
Leaf Traits and Resource Use Efficiencies of 19 Woody Plant Species in a Plantation in Fangshan, Beijing, China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Methods
2.2.1. Measurement of the Light Response Curves
2.2.2. Determination of Leaf Traits
2.3. Data Processing
2.3.1. Calculation of Leaf Functional Traits and Resource Utilization Efficiency
2.3.2. Data Analysis
3. Results
3.1. Interspecific Differences in Leaf Functional Traits and Resource Use Efficiency
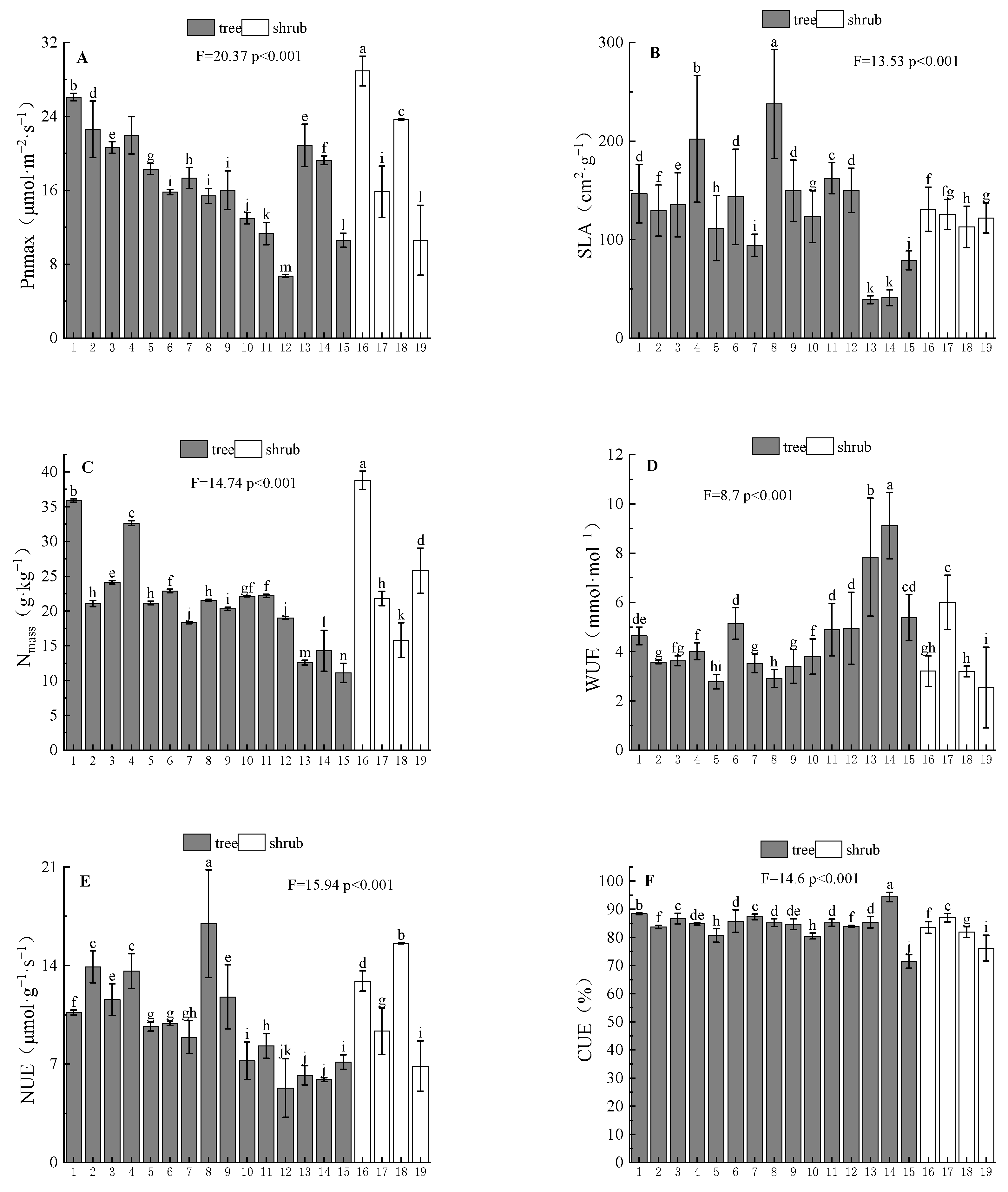
3.1.1. Interspecific Differences of Leaf Functional Traits
3.1.2. Interspecific Differences in Leaf Resource Utilization Efficiency
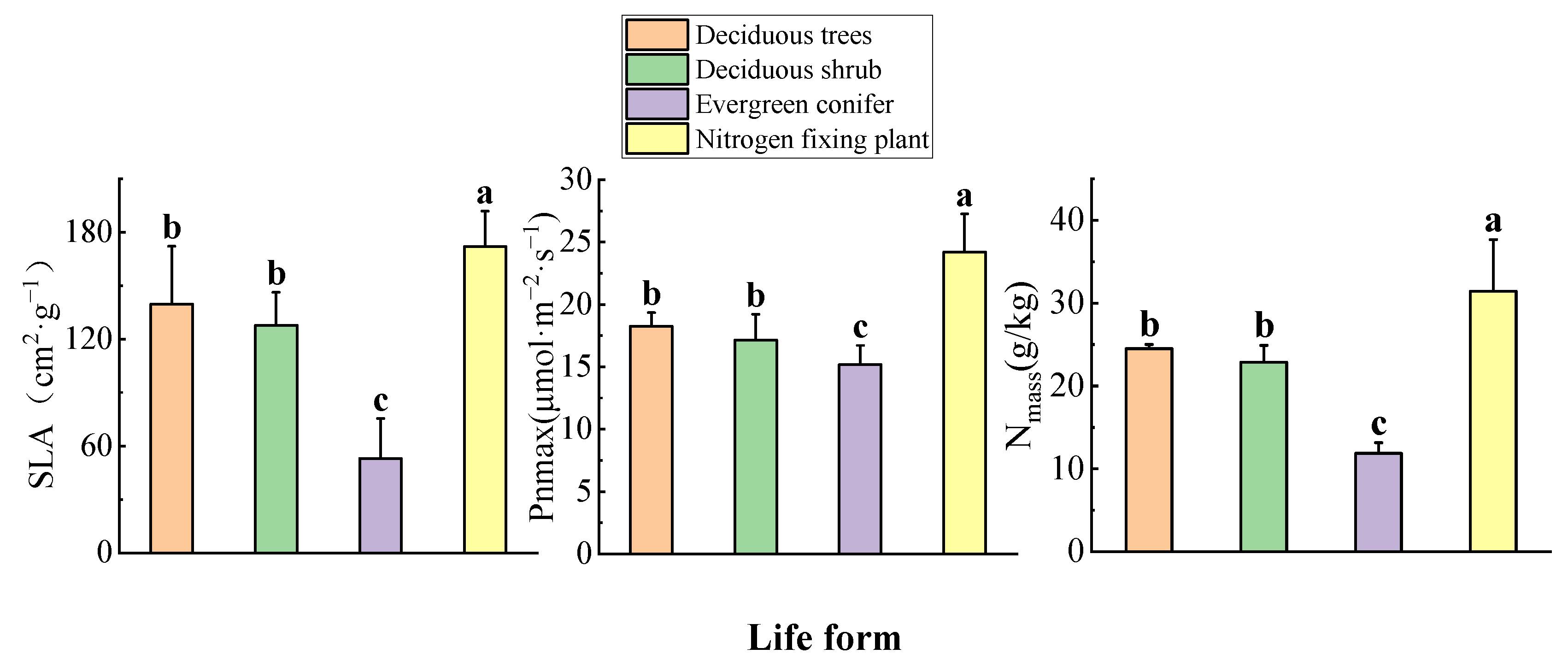
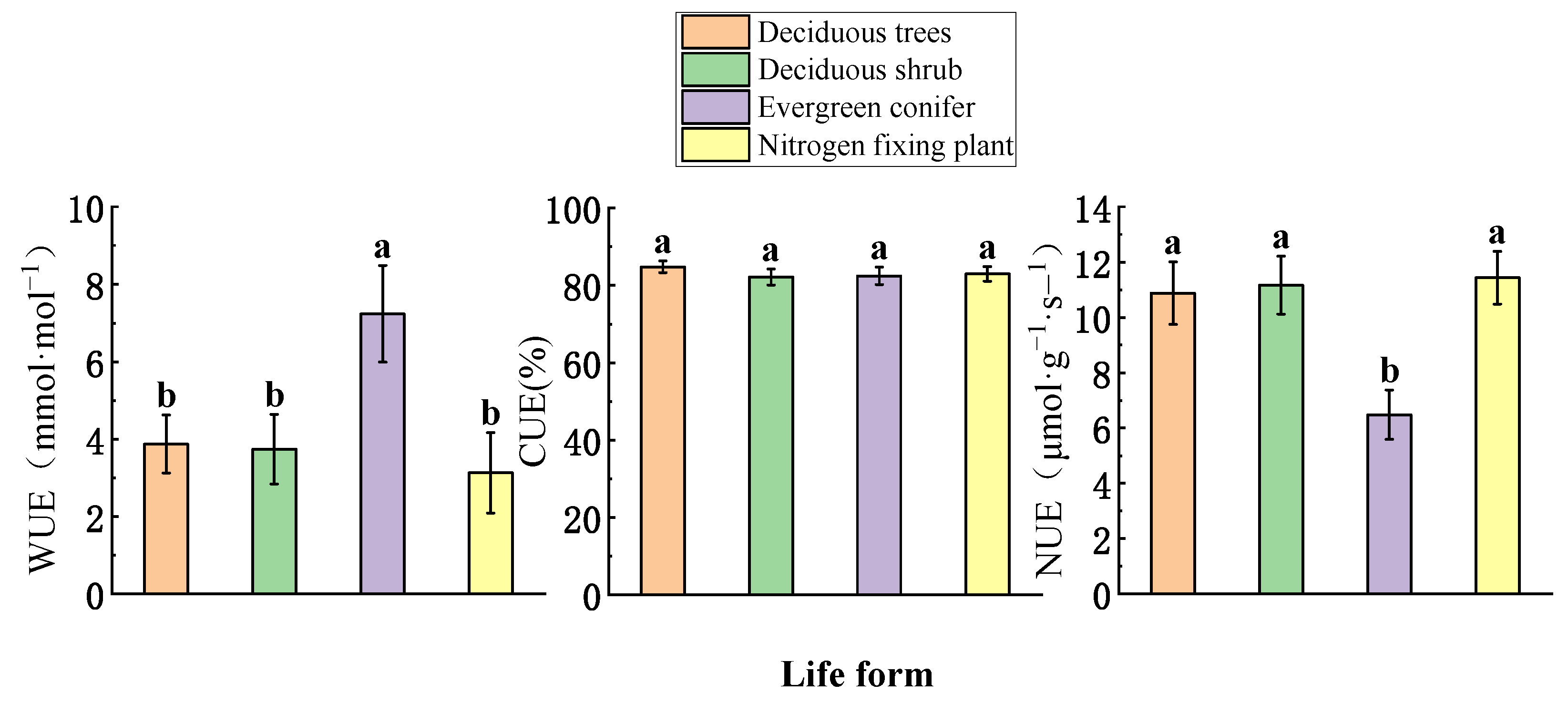
3.2. Differences in Life Forms
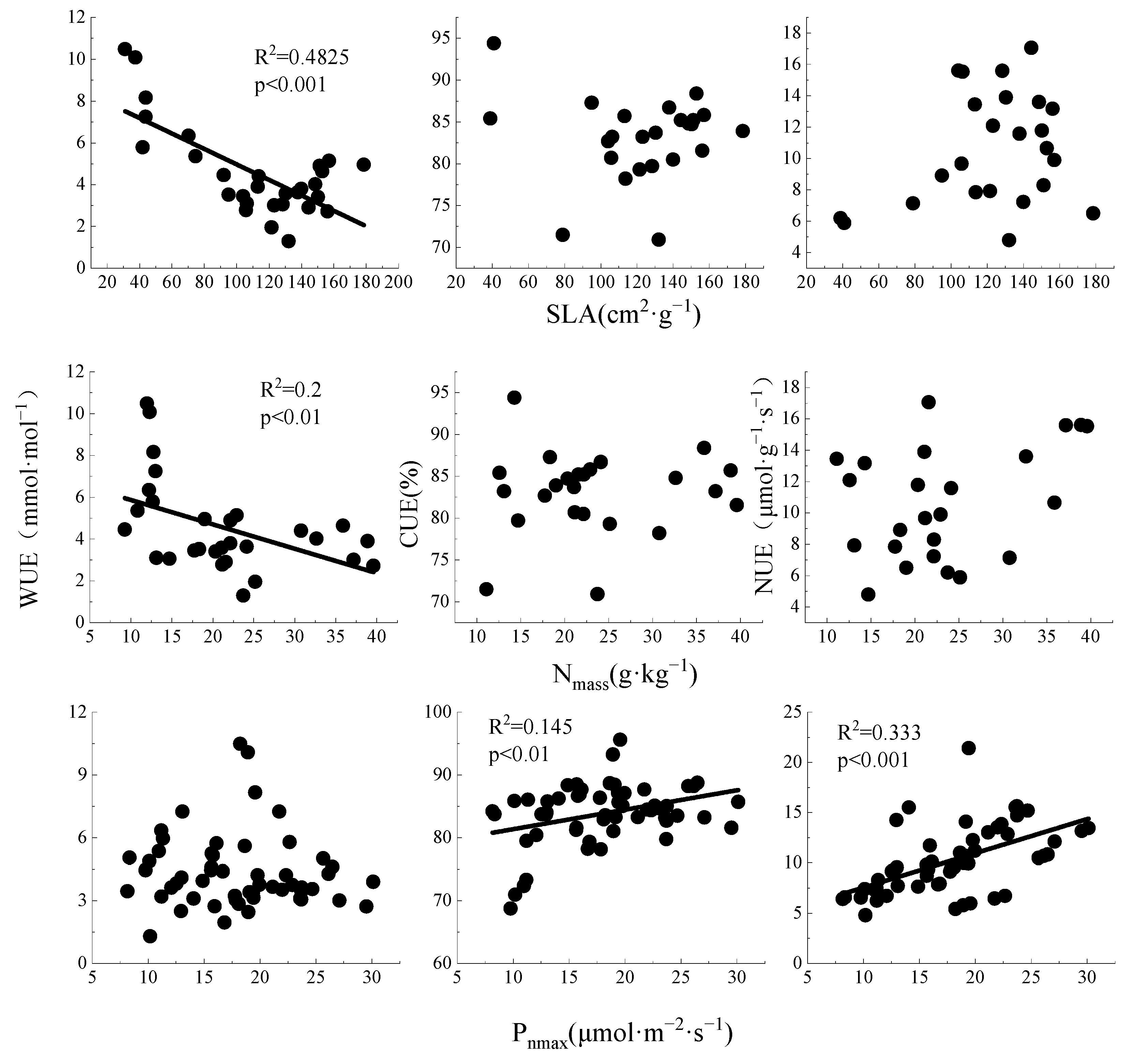
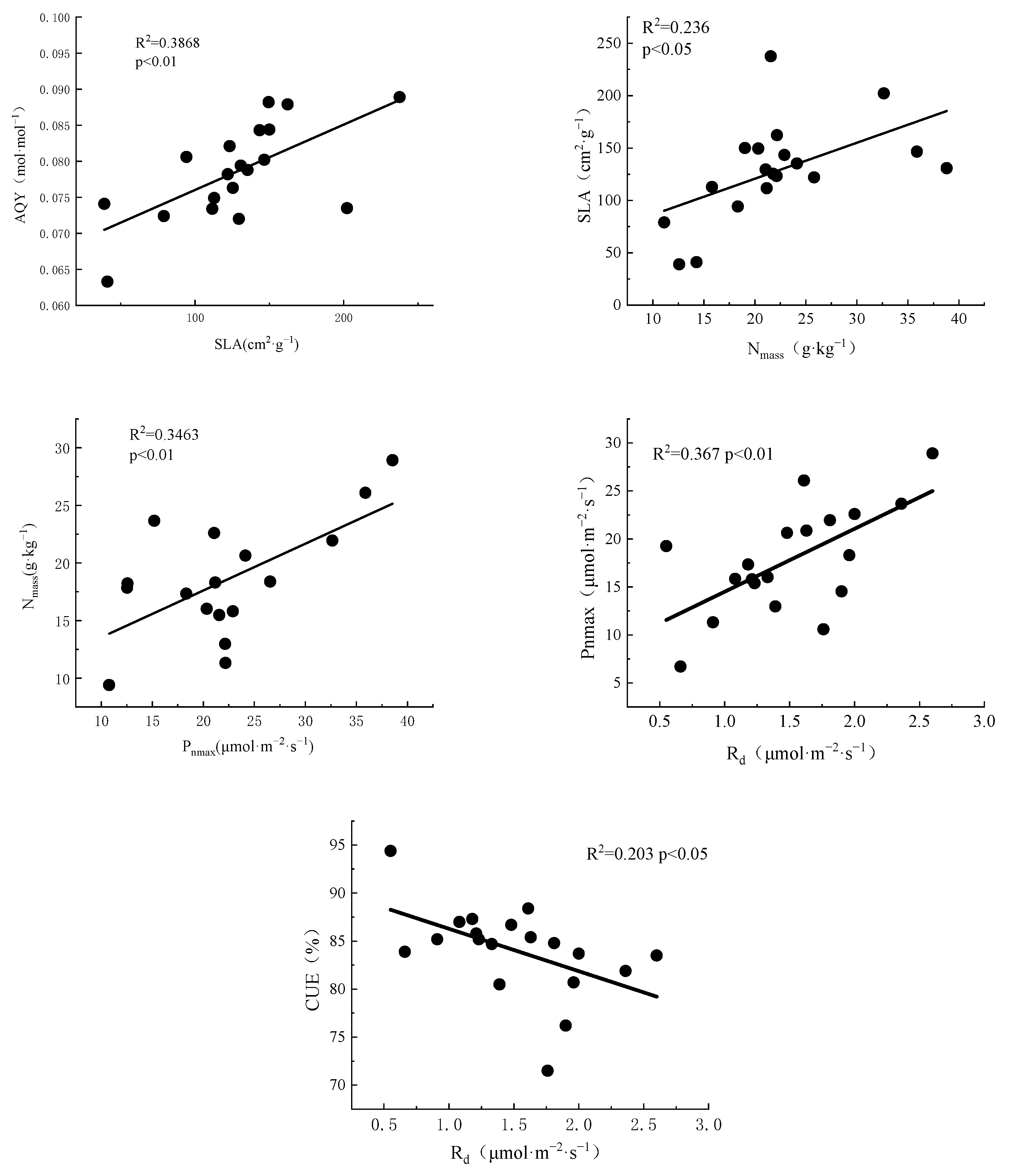
3.3. Correlations between Resource Use Efficiency and Leaf Functional Traits
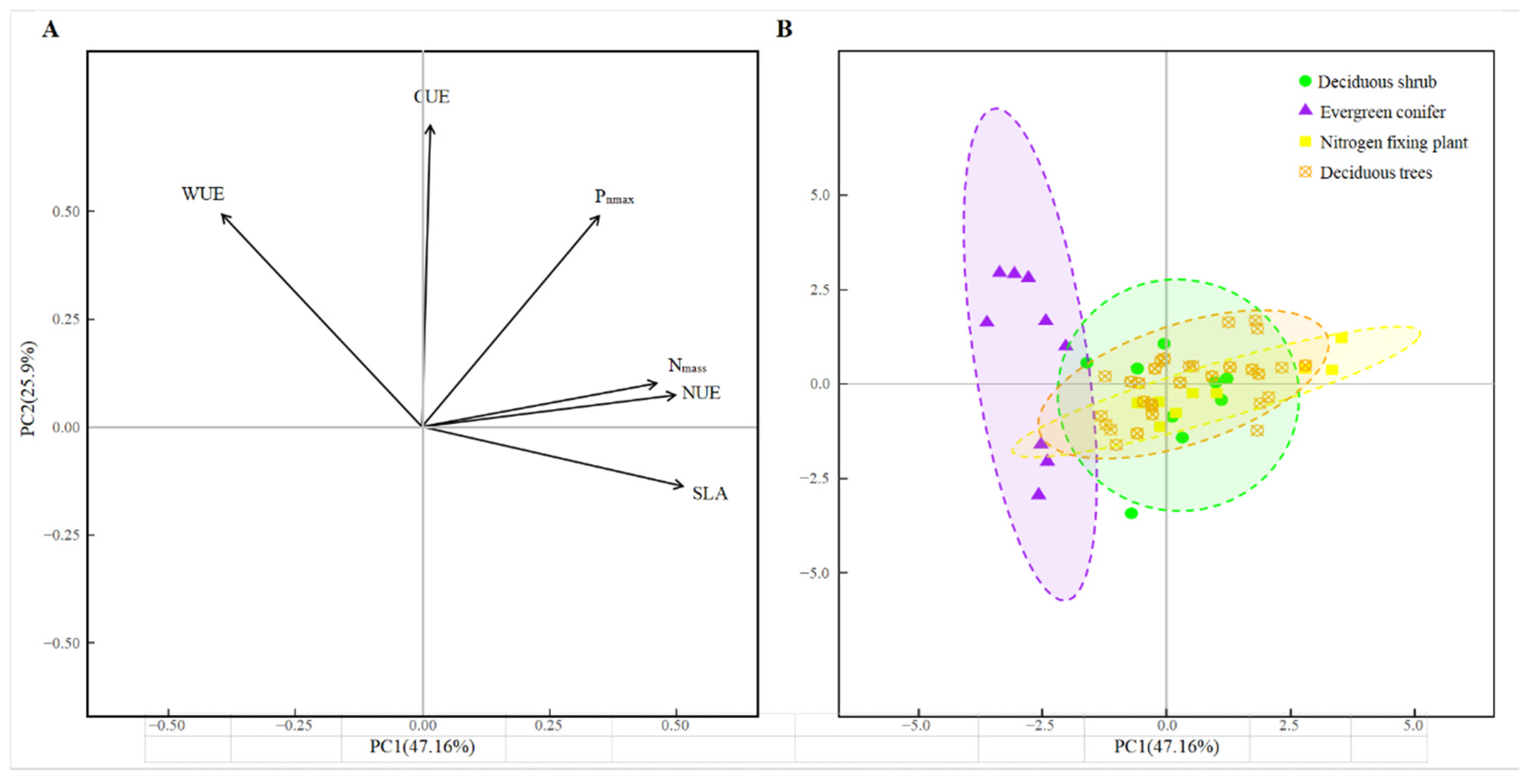
3.4. Principal Component Analyses
4. Discussion
4.1. Differences in Leaf Functional Traits and Resource Utilization Efficiency of Different Species
4.2. Differences in Leaf Functional Traits and Resource Utilization Efficiency among Life Forms
4.3. Correlations between Leaf Functional Traits and Resource Use Efficiency
4.3.1. Correlations between Resource Utilization Efficiency
4.3.2. Correlations between Leaf Functional Traits and Resource Use Efficiency
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reich, P.B.; Cornelissen, H. The world-wide ‘fast-slow’ plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Anderegg, L.D.L.; Berner, L.T.; Badgley, G.; Sethi, M.L.; Law, B.E.; Hille Ris Lambers, J. Within-species patterns challenge our understanding of the leaf economics spectrum. Ecol. Lett. 2018, 21, 734–744. [Google Scholar] [CrossRef]
- Sun, M.; Tian, K.; Zhang, Y.; Wang, H.; Guan, D.X.; Yue, H.T. Research on leaf functional traits and their environmental adaptation. Plant Sci. J. 2017, 35, 940–949. [Google Scholar]
- Petrik, P.; Peter-Petrik, A.; Kurjak, D.; Mukarram, M.; Klein, T.; Gömöry, D.; Střelcová, K.; Frýdl, J.; Konôpková, A. Interannual adjustments in stomatal and leaf morphologicaltraits of European beech (Fagus sylvatica L.) demonstrate itsclimate change acclimation potential. Plant Biol. 2022, 24, 1287–1296. [Google Scholar] [CrossRef]
- Webb, C. Exploring the Phylogenetic Structure of Ecological Communities: An Example for Rain Forest Trees. Am. Nat. 2000, 156, 145–155. [Google Scholar] [CrossRef]
- Yao, T.T.; Meng, T.T.; Ni, J.; Yan, S.; Feng, X.H.; Wang, G.H. Leaf functional trait variation and its relationship with plant phylogenic background and the climate in Xinjiang Junggar Basin, NW China. Biodivers. Sci. 2010, 18, 188–198. [Google Scholar]
- Xu, Y.D.; Dong, S.K.; Li, S.; Shen, H. Research progress on ecological filtering mechanisms for plant community assembly. Acta Ecol. Sin. 2019, 39, 2267–2281. [Google Scholar]
- Liu, J.H.; Zeng, D.H.; Lee, D.K. Leaf traits and their interrelationships of main plant species in southeast Horqin sandy land. Chin. J. Ecol. 2006, 25, 921–925. [Google Scholar]
- Wang, R.L.; Yu, G.R.; He, N.P.; Wang, Q.F.; Zhao, N.; Xu, Z.W. Latitudinal variation of leaf morphological traits from species to communities along a forest transect in eastern China. J. Geogr. Sci. 2016, 26, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Sultan, S.E. Evolutionary Implications of Phenotypic Plasticity in Plants. Evol. Biol. 1987, 138, 213–223. [Google Scholar]
- Smith, E.C.; Griffiths, H.; Wood, L.; Gillon, J. Intra-specific variation in the photosynthetic responses of cyanobiont lichens from contrasting habitats. New Phytol. 1998, 138, 213–224. [Google Scholar] [CrossRef]
- Ackerly, D.D.; Dudley, S.A.; Sultan, S.E.; Schmitt, J.; Coleman, J.S.; Linder, C.R.; Sandquist, D.R.; Geber, M.A.; Evans, A.S.; Dawson, T.E.; et al. The Evolution of Plant Ecophysiological Traits: Recent Advances and Future Directions. BioScience 2000, 50, 979–995. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Meng, T.T.; Ni, J.; Wang, G.H. Plant functional traits, environments and ecosystem functioning. Chin. J. Plant Ecol. 2007, 31, 150–165. [Google Scholar]
- Wang, M.J.; Rong, L.; Li, T.T.; Wang, Q.; Ye, T.M. Studies on leaf functional traits of nine woody lianas in the Karst area of central Guizhou province. J. Trop. Subtrop. Bot. 2021, 29, 455–464. [Google Scholar]
- Wang, Z.Q.; Huang, H.; Wang, H.; Peñuelas, J.; Sardans, J.; Niinemets, Ü.; Niklas, K.J.; Li, Y.; Xie, J.B.; Wright, L.J. Leaf water content contributes to global leaf trait relationships. Nat. Commun. 2022, 13, 5525. [Google Scholar] [CrossRef]
- Li, H.L.; Xu, D.H.; Du, G.Z. Effect of change of plant community composition along degradation gradients on water conditions in an alpine swamp wetland on the Qinghai-Tibetan Plateau of China. Chin. J. Plant Ecol. 2012, 36, 403–410. [Google Scholar] [CrossRef]
- Ren, Q.J.; Li, H.L.; Bu, H.Y. Comparison of physiological and leaf morphological traits for photosynthesis of the 51 plant species in the Maqu alpine swamp meadow. Chin. J. Plant Ecol. 2015, 39, 593–603. [Google Scholar]
- Grime, J.P. Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Dold, C. Water-Use Efficiency: Advances and Challenges in a Changing Climate. Front. Plant Sci. 2019, 10, 103. [Google Scholar] [CrossRef] [Green Version]
- Gifford, R.M. Plant respiration in productivity models: Conceptualisation, representation and issues for global terrestrial carbon-cycle research. Funct. Plant Biol. 2003, 30, 171–186. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Su, B.; Currie, W.; Dukes, J.; Finzi, A.; Hartwig, U.; Hungate, B.; McMurtrie, R.; Oren, R.; Parton, W.; et al. Progressive Nitrogen Limitation of Ecosystem Responses to Rising Atmospheric Carbon Dioxide. BioScience 2004, 54, 731–739. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.W.; Yu, D.P.; Dai, L.M.; Zhou, L.; Zhou, W.M.; Qi, G.; Qi, L.; Ye, Y.J. Research progress in water use efficiency of plants under global climate change. Chin. J. Appl. Ecol. 2010, 21, 3255–3265. [Google Scholar]
- Bloom, A.J.; Chapin, F.S.; Mooney, H.A. Resource Limitation in Plants-An Economic Analogy. Ann. Rev. Ecol. System. 1985, 16, 363–392. [Google Scholar] [CrossRef]
- Limousin, J.M.; Yepez, E.A.; McDowell, N.G.; Pockman, W.T.; Tjoelker, M. Convergence in resource use efficiency across trees with differing hydraulic strategies in response to ecosystem precipitation manipulation. Funct. Ecol. 2015, 29, 1125–1136. [Google Scholar] [CrossRef]
- Szatniewska, J.; Zavadilova, I.; Nezval, O.; Krejza, J.; Petrik, P.; Cater, M.; Stojanovic, M. Species-specific growth and transpiration response to changing environmental conditions in floodplain forest. For. Ecol. Manag. 2022, 516, 120248. [Google Scholar] [CrossRef]
- Gong, X.Y.; Chen, Q.; Lin, S.; Brueck, H.; Dittert, K.; Taube, F.; Schnyder, H. Tradeoffs between nitrogen- and water-use efficiency in dominant species of the semiarid steppe of Inner Mongolia. Plant Soil 2011, 340, 227–238. [Google Scholar] [CrossRef]
- Loreau, M. Biodiversity and ecosystem functioning: Recent theoretical advances. Oikos 2000, 91, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Loreau, M.; Hector, A. Partitioning selection and complementarity in biodiversity experiments. Nature 2001, 412, 72–76. [Google Scholar] [CrossRef]
- Li, X.H.; Yan, H.J.; Wei, T.Z.; Zhou, W.J.; Jia, X.; Zha, T.S. Relative changes of resource use efficiencies and their responses to environmental factors in Artemisia ordosica during growing season. Chin. J. Plant Ecol. 2019, 43, 889–898. [Google Scholar] [CrossRef]
- Zhou, W.J.; Zha, T.S.; Jia, X.; Tian, Y.; Wei, T.Z.; Jin, C. Dynamics of water use efficiency of Artemisia ordosica leaf in growing season in response to environmental factors in Yanchi, Ningxia of northwestern China. J. Beijing For. Univ. 2020, 42, 98–105. [Google Scholar]
- Jiang, Y.; Jin, C.; Jiang, X.Y.; Li, X.H.; Wei, N.N.; Gao, S.J.; Liu, P.; Jia, X.; Zha, T.S. Relative changes and biophysical controls of leaf resource use efficiencies in Artemisia ordosica. Acta Ecol. Sin. 2022, 42, 15. [Google Scholar]
- Hu, L.D.; Zhou, H.J.; Huang, Y.Z.; Yao, X.Y.; Ye, S.M.; Yu, S.F. A study on plant species diversity and soil carbon and nitrogen in different Cunninghamia lanceolata stand types. Ecol. Environ. Sci. 2022, 31, 451–459. [Google Scholar]
- Cao, S. Excessive reliance on afforestation in China’s arid and semi-arid regions: Lessons in ecological restoration. Earth Sci. Rev. 2011, 104, 240–245. [Google Scholar] [CrossRef]
- Ye, Z.P.; Yu, Q.; Kang, H.J. Evaluation of photosynthetic electron flow using simultaneous measurements of gas exchange and chlorophyll fluorescenceunder photorespiratory conditions. Photosynthetica 2012, 50, 472–476. [Google Scholar] [CrossRef]
- Li, X.Q.; Lu, Y.M.; Huang, A.M.; Yuan, R.B.; Li, J.L.; Hu, D.D.; Zhong, Q.L.; Cheng, D.L. Light response model fitting and photosynthetic characteristics of tendifferent fern species in subtropics. Acta Ecol. Sin. 2022, 42, 3333–3344. [Google Scholar]
- Jiang, G.M.; He, W.M. Species-and habitat-variability of photosynthesis transpiration and water use efficiency of different plant species in Maowusu sand area. Chin. J. Plant Ecol. 1999, 41, 1114–1124. [Google Scholar]
- Zhao, H.Y.; Li, Y.L.; Wang, X.Y.; Mao, W.; Zhao, X.Y.; Zhang, T.H. Study on the variation of leaf characters of 52 species of plants in Horqin sandy land. J. Desert Res. 2010, 30, 1292–1298. [Google Scholar]
- Li, Y.L.; Cui, J.Y.; Su, Y.Z. Specific leaf area and leaf dry matter content of some plants in different dune habitats. Acta Ecol. Sin. 2005, 25, 304–311. [Google Scholar]
- Lv, J.Z.; Miao, Y.M.; Zhang, H.F.; Bi, R.C. Comparisons of leaf traits among differernt functional types of plant from Huoshan mountain in the Shanxi province. J. Wuhan Botan. Res. 2010, 28, 460–465. [Google Scholar]
- Zhang, Z.G.; Wei, H.X. Specific Leaf Area and Leaf Dry Matter Content of Main Plant Species in Red Lotus Wetland of Weishan Lake. Acta Agric. Jiangxi 2021, 33, 63–69. [Google Scholar]
- Yang, Q.; Chen, Z.F.; Zhou, J.J.; Lai, S.B.; Jian, C.X.; Wang, Z.; Xu, B.C. Responses of leaf functional traits of dominant plant species in grassland communities to nitrogen and phosphorus addition in loess hilly—Gully region. Chin. J. Appl. Ecol. 2019, 30, 3697–3706. [Google Scholar]
- Liu, X.X.; Li, J.Q. Study on the functional traits of plant leaf in Beijing yeyahu wetland. J. Anhui Agric. 2008, 36, 8046–8409, 8428. [Google Scholar]
- Li, Y.H.; Luo, T.X.; Lu, Q.; Tian, X.Y.; Wu, B.; Yang, H.H. Comparisons of leaf traits among 17major plant species in Shazhuyu Sand Control Experimental Station of Qinghai Province. Acta Ecol. Sin. 2005, 25, 994–999. [Google Scholar]
- Mahmud, K.; Medlyn, B.E.; Duursma, R.A.; Company, C.; DeKauwe, M.G. Inferring the effects of sink strength on plant carbon balance processes from experimental measurements. Biogeosciences 2018, 15, 4003–4018. [Google Scholar] [CrossRef] [Green Version]
- Vona, V.; Di Martino Rigano, V.; Andreoli, C.; Lobosco, O.; Caiazzo, M.; Martello, A.; Carfagna, S.; Salbitani, G.; Rigano, C. Comparative analysis of photosynthetic and respiratory parameters in the psychrophilic unicellular green alga Koliella antarctica, cultured in indoor and outdoor photo-bioreactors. Physiol. Mol. Biol. Plants 2018, 24, 1139–1146. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Peng, Y.D.; Yu, L.Y.; Chen, X.Y.; Pan, C.L.; Lan, Z.Z.; Huang, X.Y. Study on Photosynthetic Characteristics of several Curcuma plants. Jiangsu Agric. Sci. 2021, 49, 143–149. [Google Scholar]
- Niu, S.L.; Jiang, G.M.; Gao, L.M.; Li, Y.G.; Liu, M.Z. Comparison of gas exchange traits of differernt plant species in hunshandak sand area. Chin. J. Plant Ecol. 2003, 27, 318–324. [Google Scholar]
- Qiu, J.; Tan, D.Y.; Fan, D.Y. Characteristics of photosynthesis and biomiss allocation of spring ephemerals in the junggar desert. Chin. J. Plant Ecol. 2007, 31, 883–891. [Google Scholar]
- Li, X.R.; Liu, Q.J.; Cai, Z.; Ma, Z.Q. Specific leaf area and leaf area index of conifer plantations in Qianyanzhou station of subtropical China. Chin. J. Plant Ecol. 2007, 31, 93–101. [Google Scholar]
- Chen, Z.H.; Zha, T.S.; Jia, X.; Wu, Y.; Wu, B.; Zhang, Y.; Guo, J.; Qin, S.; Chen, G.; Peltola, H. Leaf nitrogen is closely coupled to phenophases in a desert shrub ecosystem in China. J. Arid Environ. 2015, 122, 124–131. [Google Scholar] [CrossRef]
- Takashima, T.; Hikosaka, K. Photosynthesis or persistence: Nitrogen allocation in leaves of evergreen and deciduous Quercus species. Plant Cell Environ. 2004, 27, 1047–1054. [Google Scholar] [CrossRef]
- Yu, H.Y.; Chen, Y.T.; Xu, Z.Z.; Zhou, G.S. Analysis of relationships among leaf functional traits and economics spectrum of plant species in the desert steppe of Nei Mongol. Chin. J. Plant Ecol. 2014, 38, 1029–1040. [Google Scholar]
- Zhang, X.; He, X.; Gao, J.; Wang, L. Latitudinal and climate effects on key plant traits in Chinese forest ecosystems. Glob. Ecol. Conserv. 2019, 17, e00527. [Google Scholar] [CrossRef]
- Kattge, J.; Diaz, S.; Lavorel, S.; Prentice, C.; Leadley, P.; Bonisch, G.; Garnier, E.; Westoby, M.; Reich, P.B.; Wright, I.J.; et al. TRY—A global database of plant traits. Glob. Chang. Biol. 2011, 17, 2905–2935. [Google Scholar] [CrossRef]
- Petrik, P.; Peter-Petrik, A.; Konôpková, A.; Fleischer, P.; Stojnic, S.; Zavadilova1, I.; Kurjak, D. Seasonality of PSII thermostability and water use efciency of in situ mountainous Norway spruce (Picea abies). J. For. Res. 2022, 1–12. [Google Scholar]
- Guerrieri, R.; Belmecheri, S.; Ollinger, S.V.; Asbjornsen, H.; Jennings, K.; Xiao, J.; Stocker, B.D.; Martin, M.; Hollinger, D.Y.; Bracho-Garrillo, R.; et al. Disentangling the role of photosynthesis and stomatalconductance on rising forest water-use efficiency. Proc. Natl. Acad. Sci. USA 2019, 116, 16909–16914. [Google Scholar] [CrossRef]
- Ehleringer, J.R.; Cooper, T.A. Correlations between carbon isotope ratio and microhabitat in desert plants. Oecologia 1988, 76, 562–566. [Google Scholar] [CrossRef]
- Fischer, R.A.; Turner, N.C. Plant productivity in the arid and semiarid zones. Annu. Rev. Plant Physiol. 1978, 29, 277–317. [Google Scholar] [CrossRef]
- Miller, J.M.; Williams, R.J.; Farquhar, G.D. Carbon isotope discrimination by a sequence of Eucalyptus species along a subcontinental rainfall gradient in Australia. Funct. Ecol. 2001, 15, 222–232. [Google Scholar] [CrossRef]
- Quan, X.Q.; Wang, C.K. Adaptation of leaf carbon use efficiency of Larix gmelinii to environmental change. Acta Ecol. Sin. 2016, 36, 3381–3390. [Google Scholar]
- Tilman, D.; Knops, J.; Wedin, D.; Reich, P.; Ritchie, M.; Siemann, E. The Influence of Functional Diversity and Composition on Ecosystem Processes. Am. Assoc. Adv. Sci. 1997, 277, 1300–1302. [Google Scholar] [CrossRef] [Green Version]
- Mack, M.C.; D’Antonio, C.M. Impacts of biological invasions on disturbance regimes. Trends Ecol. Evol. 1998, 13, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Lauenroth, W.K.; Dodd, J.L. The Effects of Water- and Nitrogen-Induced Stresses on Plant Community Structure in a Semiarid Grassland. Oecologia 1978, 36, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Monica, L.P.; Roger, L.S.; Catherine, A.Z.; Richard, E.E.; Tony, J.S.; Borkowski, J.J. Plant Functional Group Diversity as a Mechanism for Invasion Resistance. Restor. Ecol. 2005, 13, 448–459. [Google Scholar]
- Zheng, S.X.; ShangGuan, Z.P. Photosynthetic characteristics and their relationships with leaf nitrogen content and leaf mass per area in different plant functionaltypes. Acta Ecol. Sin. 2007, 27, 171–181. [Google Scholar]
- Onoda, Y.; Hikosaka, K.; Hirose, T. Allocation of Nitrogen to Cell Walls Decreases Photosynthetic Nitrogen-Use Efficiency. Funct. Ecol. 2004, 18, 419–425. [Google Scholar] [CrossRef]
- Huang, W.J.; Li, Z.J.; Yang, Z.P.; Bai, G.Z. The structural traits of populus euphratica heterom orphic leaves and their correlations. Acta Ecol. Sin. 2010, 30, 4636–4642. [Google Scholar]
- Shahid, N.; Lindsey, J.T.; Sharon, P.L.; John, H.L.; Woodfin, R.M. Declining biodiversity can alter the performance of ecosystems. Nature 1994, 368, 734–737. [Google Scholar]
- Anderson, J.E.; Inouye, R.S. Landscape-scale changes in plant species abundance and biodiversity of a sagebrush steppe over 45 years. Ecol. Monogr. 2001, 71, 531–556. [Google Scholar] [CrossRef]
- Patterson, T.B.; Guy, R.D.; Dang, Q.L. Whole-plant nitrogen- and water-relations traits, and their associated trade-offs, in adjacent muskeg and upland boreal spruce species. Oecologia 1997, 110, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Bai, Y.; Zhang, L.; Han, X. Comparing physiological responses of two dominant grass species to nitrogen addition in Xilin River Basin of China. Environ. Exp. Bot. 2005, 53, 65–75. [Google Scholar] [CrossRef]
- Wang, G.H. Can the restoration of natural vegetation be accelerated on the Chinese Loess Plateau? A study of the response of the leaf carbon isotope ratio of dominant species to changing soil carbon and nitrogen levels. Ecol. Res. 2005, 21, 188–196. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Li, L.H. Soil water status influences plant nitrogen use: A case study. Plant Soil 2007, 301, 303–313. [Google Scholar] [CrossRef]
- Zhan, X.Y.; Yu, G.R.; Sheng, W.P.; Fang, H.J. Foliar water use efficiency and nitrogen use efficiency of dominant plant species in main forests along the North-South Transect of East China. Chin. J. Appl. Ecol. 2012, 23, 587–594. [Google Scholar]
- Tarvainen, L.; Rantfors, M.; Wallin, G. Seasonal and within-canopy variation in shoot-scale resource-use efficiency trade-offs in a Norway spruce stand. Plant Cell Environ. 2015, 38, 2487–2496. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Life Form | Species | Family | Height H/m | Breast Diameter B/cm | Stand Age S/years | Central Latitude and Longitude | Elevation/m | |
|---|---|---|---|---|---|---|---|---|
| Tree | Deciduous trees | Rhus typhina | Anacardiaceae | 5.16 | 12.01 | 8 | 116°2′5″ E, 39°46′29″ N | 95 |
| Deciduous trees | Malus spectabilis | Rosaceae | 3.54 | 10.54 | 8 | 116°2′10″ E, 39°46′19″ N | 100 | |
| Deciduous trees | Prunus davidiana | Rosaceae | 5.94 | 8.76 | 8 | 116°2′25″ E, 39°46′8″ N | 110 | |
| Deciduous trees | Ulmus pumila ‘Jinye’ | Ulmaceae | 4.63 | 11.81 | 8 | 116°2′27″ E, 39°46′46″ N | 100 | |
| Deciduous trees | Fraxinus chinensis | Oleaceae | 5.37 | 9.16 | 8 | 116°2′1″ E, 39°46′26″ N | 100 | |
| Nitrogen fixing plant | Styphnolobium japonicum | Leguminosae | 6.08 | 7.52 | 8 | 116°2′1″ E, 39°46′31″ N | 100 | |
| Deciduous trees | Crataegus pinnatifida | Rosaceae | 2.5 | 7.86 | 8 | 116°1′41″ E, 39°46′35″ N | 100 | |
| Deciduous trees | Prunus cerasifera ‘Atropurpurea’ | Rosaceae | 4.68 | 11.28 | 8 | 116°2′2″ E, 39°46′41″ N | 90 | |
| Nitrogen fixing plant | Robinia pseudoacacia | Leguminosae | 6.24 | 9.69 | 8 | 116°1′46″ E, 39°46′26″ N | 110 | |
| Deciduous trees | Populus tomentosa. | Salicaceae | 6.93 | 12.3 | 8 | 116°2′16″ E, 39°46′44″ N | 100 | |
| Deciduous trees | Prunus armeniaca | Rosaceae | 5.52 | 18.25 | 8 | 116°2′10″ E, 39°46′19″ N | 80 | |
| Deciduous trees | Yulania denudata | Magnoliaceae | 3.24 | 25.7 | 8 | 116°2′10″ E, 39°46′19″ N | 95 | |
| Evergreen conifer | Pinus bungeana | Pinaceae | 3.65 | 16.56 | 8 | 116°2′13″ E, 39°46′18″ N | 110 | |
| Evergreen conifer | Pinus tabuliformis | Pinaceae | 3.07 | 8.07 | 8 | 116°2′13″ E, 39°46′18″ N | 110 | |
| Evergreen conifer | Platycladus orientalis | Cupressaceae | 4.62 | 9.55 | 8 | 116°2′26″ E, 39°46′40″ N | 95 | |
| Shrub | Nitrogen fixing plant | Amorpha fruticosa | Leguminosae | 2.04 | 4.25 | 8 | 116°2′26″ E, 39°46′44″ N | 95 |
| Deciduous shrub | Cotinus coggygria | Anacardiaceae | 3.25 | 7.26 | 8 | 116°2′27″ E, 39°46′44″ N | 95 | |
| Deciduous shrub | Forsythia suspensa | Oleaceae | 0.74 | 2.55 | 8 | 116°2′13″ E, 39°46′18″ N | 110 | |
| Deciduous shrub | Euonymus japonicus ‘Aureo-marginatus’ | Celastraceae | 0.65 | 2.12 | 8 | 116°2′10″ E, 39°46′19″ N | 100 | |
| Species | Pnmax (μmol·m2·s−1) | SLA (cm2·g−1) | Nmass (g/kg) | WUE (mmol·mol−1) | NUE (μmol·g−1·s−1) | CUE (%) |
|---|---|---|---|---|---|---|
| Rhus typhina | 26.09 ± 0.41 | 146.6 ± 29.58 | 35.88 ± 0.23 | 4.64 ± 0.36 | 10.66 ± 0.17 | 88.6 ± 0.35 |
| Malus spectabilis | 22.6 ± 3.07 | 129.5 ± 25.98 | 21.06 ± 0.46 | 3.58 ± 0.07 | 13.9 ± 1.13 | 84.2 ± 0.67 |
| Prunus davidiana | 20.64 ± 0.62 | 135.3 ± 32.63 | 24.12 ± 0.25 | 3.63 ± 0.2 | 11.58 ± 1.12 | 86.8 ± 1.96 |
| Ulmus pumila ‘Jinye’ | 21.95 ± 2.01 | 202.2 ± 64.29 | 32.64 ± 0.35 | 4.01 ± 0.34 | 13.6 ± 1.25 | 85.2 ± 0.45 |
| Fraxinus chinensis | 18.31 ± 0.59 | 143.4 ± 33.06 | 21.16 ± 0.27 | 2.78 ± 0.29 | 9.66 ± 0.31 | 81.1 ± 2.61 |
| Styphnolobium japonicum | 15.81 ± 0.3 | 149.5 ± 48.43 | 22.9 ± 0.25 | 5.14 ± 0.64 | 9.9 ± 0.16 | 86.2 ± 3.99 |
| Crataegus pinnatifida | 17.34 ± 1.14 | 94.2 ± 11.15 | 18.33 ± 0.16 | 3.52 ± 0.38 | 8.91 ± 1.17 | 87.6 ± 1.08 |
| Prunus cerasifera ‘Atropurpurea’ | 15.4 ± 0.81 | 237.6 ± 125.38 | 21.56 ± 0.14 | 2.91 ± 0.36 | 16.97 ± 3.82 | 85.5 ± 1.38 |
| Robinia pseudoacacia | 16.02 ± 2.09 | 149.5 ± 31.28 | 20.34 ± 0.22 | 3.4 ± 0.68 | 11.77 ± 2.27 | 84.9 ± 1.94 |
| Populus tomentosa | 12.98 ± 0.62 | 123.3 ± 26.24 | 22.13 ± 0.07 | 3.8 ± 0.71 | 7.23 ± 1.32 | 80.7 ± 1.05 |
| Prunus armeniaca | 11.32 ± 1.21 | 162.3 ± 15.72 | 22.17 ± 0.23 | 4.89 ± 1.07 | 8.29 ± 0.88 | 85.4 ± 1.27 |
| Yulania denudata | 6.72 ± 0.14 | 150 ± 22.58 | 19.02 ± 0.2 | 4.95 ± 1.46 | 5.3 ± 2.08 | 84.4 ± 0.36 |
| Pinus bungeana | 20.87 ± 2.3 | 39 ± 4.04 | 12.58 ± 0.35 | 7.84 ± 2.4 | 6.2 ± 0.69 | 85.8 ± 2.09 |
| Pinus tabuliformis | 19.26 ± 0.46 | 41 ± 8.05 | 14.29 ± 2.95 | 9.11 ± 1.35 | 5.89 ± 0.14 | 94.8 ± 1.67 |
| Platycladus orientalis | 10.6 ± 0.76 | 79 ± 9.61 | 11.11 ± 1.38 | 5.38 ± 0.94 | 7.14 ± 0.51 | 71.9 ± 2.45 |
| Amorpha fruticosa | 28.92 ± 1.6 | 217.4 ± 22.46 | 38.8 ± 1.32 | 3.21 ± 0.62 | 12.9 ± 0.72 | 83.6 ± 2.11 |
| Cotinus coggygria | 15.84 ± 2.79 | 125.4 ± 15.28 | 21.8 ± 1.0 | 6 ± 1.1 | 9.34 ± 1.65 | 87.2 ± 1.54 |
| Forsythia suspensa | 23.67 ± 0.06 | 135.9 ± 21.06 | 15.8 ± 2.5 | 3.2 ± 0.22 | 15.58 ± 0.04 | 82.4 ± 1.87 |
| Euonymus japonicus ‘Aureo-marginatus’ | 10.6 ± 3.79 | 122 ± 15.28 | 25.8 ± 3.25 | 2.54 ± 1.64 | 6.86 ± 1.79 | 76.5 ± 4.56 |
| ANOVA | ||||
|---|---|---|---|---|
| Variable | Sum of Squares | df | Mean Square | F |
| WUE | 136.83 | 18 | 7.6 | 8.7 *** |
| CUE | 1125 | 18 | 62.5 | 14.6 *** |
| NUE | 530.13 | 18 | 29.45 | 15.26 *** |
| Pnmax | 1657.38 | 18 | 92.08 | 20.79 *** |
| SLA | 288,360.43 | 18 | 16,020.02 | 5.92 *** |
| Nmass | 5637.81 | 18 | 313.21 | 55.32 *** |
| Traits | Principal Component 1 | Principal Component 2 |
|---|---|---|
| Pnmax | 0.35 | 0.49 |
| SLA | 0.51 | −0.14 |
| Nmass | 0.46 | 0.1 |
| WUE | −0.39 | 0.49 |
| CUE | 0.02 | 0.69 |
| NUE | 0.49 | 0.07 |
| Variance ratio | 47.16% | 25.9% |
| Cumulative variance ratio | 47.16% | 73.06% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, G.; Tian, Y.; Liu, P.; Jia, X.; Zha, T. Leaf Traits and Resource Use Efficiencies of 19 Woody Plant Species in a Plantation in Fangshan, Beijing, China. Forests 2023, 14, 63. https://doi.org/10.3390/f14010063
Zhong G, Tian Y, Liu P, Jia X, Zha T. Leaf Traits and Resource Use Efficiencies of 19 Woody Plant Species in a Plantation in Fangshan, Beijing, China. Forests. 2023; 14(1):63. https://doi.org/10.3390/f14010063
Chicago/Turabian StyleZhong, Guowei, Yun Tian, Peng Liu, Xin Jia, and Tianshan Zha. 2023. "Leaf Traits and Resource Use Efficiencies of 19 Woody Plant Species in a Plantation in Fangshan, Beijing, China" Forests 14, no. 1: 63. https://doi.org/10.3390/f14010063
APA StyleZhong, G., Tian, Y., Liu, P., Jia, X., & Zha, T. (2023). Leaf Traits and Resource Use Efficiencies of 19 Woody Plant Species in a Plantation in Fangshan, Beijing, China. Forests, 14(1), 63. https://doi.org/10.3390/f14010063