
HS-SPEM-GC-MS Analyses of Volatiles and Transcriptome Analysis of Camellia tetracocca in Response to Feeding by Empoasca onukii Matsuda


Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of C. tetracocca Leaves
2.2. Volatile Component Analysis Using the Leaves of C. tetracocca Attacked by E. onukii and Healthy Leaves
2.3. RNA Extraction, Library Construction, Sequencing, and Validation
2.4. RNA-Sequencing Data Analysis
2.5. Gene Ontology and Clustering Analysis
2.6. qPCR Validation
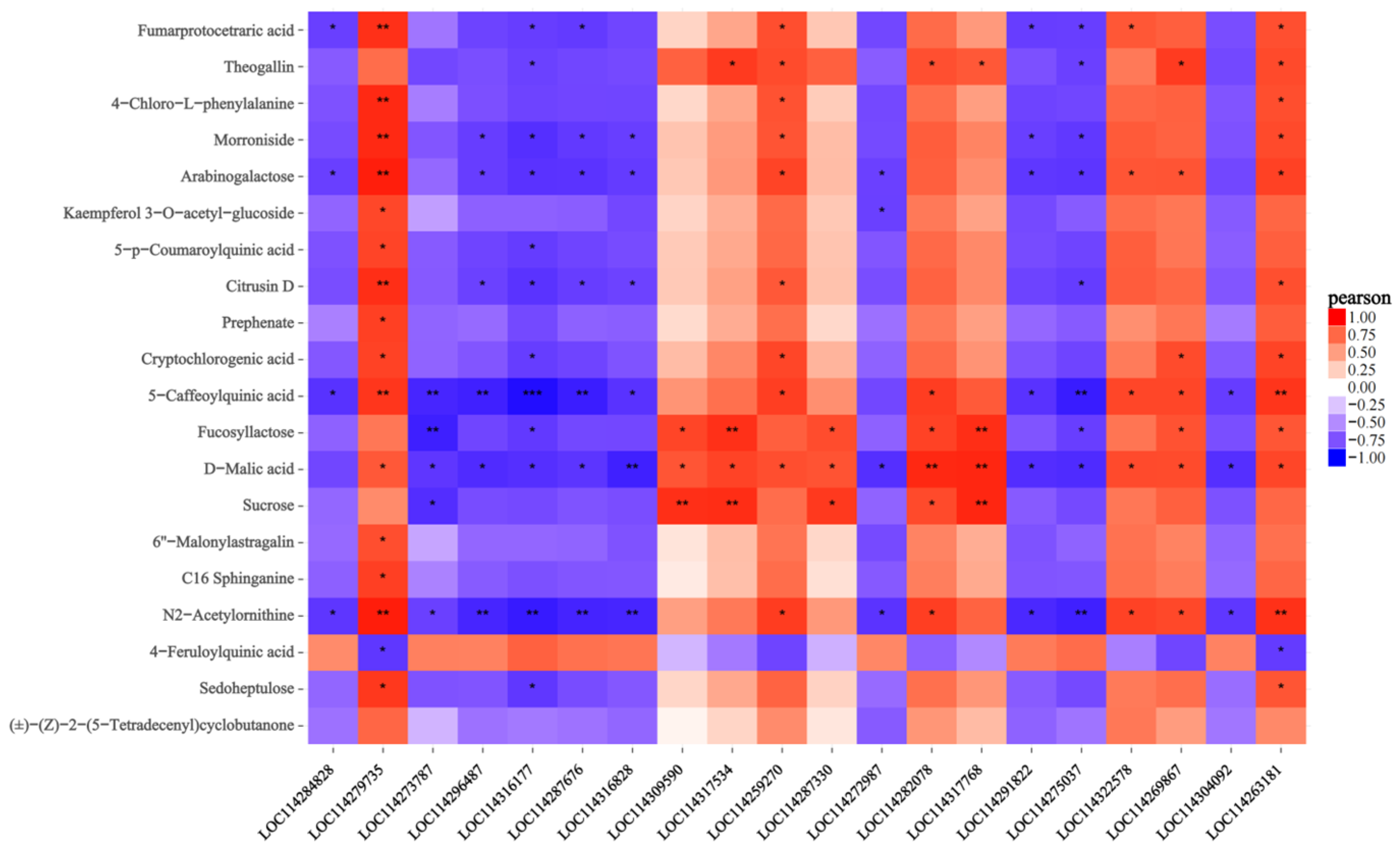
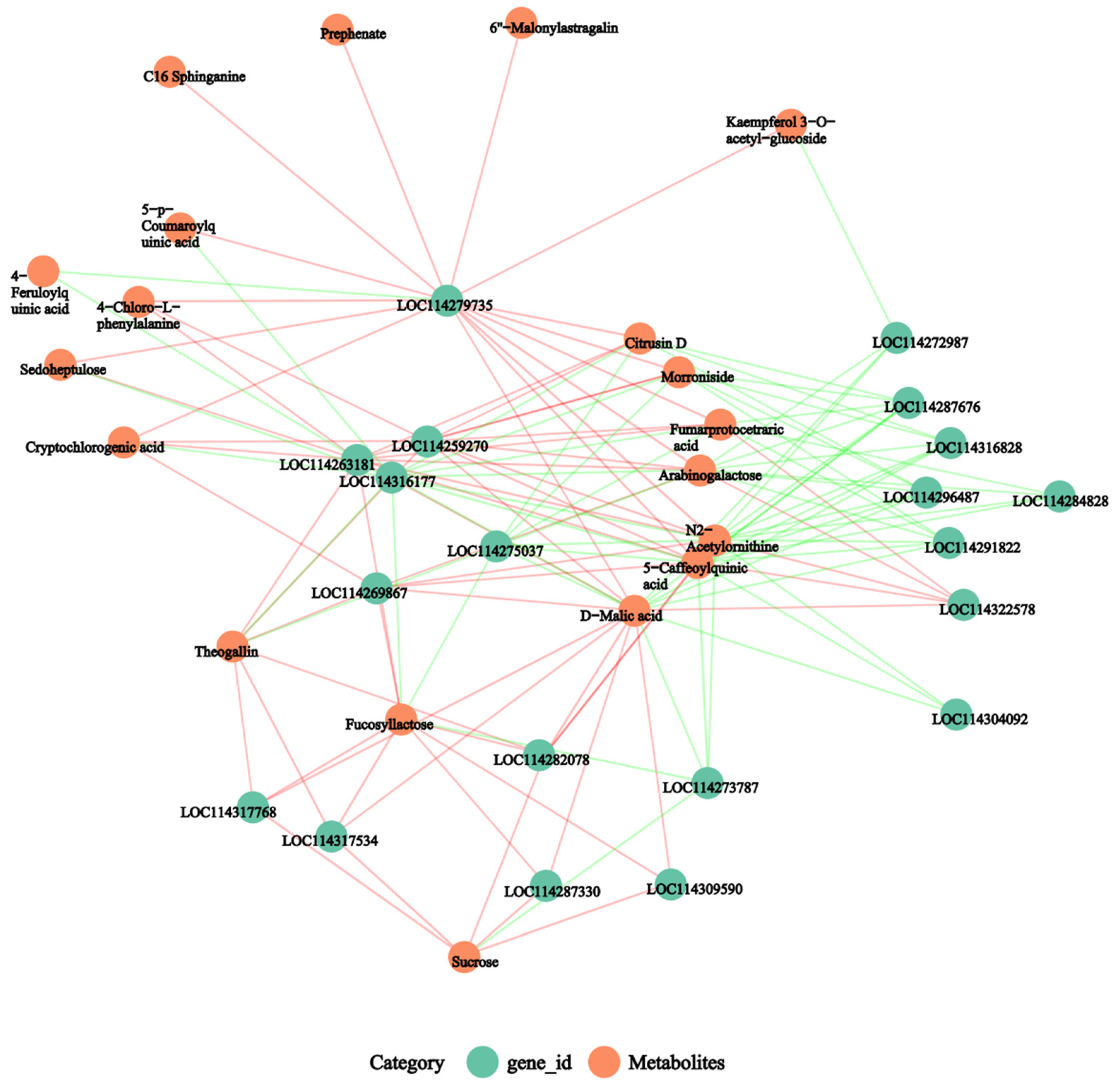
2.7. Joint Analysis of the Transcriptome and Metabolome
3. Results
3.1. Volatile Component Analysis of the Leaves of C. tetracocca Attacked by E. onukii and Healthy Leaves
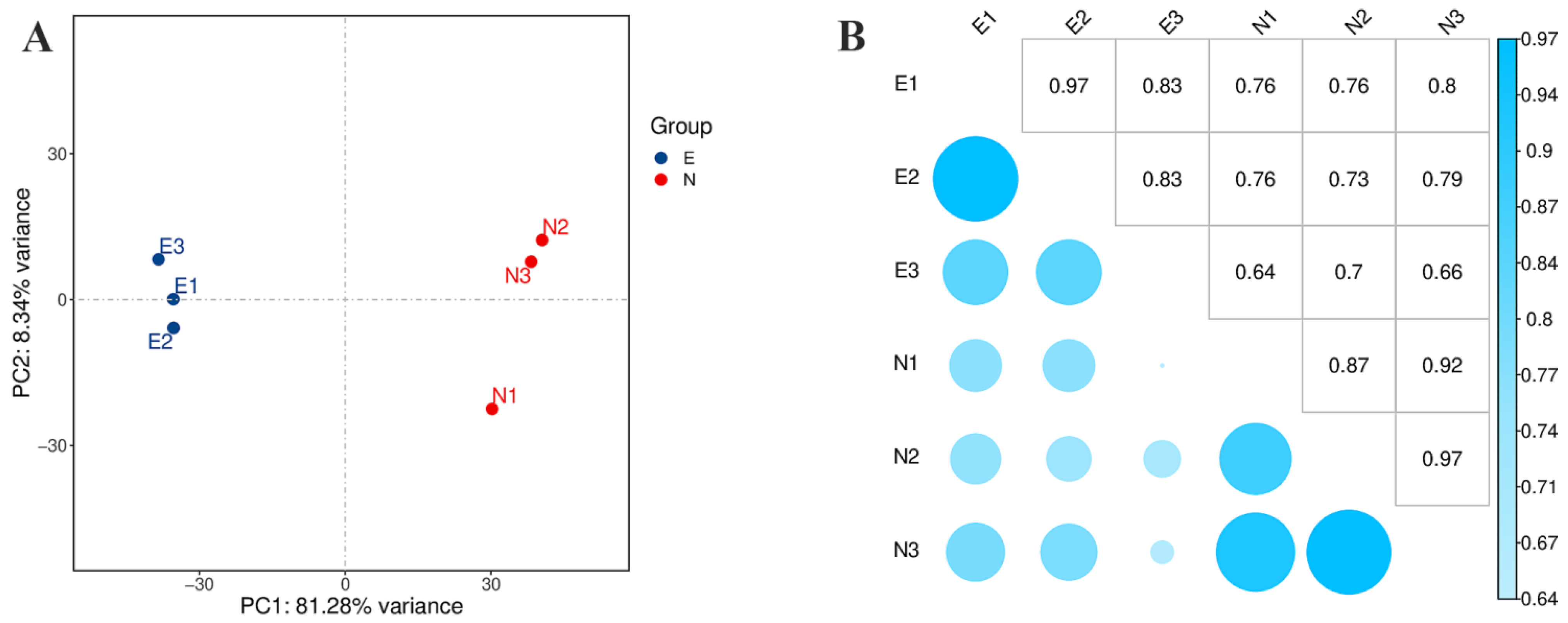
3.2. Sequencing and Assembly
3.3. Differential Expression of C. tetracocca Genes in Response to E. onukii
3.4. DEGs Associated with Phytohormones Biosynthesis and Signal Transduction
3.5. DEGs Associated with Secondary Metabolites
3.6. DEGs Associated with Plant–Pathogen Interaction and Defense Response to Insect
3.7. DEGs Associated with Transcription Factors
3.8. DEGs Associated with Protease Inhibitor, Chitinase, and MAPK
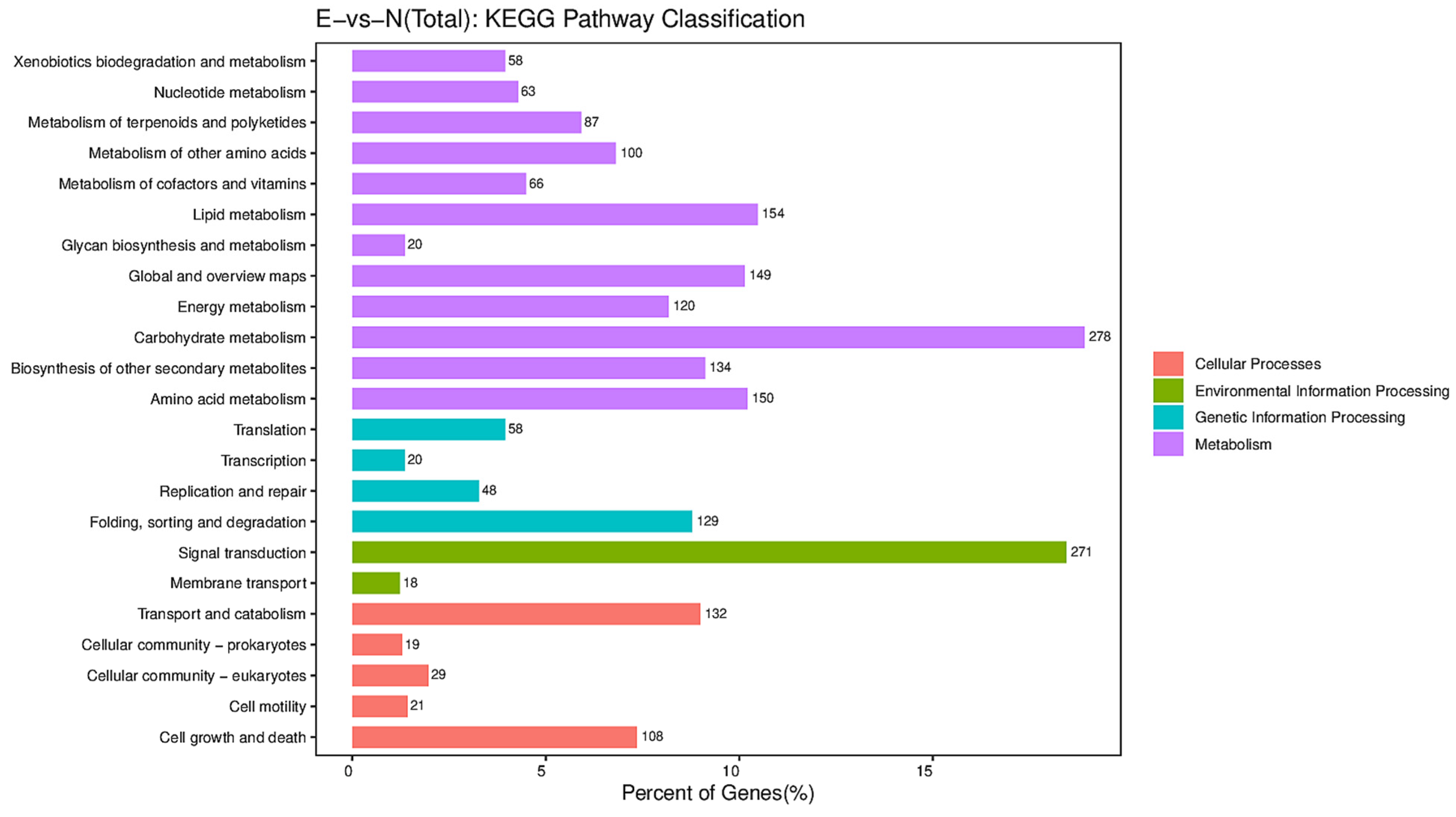
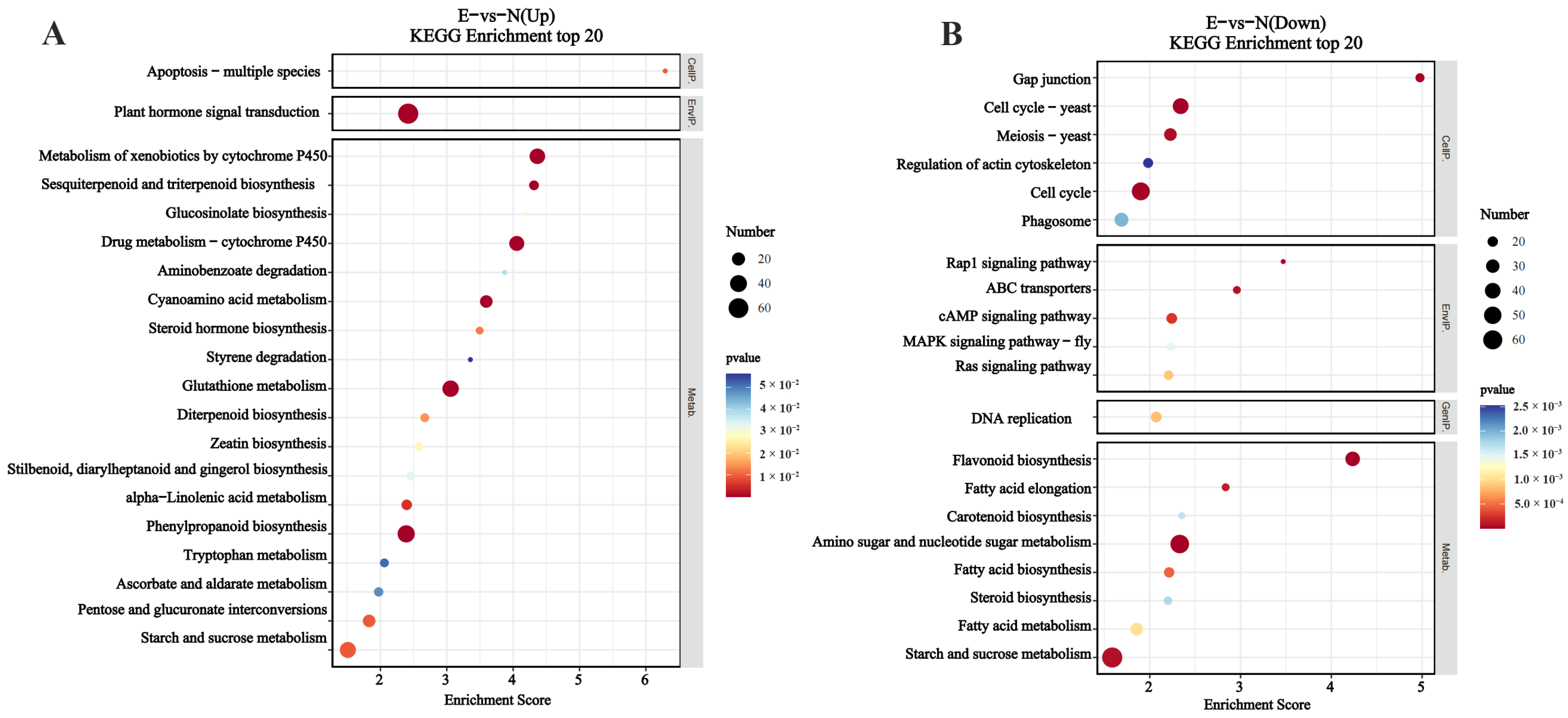
3.9. Functional Classification of DEGs
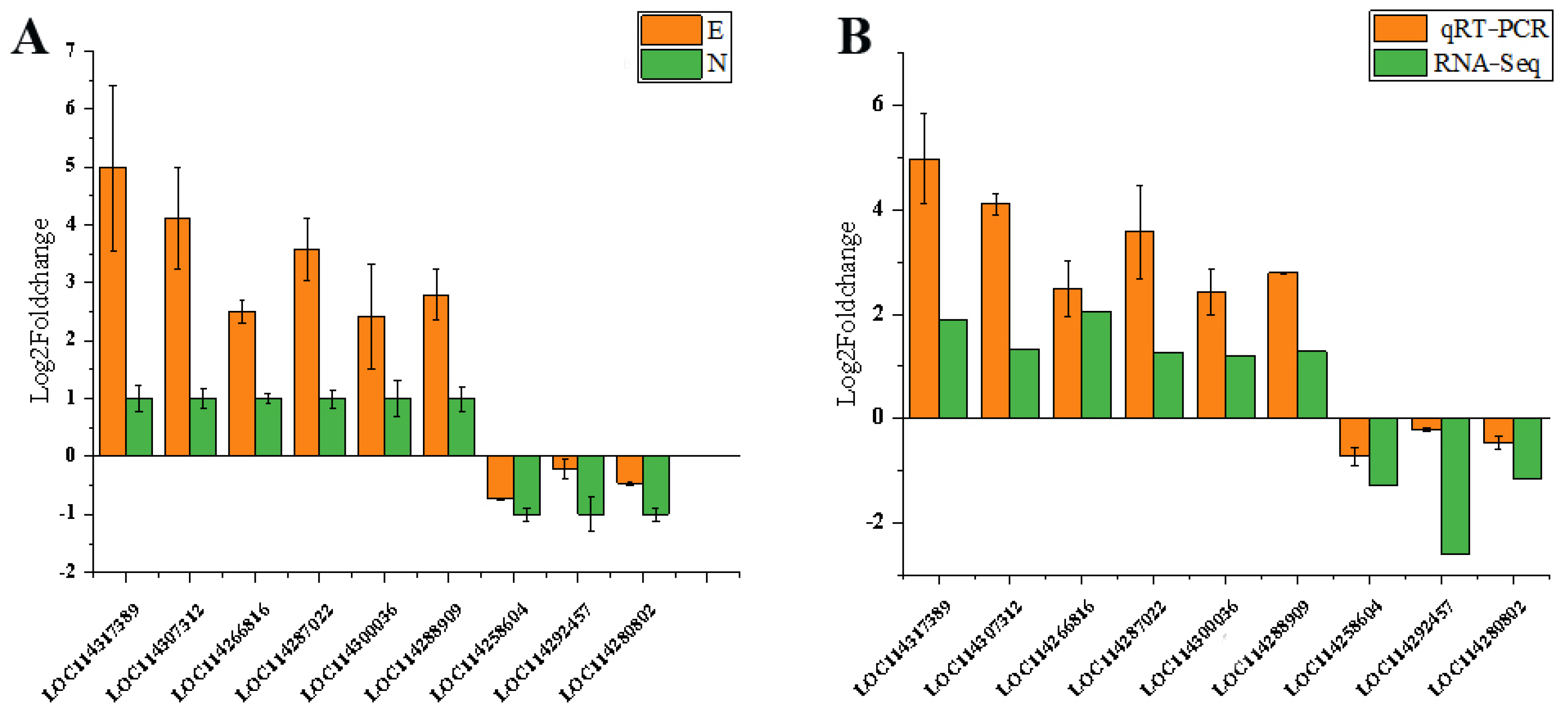
3.10. Validation of DEGs by qPCR
3.11. Joint Analysis of the Transcriptom
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhao, Q.; Shi, L.Q.; He, W.Y.; Li, J.Y.; You, S.J.; Chen, S.; Lin, J.; Wang, Y.B.; Zhang, L.W.; Yang, G.; et al. Genomic variations in the tea leafhopper reveal the basis of its adaptive evolution. Genom. Proteom. Bioinf. 2022, 20, 1092–1105. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.D. Systematic classification of tea plants. Acta Sci. Nat. Univ. Sunyatseni 1981, 1, 87–99. [Google Scholar]
- Yuan, M.Q.; Qian, C.J. Study on endemic plant Camellia tetracocca in Pu’an county, Guizhou. Guizhou Sci. 2009, 2, 80–85. [Google Scholar]
- Duan, X.Y.; Hu, H.J.; Wang, J.L.; Cao, Y.; Zhao, H.F. Present situation, protection and utilization of big tea tree Resources in southwest Guizhou. Agric. Technol. Serv. 2011, 28, 1736–1737. [Google Scholar]
- Chen, L.L.; Yuan, P.; Pozsgai, G.; Chen, P.; Zhu, H.P.; You, M.S. The impact of cover crops on the predatory mite Anystis baccarum (Acari, Anystidae) and the leafhopper pest Empoasca onukii (Hemiptera, Cicadellidae) in a tea plantation. Pest Manag. Sci. 2019, 75, 3371–3380. [Google Scholar] [CrossRef]
- Qin, D.; Zhang, L.; Xiao, Q.; Dietrich, C.; Matsumura, M. Clarification of the identity of the tea green leafhopper based on morphological comparison between Chinese and Japanese specimens. PLoS ONE 2015, 10, e0139202. [Google Scholar] [CrossRef]
- Zhang, N.; Tan, W.W.; Luo, G.M.; Pu, T.Y.; Wang, J.Q.; Zhang, X.H.; Song, Y.H. Untargeted metabolite profiling of Camellia tetracocca’s response to an Empoasca onukii attack using GC-MS and LC-MS. Agronomy 2023, 13, 791. [Google Scholar] [CrossRef]
- Mumm, R.; Hilker, M. Direct and indirect chemical defense of pine against folivorous insects. Trends Plant Sci. 2006, 11, 351–358. [Google Scholar] [CrossRef]
- Degenhardt, J. Indirect defense responses to herbivory in grasses. Plant Physiol. 2009, 149, 96–102. [Google Scholar] [CrossRef]
- Tamiru, A.; Bruce, T.J.A.; Woodcock, C.M.; Caulfield, J.C.; Midega, C.A.O.; Ogol, C.K.P.O.; Mayon, P.; Birkett, M.A.; Pickett, J.A.; Khan, Z.R. Maize landraces recruit egg and larval parasitoids in response to egg deposition by a herbivore. Ecol. Lett. 2011, 14, 1075–1083. [Google Scholar] [CrossRef]
- Bai, Y.C.; Yang, C.; Halitschke, R.; Paetz, C.; Kessler, D.; Burkard, K.; Gaquerel, E.; Baldwin, I.T.; Li, D. Natural history-guided omics reveals plant defensive chemistry against leafhopper pests. Science 2022, 375, eabm2948. [Google Scholar] [CrossRef] [PubMed]
- Xue, N.; Zhan, C.; Song, J.; Li, Y.; Zhang, J.; Qi, J.; Wu, J. The glutamate receptor-like 3.3 and 3.6 mediate systemic resistance to insect herbivores in Arabidopsis. J. Exp. Bot. 2022, 73, 7611–7627. [Google Scholar] [CrossRef] [PubMed]
- Afsah-Hejri, L.; Rajaram, P.; O’Leary, J.; McGivern, J.; Baxter, R.; Mesbah, A.; Maboudian, R.; Ehsani, R. Identification of volatile organic compounds (VOCs) by SPME-GC-MS to detect Aspergillus flavus infection in pistachios. Food Control 2023, 154, 110033. [Google Scholar] [CrossRef]
- Villagra, C.; Vera, W.; Lenitz, S.; Bergmann, J. Differences in volatile emissions between healthy and gall-induced branches of Haplopappus foliosus (Asteraceae). Biochem. Syst. Ecol. 2021, 98, 104309. [Google Scholar] [CrossRef]
- Chesnais, Q.; Golyaev, V.; Velt, A.; Rustenholz, C.; Brault, V.; Pooggin, M.M.; Drucker, M. Comparative plant transcriptome profiling of Arabidopsis thaliana Col-0 and Camelina sativa var. Celine infested with Myzus persicae aphids acquiring circulative and noncirculative viruses reveals virus- and plant-specific alterations relevant to aphid feeding behavior and transmission. Microbiol. Spectr. 2022, 10, 00136. [Google Scholar]
- Pamplona, R.S.; Kim, J.; Lee, J.W.; Kim, C.S.; Boo, K.H. Comparative transcriptome analysis of spinach in response to insect herbivory. Plant Biotechnol. Rep. 2022, 16, 43–55. [Google Scholar] [CrossRef]
- Wang, D.D.; Li, P.; Chen, Q.Y.; Chen, X.Y.; Yan, Z.W.; Wang, M.Y.; Mao, Y.B. Differential contributions of MYCs to insect defense reveals flavonoids alleviating growth inhibition caused by wounding in Arabidopsis. Front. Plant Sci. 2021, 12, 700555. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Liu, Q.S.; Du, L.X.; Hallerman, E.M.; Li, Y.H. Transcriptomic and metabolomic responses of rice plants to Cnaphalocrocis medinalis Caterpillar infestation. Insects 2022, 11, 705. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, E.L.; Huang, X.Z.; Kou, J.F.; Teng, D.; LV, B.B.; Han, X.Q.; Zhang, Y.J. Characterization of a novel insect-Induced sesquiterpene synthase GbTPS1 based on the transcriptome of Gossypium barbadense feeding by cotton bollworm. Front. Plant Sci. 2022, 13, 898541. [Google Scholar] [CrossRef]
- Zhao, X.M.; Chen, S.; Wang, W.N.; Wang, X.X.; Lin, Y.Z.; Su, F.; Yang, Z.B.; Yu, X.M. Defensive responses of tea plants (Camellia sinensis) against tea green leafhopper attack: A multi-omics study. Front. Plant Sci. 2020, 10, 1705. [Google Scholar] [CrossRef]
- Liu, H.F.; Li, S.F.; Xiao, G.S.; Wang, Q. Formation of volatiles in response to tea green leafhopper (Empoasca onukii Matsuda) herbivory in tea plants: A multi-omics study. Plant Cell Rep. 2021, 40, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Xiang, P.; Wilson, I.W.; Huang, J.; Zhu, Q.; Tan, M.; Lu, J.; Gao, S.; Zheng, S.; Lin, D.; Zhang, Y.; et al. Co-regulation of catechins biosynthesis responses to temperature changes by shoot growth and catechin related gene expression in tea plants (Camellia sinensis L.). J. Hortic. Sci. Biotechnol. 2021, 96, 228–238. [Google Scholar] [CrossRef]
- Fu, X.; Chen, Y.; Mei, X.; Katsuno, T.; Kobayashi, E.; Dong, F.; Watanabe, N.; Yang, Z. Regulation of formation of volatile compounds of tea (Camellia sinensis) leaves by single light wavelength. Sci. Rep. 2015, 5, 16858. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.F.; Zhou, Y.P.; Chen, Y.R.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Roberts, A.; Trapnell, C.; Donaghey, J.; Rinn, J.L.; Pachter, L. Improving RNA-Seq expression estimates by correcting for fragment bias. Genome Biol. 2011, 12, R22. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- The Gene Ontology Consortium. The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar] [CrossRef]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Glauser, G.; Lou, Y.G.; Erb, M.; Hu, L.F. Molecular dissection of early defense signaling underlying volatile-mediated defense regulation and herbivore resistance in rice. Plant Cell 2019, 31, 687–698. [Google Scholar] [CrossRef]
- Clavijo McCormick, A. Can plant–natural enemy communication withstand disruption by biotic and abiotic factors? Ecol. Evol. 2016, 6, 8569–8582. [Google Scholar] [CrossRef] [PubMed]
- Vivaldo, G.; Masi, E.; Taiti, C.; Caldarelli, G.; Mancuso, S. The network of plants volatile organic compounds. Sci. Rep. 2017, 7, 11050. [Google Scholar] [CrossRef]
- Juráň, S.; Grace, J.; Urban, O. Temporal changes in ozone concentrations and their impact on vegetation. Atmosphere 2021, 12, 82. [Google Scholar] [CrossRef]
- Ma, W.B.; Zhao, L.L.; Zhao, W.H.; Xie, Y.L. (E)-2-Hexenal, as a potential natural antifungal compound, inhibits Aspergillus flavus Spore germination by disrupting mitochondria! Energy metabolism. J. Agric. Food Chem. 2019, 67, 1138–1145. [Google Scholar] [CrossRef]
- Onnink, C.M.; Williams, A.A.; Williams, J.S. Electrophysiological and behavioral evidence for (E)-2-Hexenal as a female-attracting pheromone produced by disturbed Megacopta cribraria (Hemiptera: Plataspidae). J. Entomol. Sci. 2017, 52, 39–51. [Google Scholar]
- Berens, M.L.; Berry, H.M.; Mine, A.; Argueso, C.T.; Tsuda, K. Evolution of hormone signaling networks in plant defense. Annu. Rev. Phytopathol. 2017, 55, 401–425. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, S.; Sun, N.; Liu, H.; Zhao, Y.; Liang, Y.; Zhang, L.; Han, Y. Functional diversity of jasmonates in rice. Rice 2015, 8, 42. [Google Scholar] [CrossRef]
- Sun, Y.H.; Hung, C.Y.; Qiu, J.; Chen, J.J.; Kittur, F.S.; Oldham, C.E.; Henny, R.J.; Burkey, K.O.; Fan, L.J.; Xie, J.H. Accumulation of high OPDA level correlates with reduced ROS and elevated GSH benefiting white cell survival in variegated leaves. Sci. Rep. 2017, 7, 44158. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.J.; Lü, X.P.; Zhang, L.; Qiao, Y.; Zhao, Q.; Wang, Y.P.; Li, M.F.; Zhang, J.L. Transcriptomic profiling and physiological analysis of Haloxylon ammodendron in response to osmotic stress. Int. J. Mol. Sci. 2018, 19, 84. [Google Scholar] [CrossRef]
- Qi, X.H.; Chen, M.Y.; Liang, D.N.; Xu, Q.; Zhou, F.C.; Chen, X.H. Jasmonic acid, ethylene and ROS are involved in the response of cucumber (Cucumis sativus L.) to aphid infestation. Sci. Hortic. 2020, 269, 109421. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Wang, Q.J.; Guo, Z.R. Progresses on plant AP2/ERF transcription factors. Yi Chuan 2012, 34, 835–847. [Google Scholar] [CrossRef]
- Wu, J.; Gao, H.; Zhu, X.W.; Li, D.F. An ERF transcription factor enhances plant resistance to Myzus persicae and Spodoptera litura. Biotechnol. Biotechnol. Equip. 2020, 34, 946–954. [Google Scholar] [CrossRef]
- Li, J.T.; Chen, L.; Ding, X.; Fan, W.Y.; Liu, J.L. Transcriptome analysis reveals crosstalk between the Abscisic Acid and Jasmonic Acid signaling pathways in rice-mediated defense against Nilaparvata lugens. Int. J. Mol. Sci. 2022, 23, 6319. [Google Scholar] [CrossRef] [PubMed]
- Karban, R.; Agrawal, A.A.; Mangel, M. The benefits of induced defenses against herbivores. Ecology 1997, 78, 1351–1355. [Google Scholar] [CrossRef]
- Arimura, G.; Kost, C.; Boland, W. Herbivore-induced, indirect plant defenses. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2005, 1734, 91–111. [Google Scholar]
- Degenhardt, J.; Kollner, T.; Gershenzon, J. Monoterpene and sesquiterpene synthases and the origin of terpene skeletal diversity in plants. Phytochemistry 2009, 70, 1621–1637. [Google Scholar] [CrossRef]
- Lin, J.Y.; Wang, D.; Chen, X.L.; Köllner, T.G.; Mazarei, M.; Guo, H.; Pantalone, V.R.; Arelli, P.; Stewart, C.N., Jr.; Wang, N. An (E, E)-a-farnesene synthase gene of soybean has a role in defense against nematodes and is involved in synthesizing insect-induced volatiles. Plant Biotechnol. J. 2017, 15, 510–519. [Google Scholar] [CrossRef]
- Zhang, Z.B.; Liu, Z.X.; Li, S.R.; Xiong, T.; Ye, F.; Han, Y.T.; Sun, M.K.; Cao, J.J.; Luo, T.; Zhang, C.; et al. Effect of prior drought and heat stress on Camellia sinensis transcriptome changes to Ectropis oblique (Lepidoptera: Geometridae) resistance. Genomics 2022, 114, 110506. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.B.; Ji, S.X.; Yang, J.G.; Wang, X.W.; Han, W.H. Genome-wide analysis of MYB family in Nicotiana benthamiana and the functional role of the key members in resistance to Bemisia tabaci. Int. J. Biol. Macromol. 2023, 235, 123759. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.P.; Li, Y.; Pan, J.J.; Lou, D.J.; Hu, Y.R.; Yu, D.Q. The bHLH transcription factors MYC2, MYC3 and MYC4 are required for jasmonate-mediated inhibition of flowering in Arabidopsis. Mol. Plant 2017, 10, 1461–1464. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Gene ID | Forward Primer 5′-3′ | Reverse Primer 5′-3′ |
|---|---|---|---|
| 12-oxophytodienoate reductase | LOC114317389 | TCCTCTCCTCACTCCCTAC | ATGATCTGTTCCTTGTTAGTGG |
| Vinorine synthase | LOC114307312 | GACAGTATATCAACCAGTGCAT | ACAGAGTTGCTGCGTTAT |
| GABA transporter | LOC114266816 | CTGGACTTTGTTCTGCCTG | GCAACTGCTGCTATGACAC |
| Serine/threonine protein kinase PCRK1 | LOC114287022 | GCCTAAGCTTGTGACGAGTA | CTCTTCGATTGTAAAGCACCA |
| (+)-neomenthol dehydrogenase | LOC114300036 | TCATTGTTAACTCTGTCTGTCC | GGCAACAGAGCAAGTCTCA |
| Ethylene-responsive transcription factor ERF118 | LOC114288909 | ACTTGTTCTGGAGTAGAGTG | GAGTTGTTTCAAGTAAGAGGC |
| Cold-responsive protein kinase | LOC114258604 | ACCTTCCTCTCTACATGGAC | GCCACTTGACCCTTATCG |
| Chalcone-flavanone isomerase | LOC114292457 | GTCGCTTGCGGTTAAGTG | CGGAGACGATATCCCTGAA |
| Caffeoyl shikimate esterase | LOC114280802 | AGGATGCTTGCATGATCTACTA | ACGACTCAGCCAATAACCTT |
| Compounds | Molecular Formula | Molecular Weight | Percentage Content (%) (Damaged Leaves) | Percentage Content (%) (Healthy Leaves) |
|---|---|---|---|---|
| (Z)-2-penten-1-ol | C5H10O | 86.07 | 1.546 | 0.34 |
| 3-Hexenal | C6H10O | 98.07 | 0.546 | 1.128 |
| (E)-2-Hexenal | C6H10O | 98.07 | 12.653 | 2.841 |
| (E)-3-Hexen-1-ol | C6H12O | 100.09 | 21.076 | 22.523 |
| (Z)-2-Hexen-1-ol | C6H12O | 100.09 | 1.699 | 1.941 |
| 1-Hexanol | C6H14O | 102.1 | 1.931 | 1.685 |
| (E)-3-Hexen-1-ol-acetate | C8H14O2 | 142.1 | 1.337 | 6.694 |
| β-Ocimene | C10H16 | 136.13 | 1.077 | 0.772 |
| 1-Octanol | C8H18O | 130.14 | 0.577 | 0.313 |
| cis-2-Furanmethanol, 5-Ethenyltetrahydro-α,α, 5-trimethyl | C10H18O2 | 170.13 | 1.333 | 0.821 |
| Ethyl 2-(5-methyl-5-vinyltetrahydrofuran-2-yl)propan-2-yl carbonate | C13H22O4 | 242.15 | 1.89 | 1.769 |
| Linalool | C10H18O | 154.14 | 17.472 | 14.021 |
| (Z)-Butanoic acid, 3-hexenyl ester | C10H18O2 | 170.13 | 1.001 | 4.349 |
| Methyl salicylate | C8H8O3 | 152 | 7.649 | 18.568 |
| (Z)-Hexanoic acid, 3-hexenyl ester | C12H22O2 | 198.16 | 1.053 | 0.775 |
| 1H-3a,7-Methanoazulene, octahydro-3,8,8-trimethyl-6-methylene-, [3R-(3α, 3aβ, 7β, 8aα)] | C15H24 | 204.19 | 2.433 | 2.693 |
| α-Farnesene | C15H24 | 204.19 | 4.254 | 6.75 |
| Cedrol | C15H26O | 222.2 | 1.311 | 0.745 |
| Sample | Raw Reads (M) | Raw Bases (G) | Clean Reads (M) | Clean Bases (G) | Valid Bases (%) | Q30 (%) | GC (%) |
|---|---|---|---|---|---|---|---|
| E1 | 51.87 | 7.78 | 51.73 | 7.49 | 96.3 | 91.75 | 44.68 |
| E2 | 56.3 | 8.44 | 56.15 | 8.14 | 96.42 | 92.17 | 44.79 |
| E3 | 48.5 | 7.28 | 48.37 | 7.02 | 96.46 | 91.95 | 46.19 |
| N1 | 54.94 | 8.24 | 54.78 | 7.93 | 96.18 | 91.73 | 45.1 |
| N2 | 51.55 | 7.73 | 51.43 | 7.48 | 96.7 | 92.33 | 45.49 |
| N3 | 51.35 | 7.7 | 51.22 | 7.39 | 95.96 | 92.16 | 45.23 |
| Sample | Total Reads | Total Mapped Reads | Mapped Rate |
|---|---|---|---|
| E1 | 51,732,120 | 41,868,448 | 80.93% |
| E2 | 56,151,682 | 44,204,355 | 78.72% |
| E3 | 48,370,234 | 39,981,306 | 82.66% |
| N1 | 54,783,750 | 45,083,014 | 82.29% |
| N2 | 51,428,726 | 42,485,786 | 82.61% |
| N3 | 51,217,692 | 42,052,676 | 82.11% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, N.; Wang, J.; Pu, T.; Zhao, Z.; Tan, W.; Yu, Z.; Song, Y. HS-SPEM-GC-MS Analyses of Volatiles and Transcriptome Analysis of Camellia tetracocca in Response to Feeding by Empoasca onukii Matsuda. Forests 2023, 14, 2059. https://doi.org/10.3390/f14102059
Zhang N, Wang J, Pu T, Zhao Z, Tan W, Yu Z, Song Y. HS-SPEM-GC-MS Analyses of Volatiles and Transcriptome Analysis of Camellia tetracocca in Response to Feeding by Empoasca onukii Matsuda. Forests. 2023; 14(10):2059. https://doi.org/10.3390/f14102059
Chicago/Turabian StyleZhang, Ni, Jinqiu Wang, Tianyi Pu, Zhibing Zhao, Weiwen Tan, Zhengwen Yu, and Yuehua Song. 2023. "HS-SPEM-GC-MS Analyses of Volatiles and Transcriptome Analysis of Camellia tetracocca in Response to Feeding by Empoasca onukii Matsuda" Forests 14, no. 10: 2059. https://doi.org/10.3390/f14102059
APA StyleZhang, N., Wang, J., Pu, T., Zhao, Z., Tan, W., Yu, Z., & Song, Y. (2023). HS-SPEM-GC-MS Analyses of Volatiles and Transcriptome Analysis of Camellia tetracocca in Response to Feeding by Empoasca onukii Matsuda. Forests, 14(10), 2059. https://doi.org/10.3390/f14102059