
Variation in Water Uptake Dynamics of Dominant Wood Plants of Pinus taiwanensis Hayata Communities Based on Stable Isotopes

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
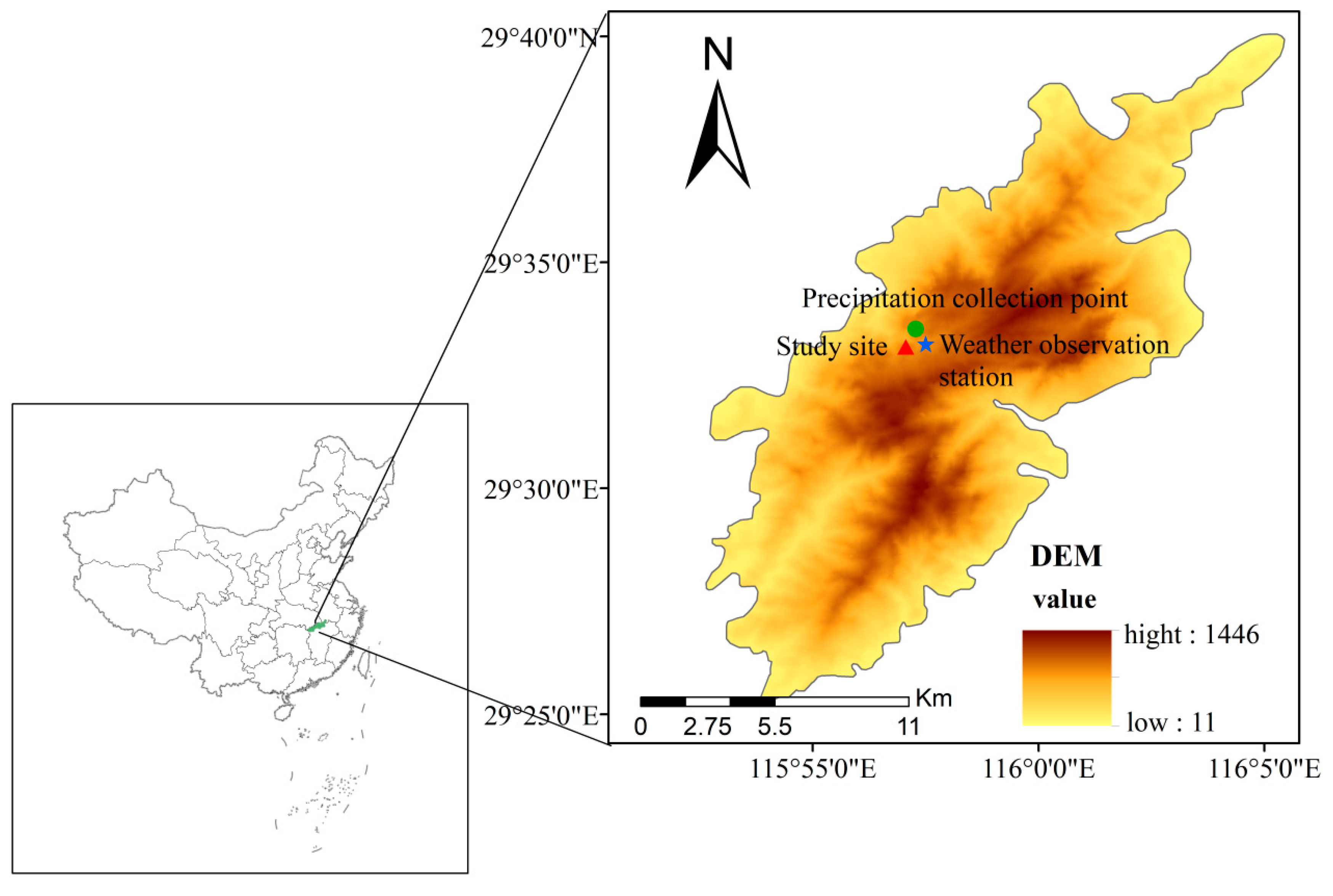
2.1. Study Site
2.2. Experimental Design and Sample Collection
2.3. Sample Analysis and Calculation
3. Results
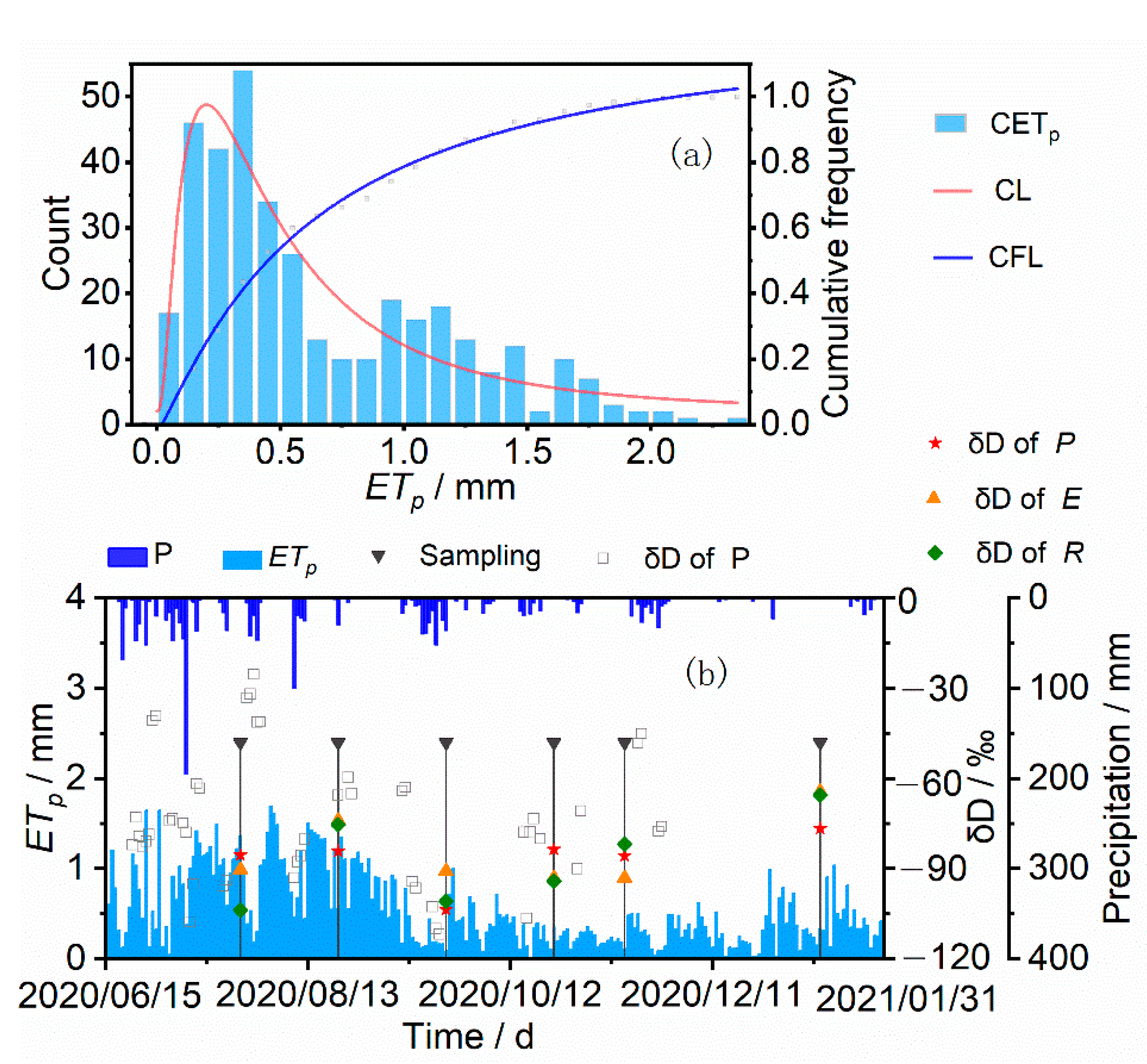
3.1. Variations of Rainfall and Soil Water Content
3.2. Isotopic Composition of Rainfall, Soil Water and Plant Water
3.3. Relationship between Isotopic Compositions in Soil and Xylem Water
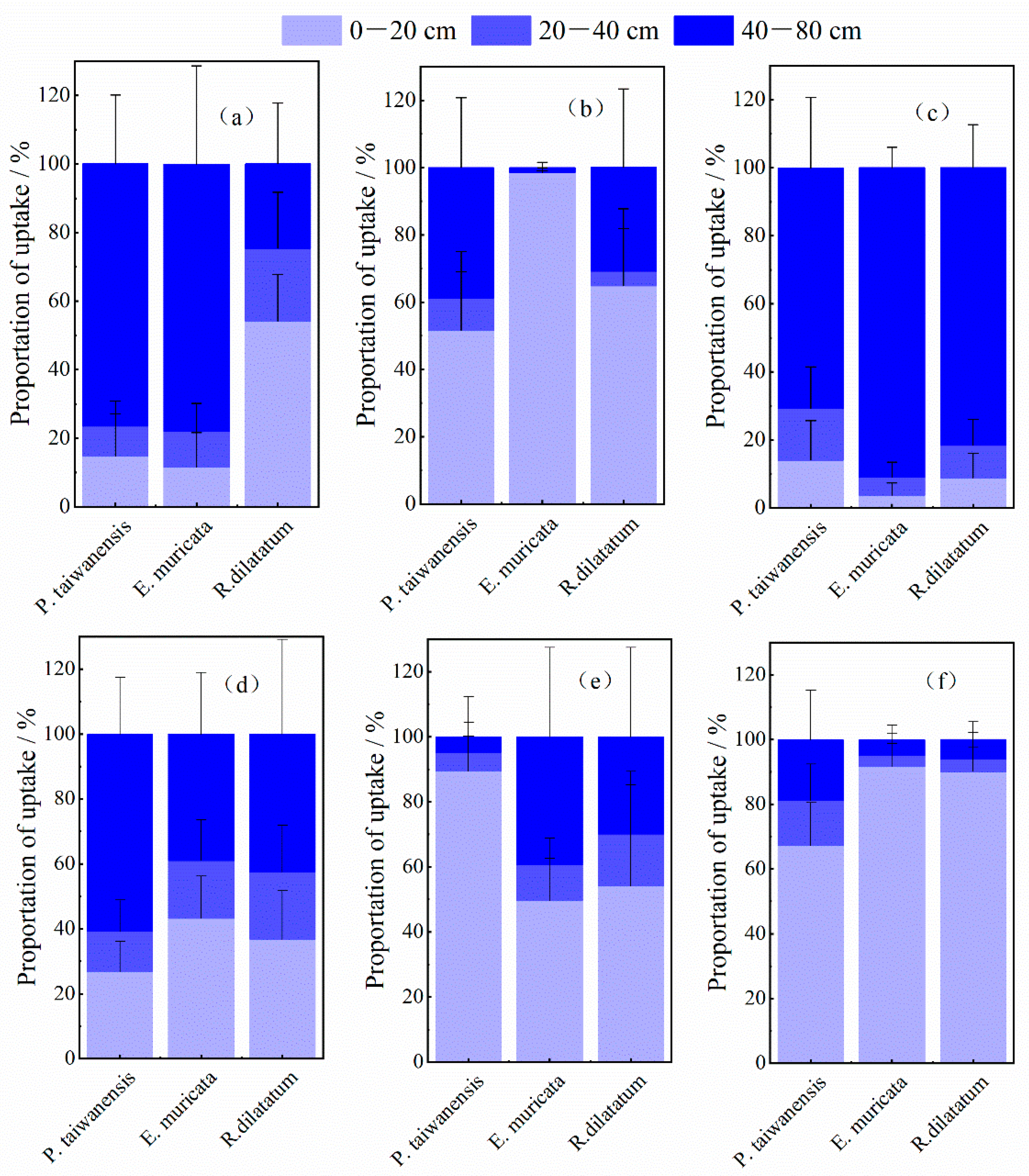
3.4. Water Uptake Patterns of P. taiwanensis and Understory Shrub Species
4. Discussion
4.1. Water Use Patterns of Main Arbor and Shrub Species in P. taiwanensis Community
4.2. Dynamic Niche Partitioning in Water Uptake among P. taiwanensis Community
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, L.; He, B.; Chen, A.; Wang, H.; Liu, J.; Lű, A.; Chen, Z. Drought Dominates the Interannual Variability in Global Terrestrial Net Primary Production by Controlling Semi-Arid Ecosystems. Sci. Rep. 2016, 6, 24639. [Google Scholar] [CrossRef] [Green Version]
- Jolly, W.M.; Cochrane, M.A.; Freeborn, P.H.; Holden, Z.A.; Brown, T.J. Climate-Induced Variations in Global Wildfire Danger from 1979 to 2013. Nat. Commun. 2015, 6, 7537. [Google Scholar] [CrossRef]
- Pedrono, M.; Locatelli, B.; Ezzine-de-Blas, D.; Pesche, D.; Morand, S.; Binot, A. Impact of Climate Change on Ecosystem Services. In Climate Change and Agriculture Worldwide; Torquebiau, E., Ed.; Springer: Dordrecht, The Netherlands, 2016; pp. 251–261. ISBN 978-94-017-7460-4. [Google Scholar]
- Halada, L. Ecosystem Services of Mountains: An Urgent Research Area; ALTERNet: Brussels, Belgium, 2010. [Google Scholar]
- Huber, R.; Bugmann, H.; Buttler, A.; Rigling, A. Sustainable Land-Use Practices in European Mountain Regions under Global Change: An Integrated Research Approach. Ecol. Soc. 2013, 18, 37. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Pomeroy, J.W. Diagnosis of Future Changes in Hydrology for a Canadian Rockies Headwater Basin. Hydrol. Earth Syst. Sci. 2020, 24, 2731–2754. [Google Scholar] [CrossRef]
- Pomeroy, J.; Fang, X.; Ellis, C. Sensitivity of Snowmelt Hydrology in Marmot Creek, Alberta, to Forest Cover Disturbance. Hydrol. Processes 2012, 26, 1891–1904. [Google Scholar] [CrossRef]
- Carroll, C.; Knapp, A.K.; Martin, P.H. Dominant Tree Species of the Colorado Rockies Have Divergent Physiological and Morphological Responses to Warming. For. Ecol. Manag. 2017, 402, 234–240. [Google Scholar] [CrossRef]
- Harder, P.; Pomeroy, J.W.; Westbrook, C.J. Hydrological Resilience of a Canadian Rockies Headwaters Basin Subject to Changing Climate, Extreme Weather, and Forest Management. Hydrol. Processes 2015, 29, 3905–3924. [Google Scholar] [CrossRef]
- Kelsey, K.C.; Redmond, M.D.; Barger, N.N.; Neff, J.C. Species, Climate and Landscape Physiography Drive Variable Growth Trends in Subalpine Forests. Ecosystems 2017, 21, 125–140. [Google Scholar] [CrossRef]
- Carroll, R.W.; Huntington, J.L.; Snyder, K.A.; Niswonger, R.G.; Morton, C.; Stringham, T.K. Evaluating Mountain Meadow Groundwater Response to Pinyon-Juniper and Temperature in a Great Basin Watershed. Ecohydrology 2017, 10, e1792. [Google Scholar] [CrossRef]
- Christensen, C.W.; Hayashi, M.; Bentley, L.R. Hydrogeophysical Survey of Groundwater Flow Pathways in an Alpine Headwater Basin. In Proceedings of the Near Surface Geoscience 2016—22nd European Meeting of Environmental and Engineering Geophysics, Strasbourg, France, 4–8 September 2016. [Google Scholar]
- Harrington, J.S.; Hayashi, M.; Kurylyk, B.L. Influence of a Rock Glacier Spring on the Stream Energy Budget and Cold-Water Refuge in an Alpine Stream. Hydrol. Processes 2017, 31, 4719–4733. [Google Scholar] [CrossRef]
- Hood, J.L.; Hayashi, M. Characterization of Snowmelt Flux and Groundwater Storage in an Alpine Headwater Basin. J. Hydrol. 2015, 521, 482–497. [Google Scholar] [CrossRef]
- Dubbert, M.; Caldeira, M.C.; Dubbert, D.; Werner, C. A Pool-Weighted Perspective on the Two-Water-Worlds Hypothesis. New Phytol. 2019, 222, 1271–1283. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Miguez-Macho, G.; Jobbágy, E.G.; Jackson, R.B.; Otero-Casal, C. Hydrologic Regulation of Plant Rooting Depth. Proc. Natl. Acad. Sci. USA 2017, 114, 10572–10577. [Google Scholar] [CrossRef] [Green Version]
- Schenk, H.J.; Jackson, R.B. Rooting Depths, Lateral Root Spreads and below-Ground/above-Ground Allometries of Plants in Water-Limited Ecosystems. J. Ecol. 2010, 90, 480–494. [Google Scholar] [CrossRef] [Green Version]
- West, A.G.; Dawson, T.E.; February, E.C.; Midgley, G.F.; Bond, W.J.; Aston, T.L. Diverse Functional Responses to Drought in a Mediterranean-Type Shrubland in South Africa. New Phytol. 2012, 195, 396–474. [Google Scholar] [CrossRef]
- Allen, S.T.; Kirchner, J.W.; Braun, S.; Siegwolf, R.T.W.; Goldsmith, G.R. Seasonal Origins of Soil Water Used by Trees. Hydrol. Earth Syst. Sci. 2019, 23, 1199–1210. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, G.; Masini, J.; Goldscheider, N.; Meeks, J.; Lavastre, V.; Celle-Jeanton, H.; Gobat, J.-M.; Hunkeler, D. Determination of Spatiotemporal Variability of Tree Water Uptake Using Stable Isotopes (Δ18O, Δ2H) in an Alluvial System Supplied by a High-Altitude Watershed, Pfyn Forest, Switzerland. Ecohydrology 2014, 7, 319–333. [Google Scholar] [CrossRef]
- Brooks, J.R.; Barnard, H.R.; Coulombe, R.; Mcdonnell, J.J. Ecohydrologic Separation of Water between Trees and Streams in a Mediterranean Climate. Nat. Geosci. 2010, 3, 100–104. [Google Scholar] [CrossRef]
- Dawson, T.E.; Ehleringer, J.R. Streamside Trees That Do Not Use Stream Water. Nature 1991, 350, 335–337. [Google Scholar] [CrossRef]
- Goldsmith, G.R.; Lyssette, E.; Muoz-Villers, L.E.; Holwerda, F.; Mcdonnell, J.J.; Asbjornsen, H.; Dawson, T.E. Stable Isotopes Reveal Linkages among Ecohydrological Processes in a Seasonally Dry Tropical Montane Cloud Forest. Ecohydrology 2012, 5, 779–790. [Google Scholar] [CrossRef]
- Langs, L.E.; Petrone, R.M.; Pomeroy, J.W. A Δ18O and Δ2H Stable Water Isotope Analysis of Subalpine Forest Water Sources under Seasonal and Hydrological Stress in the Canadian Rocky Mountains. Hydrol. Processes 2020, 34, 5642–5658. [Google Scholar] [CrossRef]
- Rossatto, D.R.; da Silveira Lobo Sternberg, L.; Franco, A.C. The Partitioning of Water Uptake between Growth Forms in a Neotropical Savanna: Do Herbs Exploit a Third Water Source Niche? Plant Biol. 2013, 15, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Schwendenmann, L.; Pendall, E.; Sanchez-Bragado, R.; Kunert, N.; Hölscher, D. Tree Water Uptake in a Tropical Plantation Varying in Tree Diversity: Interspecific Differences, Seasonal Shifts and Complementarity: Tree water uptake patterns in a tropical tree plantation. Ecohydrology 2015, 8, 1–12. [Google Scholar] [CrossRef]
- Langs, L.E.; Petrone, R.M.; Pomeroy, J.W. Subalpine Forest Water Use Behaviour and Evapotranspiration during Two Hydrologically Contrasting Growing Seasons in the Canadian Rockies. Hydrol. Processes 2021, 35, e14158. [Google Scholar] [CrossRef]
- Linderholm, H.W. Growing Season Changes in the Last Century. Agric. For. Meteorol. 2006, 137, 1–14. [Google Scholar] [CrossRef]
- Molotch, N.P.; Brooks, P.D.; Burns, S.P.; Litvak, M.; Monson, R.K.; McConnell, J.R.; Musselman, K. Ecohydrological Controls on Snowmelt Partitioning in Mixed-Conifer Sub-Alpine Forests. Ecohydrology 2009, 2, 129–142. [Google Scholar] [CrossRef]
- Smith, W.K.; Germino, M.J.; Johnson, D.M.; Reinhardt, K. The Altitude of Alpine Treeline: A Bellwether of Climate Change Effects. Bot. Rev. 2009, 75, 163–190. [Google Scholar] [CrossRef]
- Song, H.; Han, Q.; Zhang, S. Low-Altitude Boundary of Abies Faxoniana Is More Susceptible to Long-Term Open-Top Chamber Warming in the Eastern Tibetan Plateau. Front. Plant Sci. 2021, 12, 766368. [Google Scholar] [CrossRef]
- Duan, B.; Dong, T.; Zhang, X.; Zhang, Y.; Chen, J. Ecophysiological Responses of Two Dominant Subalpine Tree Species Betula Albo-Sinensis and Abies Faxoniana to Intra- and Interspecific Competition under Elevated Temperature. For. Ecol. Manag. 2014, 323, 20–27. [Google Scholar] [CrossRef]
- Liu, X.; Wang, L. Scientific Survey and Study of Biodiversity on the Lushan Nature Reserve in Jiangxi Province; Science Press: Beijing, China, 2010. [Google Scholar]
- Chen, Q.; Guo, J.; Li, C.; Wang, H.; Wu, C.; Deng, W.; Liu, Y.; Ye, Q.; Li, X. Variation characteristics of stable isotope in precipitation in Mount Lu area. J. Nat. Resour. 2019, 34, 1306–1316. [Google Scholar] [CrossRef]
- Liu, W.; Liao, L.; Liu, Y.; Wang, Q.; Murray, P.J.; Jiang, X.; Zou, G.; Cai, J.; Zhao, X. Effects of Phyllostachys Pubescens Expansion on Underground Soil Fauna Community and Soil Food Web in a Cryptomeria Japonica Plantation, Lushan Mountain, Subtropical China. J. Soils Sediments 2021, 21, 2212–2227. [Google Scholar] [CrossRef]
- Deng, W.; Guo, J.; Zou, Q.; Chen, Q.; Huang, J.; Lu, Y.; Liu, Y. Characteristics of the temporal and spatial distribution of the throughfall in Cryptomeria japonica forest in mount Lu. Acta Ecol. Sin. 2021, 41, 2428–2438. [Google Scholar]
- Yu, F.; Zhang, Z.; Chen, L.; Wang, J.; Shen, Z. Spatial Distribution Characteristics of Soil Organic Carbon in Subtropical Forests of Mountain Lushan, China. Environ. Monit. Assess. 2018, 190, 545. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.T.; Kirchner, J.W.; Goldsmith, G.R. Predicting Spatial Patterns in Precipitation Isotope (δ 2 H and δ 18 O) Seasonality Using Sinusoidal Isoscapes. Geophys. Res. Lett. 2018, 45, 4859–4868. [Google Scholar] [CrossRef] [Green Version]
- Chang, E.; Li, P.; Li, Z.; Xiao, L.; Zhao, B.; Su, Y.; Feng, Z. Using Water Isotopes to Analyze Water Uptake during Vegetation Succession on Abandoned Cropland on the Loess Plateau, China. Catena 2019, 181, 104095. [Google Scholar] [CrossRef]
- Wang, M.; Chen, H.; Zhang, W.; Wang, K. Influencing Factors on Soil Nutrients at Different Scales in a Karst Area. Catena 2019, 175, 411–420. [Google Scholar] [CrossRef]
- Zhao, F.; Li, H.; Li, C.; Cai, Y.; Wang, X.; Liu, Q. Analyzing the Influence of Landscape Pattern Change on Ecological Water Requirements in an Arid/Semiarid Region of China. J. Hydrol. 2019, 578, 124098. [Google Scholar] [CrossRef]
- Wang, J.; Fu, B.; Lu, N.; Zhang, L. Seasonal Variation in Water Uptake Patterns of Three Plant Species Based on Stable Isotopes in the Semi-Arid Loess Plateau. Sci. Total Environ. 2017, 609, 27–37. [Google Scholar] [CrossRef]
- West, A.G.; Patrickson, S.J.; Ehleringer, J.R. Water Extraction Times for Plant and Soil Materials Used in Stable Isotope Analysis. Rapid Commun. Mass Spectrom. 2006, 20, 1317–1321. [Google Scholar] [CrossRef]
- Brinkmann, N.; Seeger, S.; Weiler, M.; Buchmann, N.; Eugster, W.; Kahmen, A. Employing Stable Isotopes to Determine the Residence Times of Soil Water and the Temporal Origin of Water Taken up by Fagus Sylvatica and Picea Abies in a Temperate Forest. New Phytol. 2017, 219, 1300–1313. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Li, Y.; Tan, Z.; Guo, Q.; College of Water Resources Science and Engineering, Taiyuan University of Technology, Taiyuan 030024, China. Variations of water sources for a typical mesophyte vegetation in the Lake Poyang wetland using stable isotopes. J. Lake Sci. 2020, 32, 1749–1760. [Google Scholar] [CrossRef]
- Lyu, S.; Wang, J.; Song, X.; Wen, X. The Relationship of ΔD and Δ18O in Surface Soil Water and Its Implications for Soil Evaporation along Grass Transects of Tibet, Loess, and Inner Mongolia Plateau. J. Hydrol. 2021, 600, 126533. [Google Scholar] [CrossRef]
- Yang, B.; Wen, X.; Sun, X. Seasonal Variations in Depth of Water Uptake for a Subtropical Coniferous Plantation Subjected to Drought in an East Asian Monsoon Region. Agric. For. Meteorol. 2015, 201, 218–228. [Google Scholar] [CrossRef]
- Stock, B.C.; Semmens, B.X. Unifying Error Structures in Commonly Used Biotracer Mixing Models. Ecology 2016, 97, 2562–2569. [Google Scholar] [CrossRef] [PubMed]
- Craig, H. Isotopic Variations in Meteoric Waters. Science 1961, 133, 1702–1703. [Google Scholar] [CrossRef] [PubMed]
- Schenk, H.J.; Jackson, R.B. Mapping the Global Distribution of Deep Roots in Relation to Climate and Soil Characteristics. Geoderma 2005, 126, 129–140. [Google Scholar] [CrossRef]
- Muhammad, A.A. Waterlogging Stress in Plants: A Review. Afr. J. Agric. Res. 2012, 7, 1976–1981. [Google Scholar] [CrossRef]
- Pezeshki, S.R.; DeLaune, R.D. Soil Oxidation-Reduction in Wetlands and Its Impact on Plant Functioning. Biology 2012, 1, 196–221. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Tam, N.F.Y.; Wong, Y.S.; Lu, C.Y. Growth and Physiological Responses of Two Mangrove Species (Bruguiera Gymnorrhiza and Kandelia Candel) to Waterlogging. Environ. Exp. Bot. 2003, 49, 209–221. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, H.; Yu, X.; Jia, G.; Jiang, J. Evidence of Foliar Water Uptake in a Conifer Species. Agric. Water Manag. 2021, 255, 106993. [Google Scholar] [CrossRef]
- Wang, H.; Tetzlaff, D.; Soulsby, C. Hysteretic Response of Sap Flow in Scots Pine (Pinus sylvestris) to Meteorological Forcing in a Humid Low-energy Headwater Catchment. Ecohydrology 2019, 12, e2125. [Google Scholar] [CrossRef]
- Yan, B.; Mao, J.; Dickinson, R.E.; Thornton, P.E.; Shi, X.; Ricciuto, D.M.; Warren, J.M.; Hoffman, F.M. Modelling Tree Stem-water Dynamics over an Amazonian Rainforest. Ecohydrology 2020, 13, e2180. [Google Scholar] [CrossRef]
- Muñoz-Villers, L.E.; Holwerda, F.; Alvarado-Barrientos, M.S.; Geissert, D.R.; Dawson, T.E. Reduced Dry Season Transpiration Is Coupled with Shallow Soil Water Use in Tropical Montane Forest Trees. Oecologia 2018, 188, 303–317. [Google Scholar] [CrossRef]
- Abdallah, M.A.B.; Durfee, N.; Mata-Gonzalez, R.; Ochoa, C.G.; Noller, J.S. Water Use and Soil Moisture Relationships on Western Juniper Trees at Different Growth Stages. Water 2020, 12, 1596. [Google Scholar] [CrossRef]
- Mata-González, R.; Abdallah, M.A.B.; Ochoa, C.G. Water Use by Mature and Sapling Western Juniper (Juniperus Occidentalis) Trees. Rangel. Ecol. Manag. 2021, 74, 110–113. [Google Scholar] [CrossRef]
- Deng, W.; Jia, G.; Liu, Y.; Chen, Q.; Huang, J.; Wen, L.; Zhang, L.; Liu, X.; Jia, J.; Peng, S. Long-Term Study on the Seasonal Water Uptake of Platycladus Orientalis in the Beijing Mountain Area, Northern China. Agric. For. Meteorol. 2021, 307, 108531. [Google Scholar] [CrossRef]
- Huo, G.; Gosme, M.; Gao, X.; Dupraz, C.; Yang, J.; Zhao, X. Dynamics of Interspecific Water Relationship in Vertical and Horizontal Dimensions under a Dryland Apple-Brassica Intercropping System: Quantifying by Experiments and the 3D Hi-SAFe Model. Agric. For. Meteorol. 2021, 310, 108620. [Google Scholar] [CrossRef]
- Guderle, M.; Bachmann, D.; Milcu, A.; Gockele, A.; Bechmann, M.; Fischer, C.; Roscher, C.; Landais, D.; Ravel, O.; Devidal, S.; et al. Dynamic Niche Partitioning in Root Water Uptake Facilitates Efficient Water Use in More Diverse Grassland Plant Communities. Funct. Ecol. 2018, 32, 214–227. [Google Scholar] [CrossRef]
- Xu, Q.; Li, H.; Chen, J.; Cheng, X.; Liu, S.; An, S. Water Use Patterns of Three Species in Subalpine Forest, Southwest China: The Deuterium Isotope Approach. Ecohydrology 2011, 4, 236–244. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, M.; Qu, D.; Duan, W.; Wang, J.; Su, P.; Guo, R. Water Use Strategies of Dominant Species (Caragana Korshinskii and Reaumuria Soongorica) in Natural Shrubs Based on Stable Isotopes in the Loess Hill, China. Water 2020, 12, 1923. [Google Scholar] [CrossRef]
- Nie, Y.; Chen, H.; Ding, Y.; Wang, K. Water Source Segregation along Successional Stages in a Degraded Karst Region of Subtropical China. J. Veg. Sci. 2018, 29, 933–942. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Species | Number of Tree Species | Proportion of Tree Numbers/% | Average Tree Diameter/cm | Average Tree Height/m |
|---|---|---|---|---|
| P. taiwanensis | 177 | 28.78 | 15.95 | 14.06 |
| E. muricata | 31 | 5.04 | 5.37 | 2.82 |
| R. dilatatum | 92 | 14.96 | 4.2 | 3.47 |
| Soil Layer/cm | Clay Content/% | Silt Content/% | Sand Content/% | Saturated Moisture Holding Capacity/% | Field Moisture Holding Capacity/% |
|---|---|---|---|---|---|
| 0–10 | 0.16 ± 0.03b | 9.14 ± 0.95b | 90.70 ± 0.98a | 78.47 ± 0.83a | 49.40 ± 5.78a |
| 10–20 | 0.44 ± 0.02a | 15.43 ± 0.24a | 84.13 ± 0.27b | 74.77 ± 2.79a | 52.73 ± 4.59a |
| 20–40 | 0.24 ± 0.10b | 10.76 ± 2.32b | 89.01 ± 2.42a | 66.57 ± 14.47a | 45.99 ± 6.44a |
| 40–60 | 0.10 ± 0.07b | 7.19 ± 1.30b | 92.71 ± 1.36a | 58.56 ± 14.92a | 40.03 ± 7.64a |
| 60–80 | 0.17 ± 0.04b | 8.57 ± 0.30b | 91.27 ± 0.33a | 51.24 ± 11.26a | 37.37 ± 6.04a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, L.; Peng, Y.; Deng, W.; Liu, Y.; Bai, T.; Zou, Q.; Liu, X.; Zhang, L.; Jia, G. Variation in Water Uptake Dynamics of Dominant Wood Plants of Pinus taiwanensis Hayata Communities Based on Stable Isotopes. Forests 2022, 13, 1336. https://doi.org/10.3390/f13081336
Wen L, Peng Y, Deng W, Liu Y, Bai T, Zou Q, Liu X, Zhang L, Jia G. Variation in Water Uptake Dynamics of Dominant Wood Plants of Pinus taiwanensis Hayata Communities Based on Stable Isotopes. Forests. 2022; 13(8):1336. https://doi.org/10.3390/f13081336
Chicago/Turabian StyleWen, Linsheng, Yun Peng, Wenping Deng, Yuanqiu Liu, Tianjun Bai, Qin Zou, Xiaojun Liu, Ling Zhang, and Guodong Jia. 2022. "Variation in Water Uptake Dynamics of Dominant Wood Plants of Pinus taiwanensis Hayata Communities Based on Stable Isotopes" Forests 13, no. 8: 1336. https://doi.org/10.3390/f13081336
APA StyleWen, L., Peng, Y., Deng, W., Liu, Y., Bai, T., Zou, Q., Liu, X., Zhang, L., & Jia, G. (2022). Variation in Water Uptake Dynamics of Dominant Wood Plants of Pinus taiwanensis Hayata Communities Based on Stable Isotopes. Forests, 13(8), 1336. https://doi.org/10.3390/f13081336