174 | 840 | 20 |
| XZ | Xuzhou, Jiangsu | 34°16′37″ | 118°26′41″ | 198 | 876 | 34 |
| LX | Lanxi, Zhejiang | 29°19′19″ | 119°41′56″ | 98 | 1158 | 28 |
| LS | Lishui, Zhejiang | 28°41′18″ | 119°17′08″ | 326 | 1350 | 23 |
| Total | 203 |
| Locus Code | Repeat Motif | Primer Sequence (5′~3′) | Fragment Size (bp) | TM (°C) |
|---|---|---|---|---|
| E-SSR25 | (AAG)10 | AGGTTGACGACTTCTGTGTT | 351 | 53 °C |
| AGTTAGCCTGACCATTTCTC | ||||
| E-SSR29 | (CT)11 | ACCTTGCGAATGTTTATCC | 339 | 50 °C |
| GGGGAAAAACAGATGGAAT | ||||
| E-SSR52 | (AG)12 | CTTTTACCTTTGATGTCGG | 491 | 53 °C |
| GTTTCGGCAATTTCTCTGT | ||||
| E-SSR53 | (AG)10 | AAACAAGTGAAGTGCCCAT | 380 | 51 °C |
| TTAGCCCAGCCCATTATTA | ||||
| E-SSR55 | (TCT)10 | GCGTACCTTCTTCAATGCTC | 428 | 53 °C |
| TTCAACTTCTCTTTCCGTCA | ||||
| E-SSR58 | (AGA)11 | TCCACCTAGCGAAGTTTTG | 295 | 52 °C |
| TTGATTCCTCCCCTTGTTT | ||||
| E-SSR61 | (AAG)12 | GGTTTCTTTTGCTCTCTTC | 279 | 50 °C |
| CCGGTTACTGCATTTCATA | ||||
| E-SSR85 | (CT)10 | TTGCTCTTGGGACCTATTA | 290 | 50 °C |
| TTCTTCCCTTGTGAGTTGT | ||||
| E-SSR103 | (TC)10 | CTACCCAATCACCTCTTTC | 287 | 50 °C |
| TTCTTCTCTGTTCTGGCTC | ||||
| E-SSR106 | (AGG)10 | TCCCAGTTGACTGACGAACA | 197 | 55 °C |
| CGAGGGTGAGGTCAGAGAAG |
| Locus | Na | Ne | He | I | PIC | Fst | Nm | HWE |
|---|---|---|---|---|---|---|---|---|
| E-SSR25 | 5 | 2.411 | 0.587 | 1.043 | 0.520 | 0.042 | 5.706 | ** |
| E-SSR29 | 5 | 1.662 | 0.399 | 0.730 | 0.329 | 0.037 | 6.484 | * |
| E-SSR52 | 3 | 2.084 | 0.521 | 0.797 | 0.408 | 0.017 | 14.091 | ** |
| E-SSR53 | 5 | 1.929 | 0.483 | 0.861 | 0.427 | 0.014 | 18.192 | ** |
| E-SSR55 | 4 | 2.208 | 0.5 48 | 0.891 | 0.446 | 0.024 | 10.312 | ** |
| E-SSR58 | 4 | 1.410 | 0.292 | 0.598 | 0.269 | 0.038 | 6.389 | |
| E-SSR61 | 4 | 1.685 | 0.408 | 0.762 | 0.370 | 0.022 | 10.910 | ** |
| E-SSR85 | 5 | 2.899 | 0.657 | 1.214 | 0.614 | 0.039 | 6.239 | ** |
| E-SSR103 | 3 | 1.930 | 0.483 | 0.798 | 0.443 | 0.019 | 13.279 | ** |
| E-SSR106 | 5 | 2.124 | 0.531 | 0.846 | 0.417 | 0.012 | 19.911 | ** |
| Mean | 4.3 | 2.034 | 0.491 | 0.854 | 0.424 | 0.026 | 11.151 |
| Population Code | Na | Ne | AR | Ho | He | F |
|---|---|---|---|---|---|---|
| ZY | 2.9 | 2.047 | 2.890 | 0.563 | 0.492 | −0.146 |
| HG | 3.3 | 1.974 | 3.293 | 0.613 | 0.497 | −0.233 |
| XY | 3.5 | 2.021 | 3.245 | 0.554 | 0.498 | −0.113 |
| HS | 3.3 | 1.990 | 3.192 | 0.556 | 0.480 | −0.157 |
| LY | 3.2 | 1.989 | 3.415 | 0.595 | 0.485 | −0.228 |
| XZ | 3.4 | 2.079 | 3.185 | 0.641 | 0.503 | −0.276 |
| LX | 3.3 | 1.969 | 3.125 | 0.596 | 0.471 | −0.265 |
| LS | 3.0 | 1.908 | 3.295 | 0.565 | 0.466 | −0.212 |
| Mean | 3.24 | 1.997 | 3.205 | 0.585 | 0.486 | −0.203 |
| Source of Variance | Variance Component | Percentage of Total | p Value |
|---|---|---|---|
| Among regions | 0.155 | 4% | ** |
| Among populations within regions | 0.038 | 1% | |
| Within populations | 3.823 | 95% | ** |
| Total | 4.016 | 100% |
| ZY | LS | LX | HS | HG | XY | XZ | LY | |
|---|---|---|---|---|---|---|---|---|
| ZY | 0 | |||||||
| LS | 0.043 | 0 | ||||||
| LX | 0.051 | 0.025 | 0 | |||||
| HS | 0.038 | 0.009 | 0.019 | 0 | ||||
| HG | 0.059 | 0.026 | 0.026 | 0.035 | 0 | |||
| XY | 0.047 | 0.031 | 0.032 | 0.028 | 0.024 | 0 | ||
| XZ | 0.036 | 0.021 | 0.013 | 0.015 | 0.030 | 0.024 | 0 | |
| LY | 0.051 | 0.017 | 0.016 | 0.014 | 0.027 | 0.031 | 0.013 | 0 |
| E-SSR29-E | E-SSR55-E | E-SSR61-E | E-SSR85-C | E-SSR106-D | E-SSR106-E | |
|---|---|---|---|---|---|---|
| ZY | 0.079 | |||||
| HG | 0.021 | 0.042 | 0.021 | |||
| XY | 0.018 | 0.018 | 0.018 | 0.018 | ||
| HS | ||||||
| LY | 0.075 | |||||
| XZ | 0.044 | |||||
| LX | 0.018 | |||||
| LS | 0.022 | 0.022 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, P.; Zhou, Q.; Dong, F.; Shen, X.; Li, Y. Study on the Genetic Variation of Triadica sebifera (Linnaeus) Small Populations Based on SSR Markers. Forests 2022, 13, 1330. https://doi.org/10.3390/f13081330
Zhou P, Zhou Q, Dong F, Shen X, Li Y. Study on the Genetic Variation of Triadica sebifera (Linnaeus) Small Populations Based on SSR Markers. Forests. 2022; 13(8):1330. https://doi.org/10.3390/f13081330
Chicago/Turabian StyleZhou, Pengyan, Qi Zhou, Fengping Dong, Xin Shen, and Yingang Li. 2022. "Study on the Genetic Variation of Triadica sebifera (Linnaeus) Small Populations Based on SSR Markers" Forests 13, no. 8: 1330. https://doi.org/10.3390/f13081330
APA StyleZhou, P., Zhou, Q., Dong, F., Shen, X., & Li, Y. (2022). Study on the Genetic Variation of Triadica sebifera (Linnaeus) Small Populations Based on SSR Markers. Forests, 13(8), 1330. https://doi.org/10.3390/f13081330