
The Impact of Long-Term Dry-Season Irrigation on Eucalyptus Tree Height Growth: Insights from Leaf Photosynthesis and Water Conduction
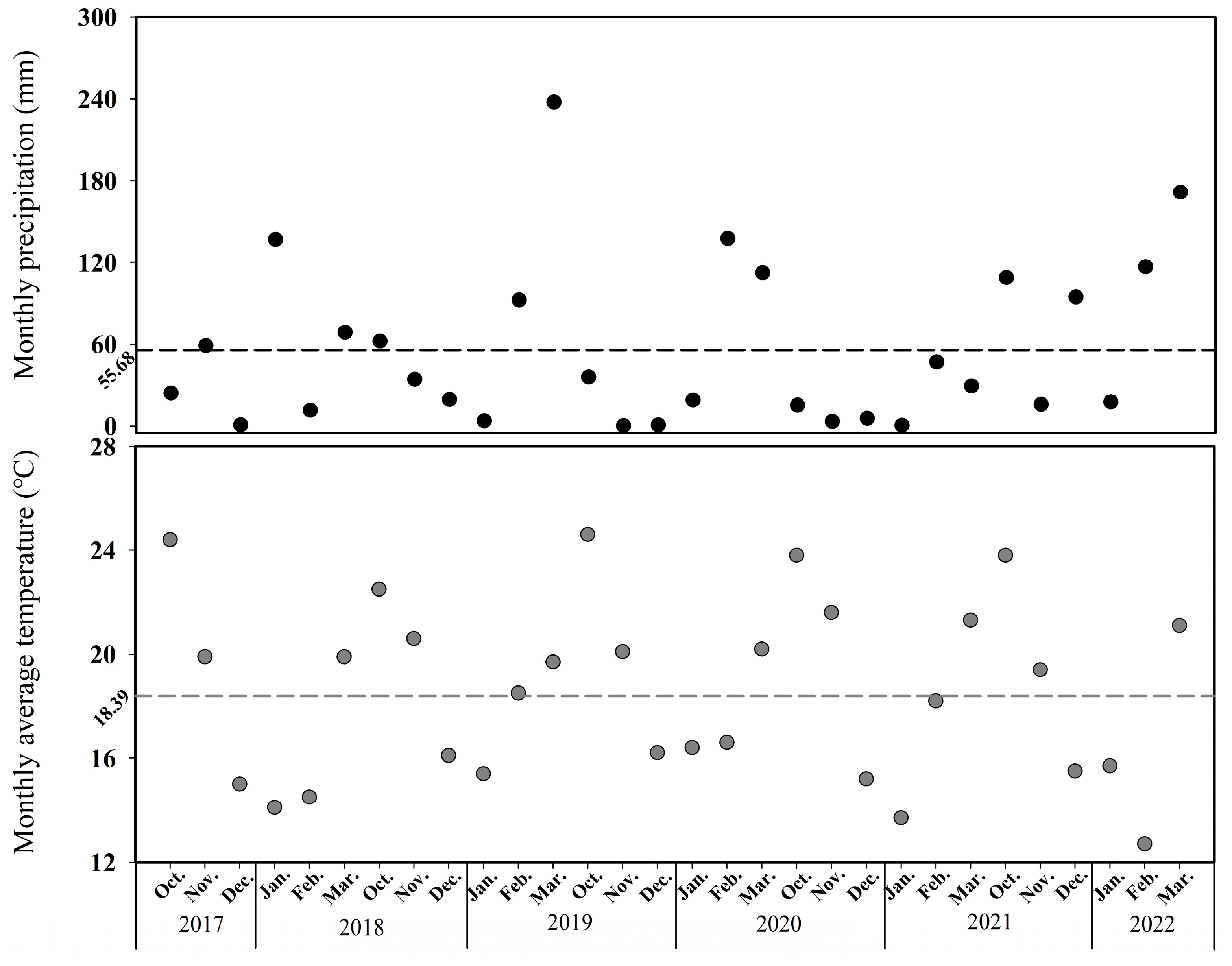{kind=link}
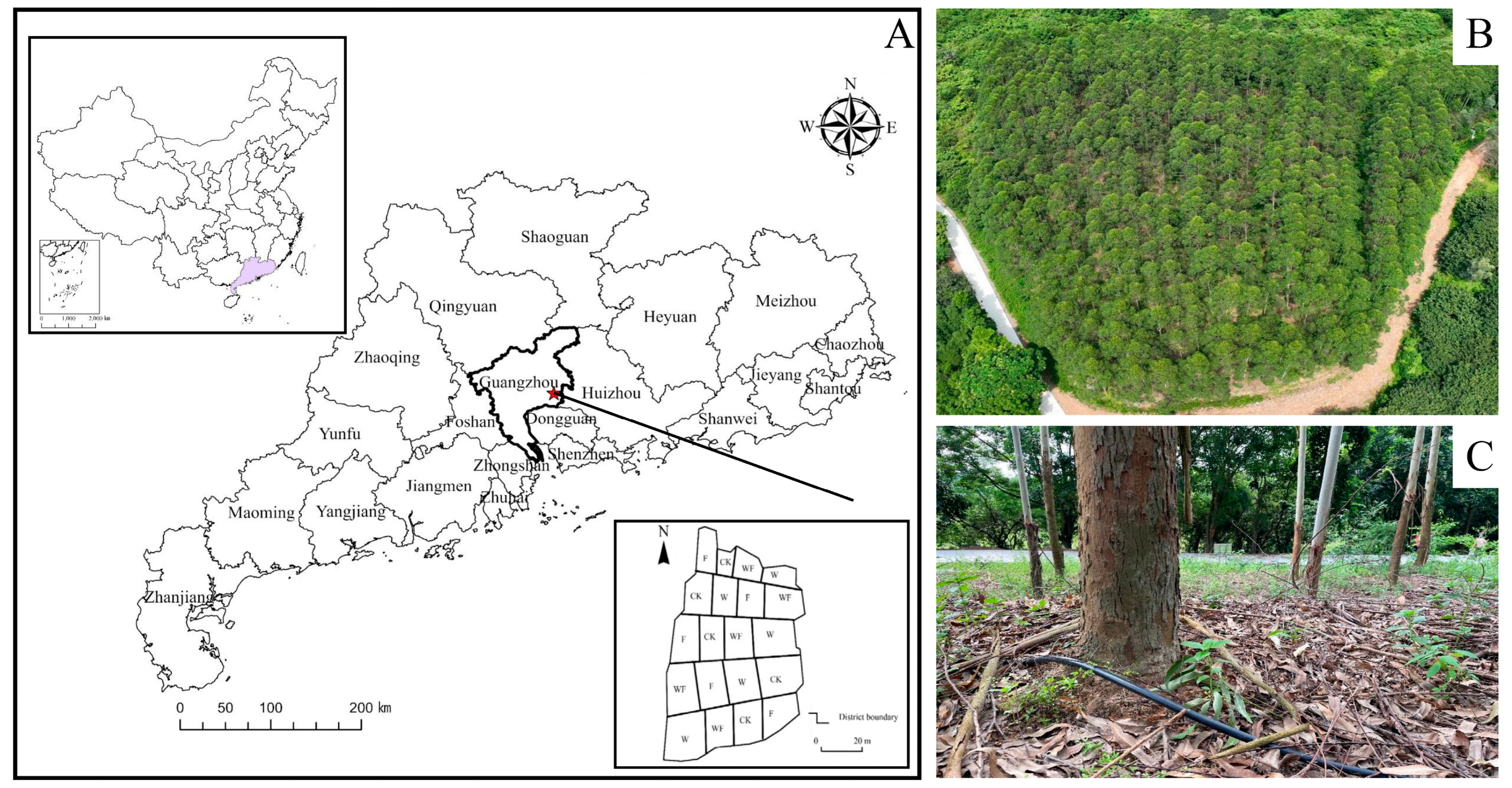{kind=link}
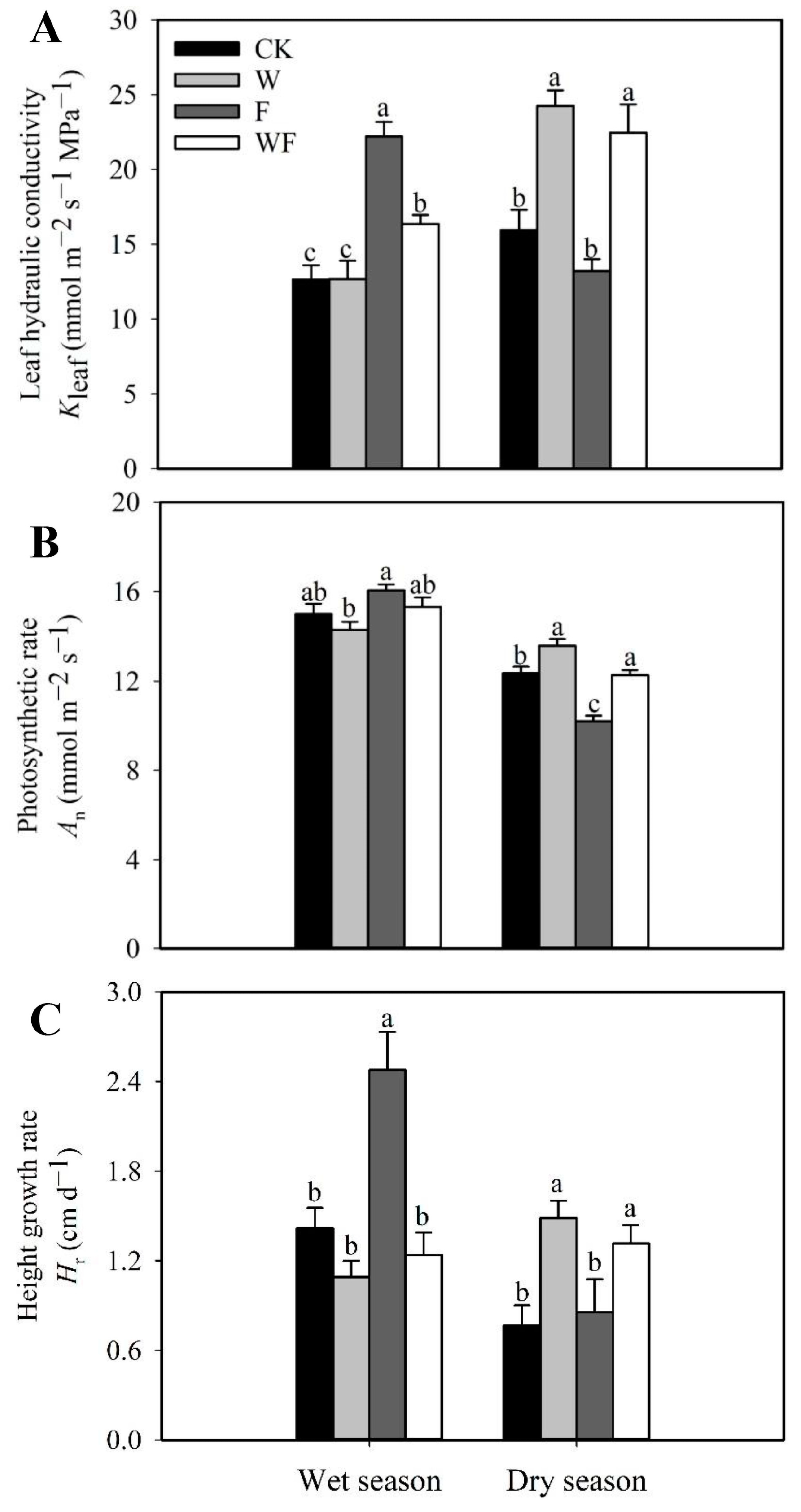{kind=link}
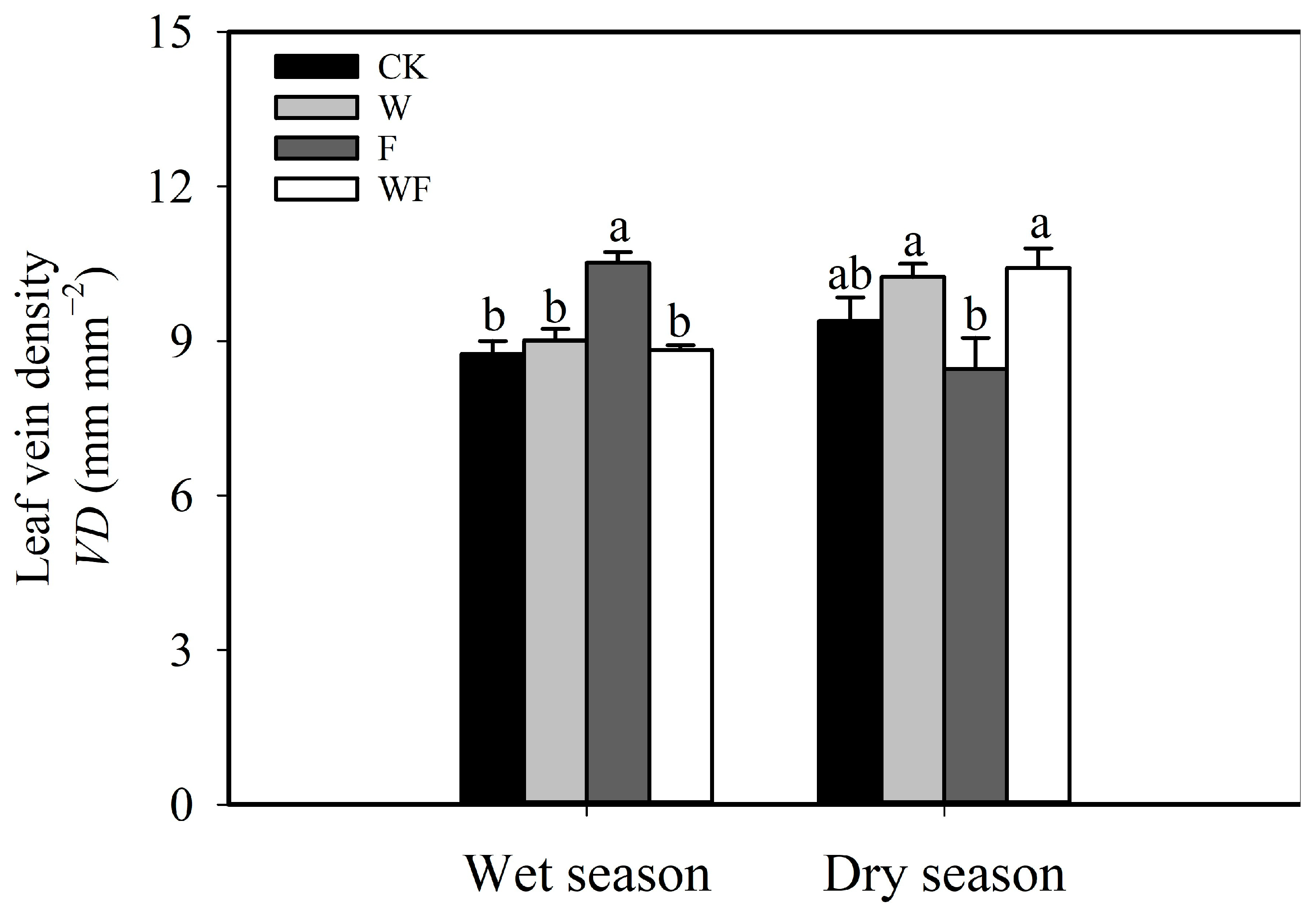{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Plant Material
2.2. Experimental Design
2.3. Measurement of Height Growth Rate
2.4. Measurement of Leaf Functional Traits
2.5. Measurement of Leaf Structural Traits
2.6. Statistical Analysis
3. Results
3.1. Response of Functional Traits to Five-Year Dry-Season Irrigation
3.2. Response of Structural Traits to Five-Year Dry-Season Irrigation
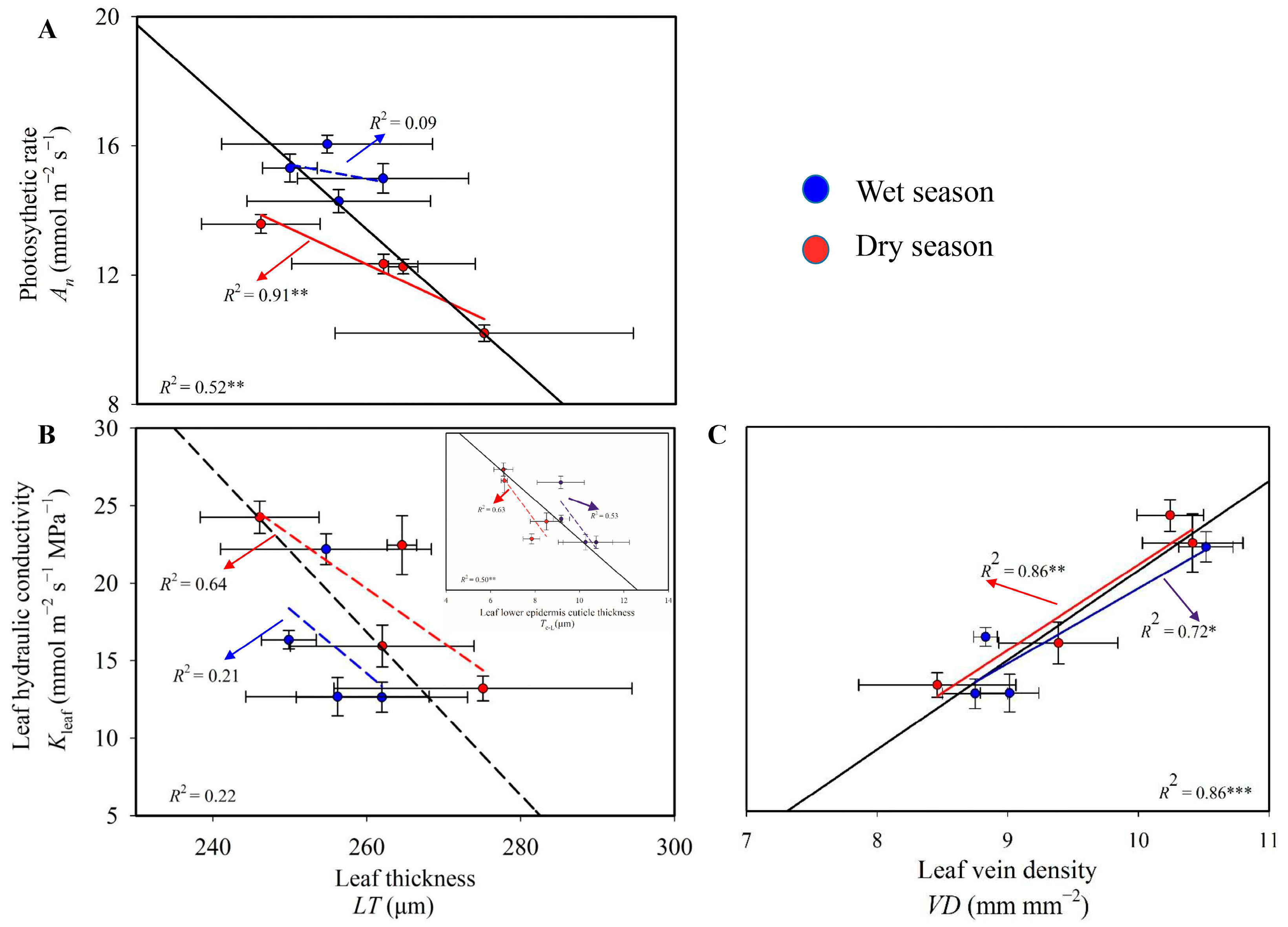
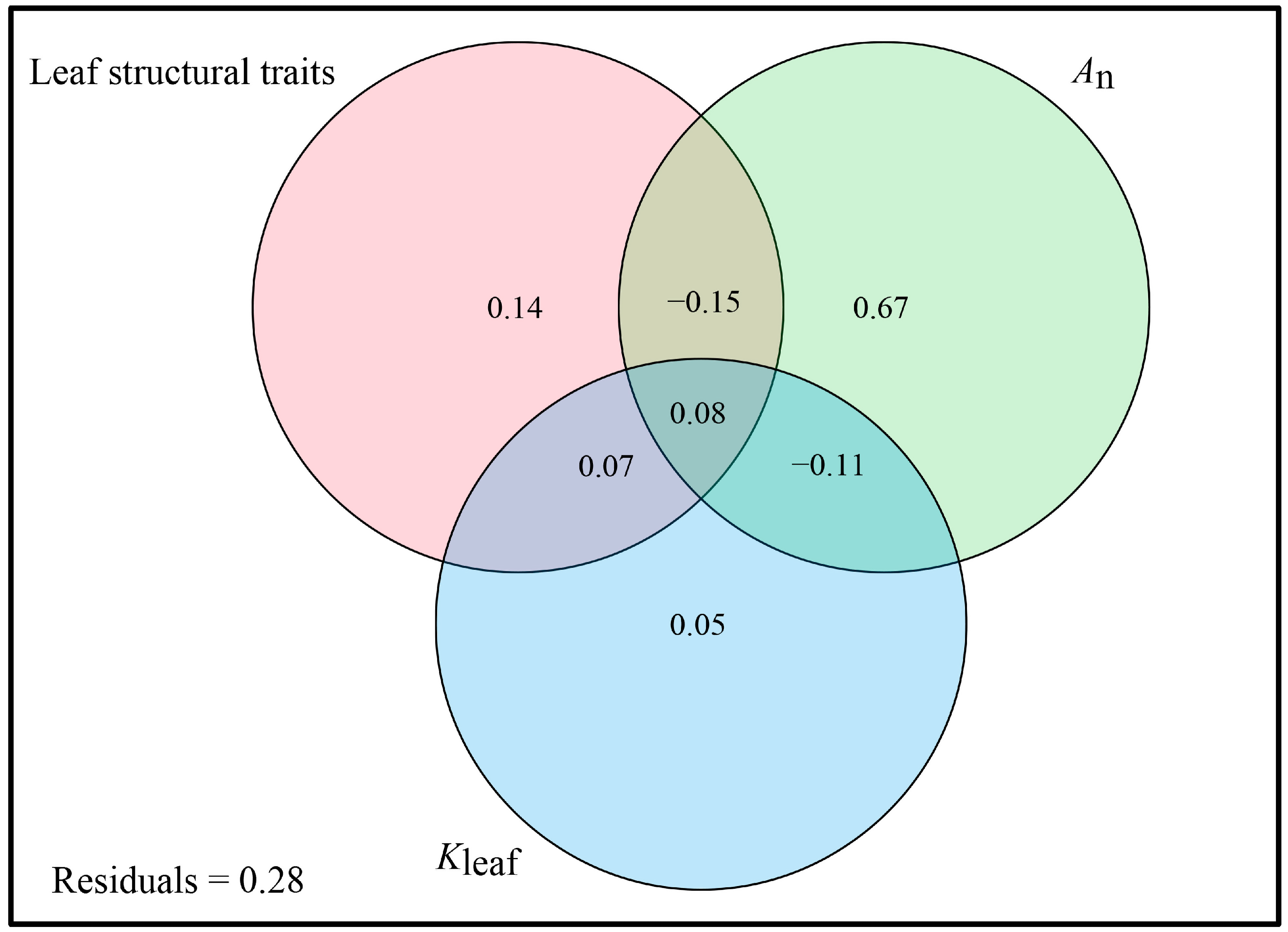
3.3. Associations between Functional Traits and Structural Traits
4. Discussion
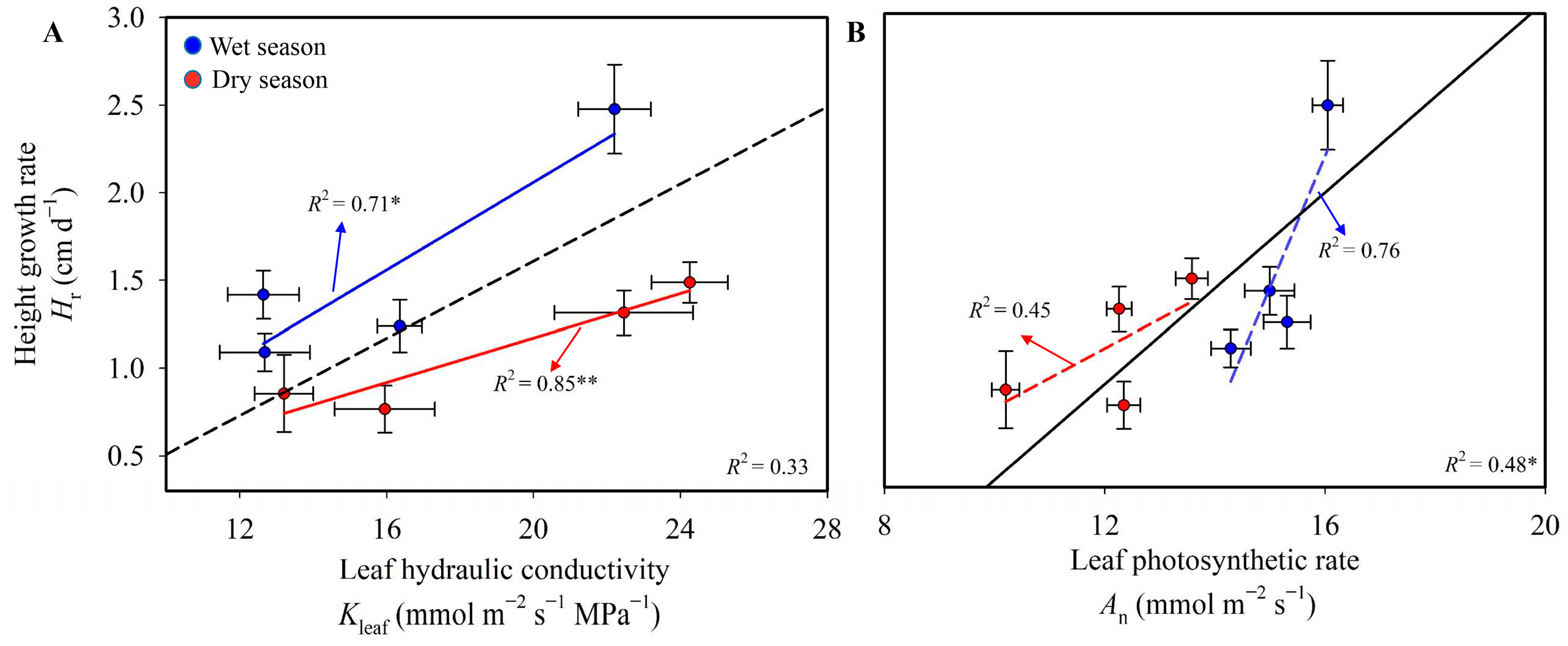
4.1. Effect of Long-Term Dry-Season Irrigation on Plant Height Growth
4.2. Effect of Leaf Structural and Functional Traits on Plant Height Growth
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, X.; Zhu, H.; Wen, Y.; Goodale, U.M.; Li, X.; You, Y.; Ye, D.; Liang, H. Effects of understory management on trade-offs and synergies between biomass carbon stock, plant diversity and timber production in eucalyptus plantations. For. Ecol. Manag. 2018, 410, 164–173. [Google Scholar] [CrossRef]
- Hua, L.; He, P.; Goldstein, G.; Liu, H.; Yin, D.; Zhu, S.; Ye, Q.; Riederer, M. Linking vein properties to leaf biomechanics across 58 woody species from a subtropical forest. Plant Biol. 2019, 22, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Hua, L.; Yu, F.; Qiu, Q.; He, Q.; Su, Y.; Liu, X.; Li, J. Relationships between diurnal and seasonal variation of photosynthetic characteristics of Eucalyptus plantation and environmental factors under dry-season irrigation with fertilization. Agric. Water Manag. 2021, 248, 106737. [Google Scholar] [CrossRef]
- Hua, L.; Yu, F.; Qiu, Q.; He, Q.; Su, Y.; Liu, X.; Li, J. Dry-season irrigation further promotes the growth of Eucalyptus urophylla × E. grandis plantations under the conventional fertilization. New For. 2023, 54, 1085–1102. [Google Scholar] [CrossRef]
- Yu, F.; Truong, T.V.; He, Q.; Hua, L.; Su, Y.; Li, J. Dry Season Irrigation Promotes Leaf Growth in Eucalyptus urophylla × E. grandis under Fertilization. Forests 2019, 10, 67. [Google Scholar] [CrossRef]
- Whitehead, D.; Beadle, C.L. Physiological regulation of productivity and water use in Eucalyptus: A review. For. Ecol. Manag. 2004, 193, 113–140. [Google Scholar] [CrossRef]
- Kenzo, T.; Ichie, T.; Watanabe, Y.; Yoneda, R.; Ninomiya, I.; Koike, T. Changes in photosynthesis and leaf characteristics with tree height in five dipterocarp species in a tropical rain forest. Tree Physiol. 2006, 26, 865–873. [Google Scholar] [CrossRef]
- Kitao, M.; Harayama, H.; Yazaki, K.; Tobita, H.; Agathokleous, E.; Furuya, N.; Hashimoto, T. Photosynthetic and Growth Responses in a Pioneer Tree (Japanese White Birch) and Competitive Perennial Weeds (Eupatorium sp.) Grown under Different Regimes with Limited Water Supply to Waterlogging. Front. Plant Sci. 2022, 13, 835068. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef]
- Zhang, A.; Liu, M.; Gu, W.; Chen, Z.; Gu, Y.; Pei, L.; Tian, R. Effect of drought on photosynthesis, total antioxidant capacity, bioactive component accumulation, and the transcriptome of Atractylodes lancea. BMC Plant Biol. 2021, 21, 293. [Google Scholar] [CrossRef]
- Sperry, J.S.; Meinzer, F.C.; McCulloh, K.A. Safety and efficiency conflicts in hydraulic architecture: Scaling from tissues to trees. Plant Cell Environ. 2008, 31, 632–645. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Chen, D.; Zhou, Z.; Ye, Q.; Wu, J. Canopy nitrogen addition enhance the photosynthetic rate of canopy species by improving leaf hydraulic conductivity in a subtropical forest. Front. Plant Sci. 2022, 13, 942851. [Google Scholar] [CrossRef] [PubMed]
- Levionnois, S.; Salmon, C.; Alméras, T.; Clair, B.; Ziegler, C.; Coste, S.; Stahl, C.; González-Melo, A.; Heinz, C.; Heuret, P. Anatomies, vascular architectures, and mechanics underlying the leaf size-stem size spectrum in 42 Neotropical tree species. J. Exp. Bot. 2021, 72, 7957–7969. [Google Scholar] [CrossRef] [PubMed]
- Pereira, J.S.; Chaves, M.M.; Fonseca, F.; Araújo, M.C.; Torres, F. Photosynthetic capacity of leaves of Eucalyptus globulus (Labill.) growing in the field with different nutrient and water supplies. Tree Physiol. 1992, 11, 381–389. [Google Scholar]
- Câmara, A.P.; Vidaurre, G.B.; Oliveira, J.C.L.; de Toledo Picoli, E.A.; Almeida, M.N.F.; Roque, R.M.; Tomazello Filho, M.; Souza, H.J.P.; Oliveira, T.R.; Campoe, O.C. Changes in hydraulic architecture across a water availability gradient for two contrasting commercial Eucalyptus clones. For. Ecol. Manag. 2020, 474, 118380. [Google Scholar] [CrossRef]
- Yin, Q.; Wang, L.; Lei, M.; Dang, H.; Quan, J.; Tian, T.; Chai, Y.; Yue, M. The relationships between leaf economics and hydraulic traits of woody plants depend on water availability. Sci. Total Environ. 2018, 621, 245–252. [Google Scholar] [CrossRef]
- Sack, L.; Frole, K. Leaf Structural Diversity is Related to Hydraulic Capacity in Tropical Rain Forest Trees. Ecology 2006, 87, 483–491. [Google Scholar] [CrossRef]
- Poorter, L.; Bongers, F. Leaf Traits are Good Predictors of Plant Performance across 53 Rain Forest Species. Ecology 2006, 87, 1733–1743. [Google Scholar] [CrossRef]
- Brodribb, T.J.; Feild, T.S.; Jordan, G.J. Leaf Maximum Photosynthetic Rate and Venation Are Linked by Hydraulics. Plant Physiol. 2007, 144, 1890–1898. [Google Scholar] [CrossRef]
- Noblin, X.; Mahadevan, L.; Coomaraswamy, I.A.; Weitz, D.A.; Holbrook, N.M.; Zwieniecki, M.A. Optimal vein density in artificial and real leaves. Proc. Natl. Acad. Sci. USA 2008, 105, 9140–9144. [Google Scholar] [CrossRef]
- Drake, P.L.; Boer, H.J.; Schymanski, S.J.; Veneklaas, E.J. Two sides to every leaf: Water and CO2 transport in hypostomatous and amphistomatous leaves. New Phytol. 2019, 222, 1179–1187. [Google Scholar] [CrossRef] [PubMed]
- Sack, L.; Scoffoni, C. Leaf venation: Structure, function, development, evolution, ecology and applications in the past, present and future. New Phytol. 2013, 198, 983–1000. [Google Scholar] [CrossRef]
- de Boer, H.J.; Eppinga, M.B.; Wassen, M.J.; Dekker, S.C. A critical transition in leaf evolution facilitated the Cretaceous angiosperm revolution. Nat. Commun. 2012, 3, 1221. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zou, Y.; Dang, Y.; Sakai, T. Spatial distribution and dynamic change monitoring of Eucalyptus plantations in China during 1994–2013. Trees 2021, 36, 405–414. [Google Scholar] [CrossRef]
- Asensio, V.; Domec, J.-C.; Nouvellon, Y.; Laclau, J.-P.; Bouillet, J.-P.; Jordan-Meille, L.; Lavres, J.; Rojas, J.D.; Guillemot, J.; Abreu-Junior, C.H. Potassium fertilization increases hydraulic redistribution and water use efficiency for stemwood production in Eucalyptus grandis plantations. Environ. Exp. Bot. 2020, 176, 104085. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Feller, I.C.; McKee, K.L.; Engelbrecht, B.M.J.; Ball, M.C. The effect of nutrient enrichment on growth, photosynthesis and hydraulic conductance of dwarf mangroves in Panama. Funct. Ecol. 2004, 18, 25–33. [Google Scholar] [CrossRef]
- Queiroz, T.B.; Campoe, O.C.; Montes, C.R.; Alvares, C.A.; Cuartas, M.Z.; Guerrini, I.A. Temperature thresholds for Eucalyptus genotypes growth across tropical and subtropical ranges in South America. For. Ecol. Manag. 2020, 472, 118248. [Google Scholar] [CrossRef]
- Yang, L.; Lin, Y.; Kong, J.; Yu, Y.; He, Q.; Su, Y.; Li, J.; Qiu, Q. Effects of fertilization and dry-season irrigation on the timber production and carbon storage in subtropical Eucalyptus plantations. Ind. Crops Prod. 2023, 192, 116143. [Google Scholar] [CrossRef]
- Brodribb, T.J.; Holbrook, N.M. Stomatal Closure during Leaf Dehydration, Correlation with Other Leaf Physiological Traits. Plant Physiol. 2003, 132, 2166–2173. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Warton, D.I.; Wright, I.J.; Falster, D.S.; Westoby, M. Bivariate line-fitting methods for allometry. Biol. Rev. 2006, 81, 259–291. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’hara, R.; Simpson, G.L.; Solymos, P.; et al. Package ‘vegan’. Community Ecol Package Version 2. Available online: http://mirror.bjtu.edu.cn/cran/web/packages/vegan/vegan.pdf (accessed on 16 February 2016).
- Prescott, C.E.; Brown, S.M. Five-year growth response of western red cedar, western hemlock, and amabilis fir to chemical and organic fertilizers. Can. J. For. Res. 1998, 28, 1328–1334. [Google Scholar] [CrossRef]
- Xu, F.; Chu, C.; Xu, Z. Effects of different fertilizer formulas on the growth of loquat rootstocks and stem lignification. Sci. Rep. 2020, 10, 1033. [Google Scholar] [CrossRef]
- Walters, S.J.; Harris, R.J.; Daws, M.I.; Gillett, M.J.; Richardson, C.G.; Tibbett, M.; Grigg, A.H. The benefits of fertiliser application on tree growth are transient in restored jarrah forest. Trees For. People 2021, 5, 100112. [Google Scholar] [CrossRef]
- Zhu, S.-D.; Song, J.-J.; Li, R.-H.; Ye, Q. Plant hydraulics and photosynthesis of 34 woody species from different successional stages of subtropical forests. Plant Cell Environ. 2013, 36, 879–891. [Google Scholar] [CrossRef] [PubMed]
- Koch, G.W.; Sillett, S.C.; Jennings, G.M.; Davis, S.D. The limits to tree height. Nature 2004, 428, 851–854. [Google Scholar] [CrossRef]
- Johnson, R.J.; Canny, M.J. Phloem translocation of organic compounds: A possible mechanism to assist osmotically-generated pressure flow in tall trees. Water 2013, 4, 112–128. [Google Scholar] [CrossRef]
- McGregor, I.R.; Helcoski, R.; Kunert, N.; Tepley, A.J.; Gonzalez-Akre, E.B.; Herrmann, V.; Zailaa, J.; Stovall, A.E.L.; Bourg, N.A.; McShea, W.J.; et al. Tree height and leaf drought tolerance traits shape growth responses across droughts in a temperate broadleaf forest. New Phytol. 2020, 231, 601–616. [Google Scholar] [CrossRef]
- England, J.R.; Attiwill, P.M. Changes in leaf morphology and anatomy with tree age and height in the broadleaved evergreen species, Eucalyptus regnans F. Muell. Trees 2006, 20, 79–90. [Google Scholar] [CrossRef]
- Sack, L.; Cowan, P.; Jaikumar, N.; Holbrook, N.J.P. The ‘hydrology’of leaves: Co-ordination of structure and function in temperate woody species. Plant Cell Environ. 2003, 26, 1343–1356. [Google Scholar] [CrossRef]
- Sack, L.; Holbrook, N.M. Leaf hydraulics. Annu. Rev. Plant Biol. 2006, 57, 361–381. [Google Scholar] [CrossRef] [PubMed]
- Tyree, M.; Cruiziat, P.; Benis, M.; LoGullo, M.; Salleo, S.J.P. The kinetics of rehydration of detached sunflower leaves from different initial water deficits. Plant Cell Environ. 1981, 4, 309–317. [Google Scholar] [CrossRef]
- Yang, S.; Tyree, M.T. Hydraulic architecture of Acer saccharum and A. rubrum: Comparison of branches to whole trees and the contribution of leaves to hydraulic resistance. J. Exp. Bot. 1994, 45, 179–186. [Google Scholar] [CrossRef]
- Nardini, A.; Salleo, S.J.T. Limitation of stomatal conductance by hydraulic traits: Sensing or preventing xylem cavitation? Trees 2000, 15, 14–24. [Google Scholar] [CrossRef]
- Blackman, C.J.; Brodribb, T.J.; Jordan, G.J. Leaf hydraulic vulnerability influences species’ bioclimatic limits in a diverse group of woody angiosperms. Oecologia 2012, 168, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hao, G.-Y.; Hoffmann, W.A.; Scholz, F.G.; Bucci, S.J.; Meinzer, F.C.; Franco, A.C.; Cao, K.-F.; Goldstein, G.J.O. Stem and leaf hydraulics of congeneric tree species from adjacent tropical savanna and forest ecosystems. Oecologia 2008, 155, 405–415. [Google Scholar] [CrossRef]
- Schneider, J.V.; Habersetzer, J.; Rabenstein, R.; Wesenberg, J.; Wesche, K.; Zizka, G. Water supply and demand remain coordinated during breakdown of the global scaling relationship between leaf size and major vein density. New Phytol. 2017, 214, 473–486. [Google Scholar] [CrossRef]
- Comas, L.H.; Eissenstat, D.M. Linking Fine Root Traits to Maximum Potential Growth Rate among 11 Mature Temperate Tree Species. Funct. Ecol. 2004, 18, 388–397. [Google Scholar] [CrossRef]
- Lihui, M.; Xiaoli, L.; Jie, C.; Youke, W.; Jingui, Y. Effects of Slope Aspect and Rainfall on Belowground Deep Fine Root Traits and Aboveground Tree Height. Front. Plant Sci. 2021, 12, 684468. [Google Scholar] [CrossRef]
- Becker, P.; Meinzer, F.C.; Wullschleger, S.D. Hydraulic Limitation of Tree Height: A Critique. Funct. Ecol. 2000, 14, 4–11. [Google Scholar] [CrossRef]
- Petit, G.; Mencuccini, M.; Carrer, M.; Prendin, A.L.; Hölttä, T. Axial conduit widening, tree height and height growth rate set the hydraulic transition of sapwood into heartwood. J. Exp. Bot. 2023, 74, 5072–5087. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hua, L.; Chen, P.; Luo, J.; Su, Y.; Li, J.; He, Q.; Yang, H. The Impact of Long-Term Dry-Season Irrigation on Eucalyptus Tree Height Growth: Insights from Leaf Photosynthesis and Water Conduction. Forests 2023, 14, 2017. https://doi.org/10.3390/f14102017
Hua L, Chen P, Luo J, Su Y, Li J, He Q, Yang H. The Impact of Long-Term Dry-Season Irrigation on Eucalyptus Tree Height Growth: Insights from Leaf Photosynthesis and Water Conduction. Forests. 2023; 14(10):2017. https://doi.org/10.3390/f14102017
Chicago/Turabian StyleHua, Lei, Penglong Chen, Jun Luo, Yan Su, Jiyue Li, Qian He, and Huizhu Yang. 2023. "The Impact of Long-Term Dry-Season Irrigation on Eucalyptus Tree Height Growth: Insights from Leaf Photosynthesis and Water Conduction" Forests 14, no. 10: 2017. https://doi.org/10.3390/f14102017
APA StyleHua, L., Chen, P., Luo, J., Su, Y., Li, J., He, Q., & Yang, H. (2023). The Impact of Long-Term Dry-Season Irrigation on Eucalyptus Tree Height Growth: Insights from Leaf Photosynthesis and Water Conduction. Forests, 14(10), 2017. https://doi.org/10.3390/f14102017