
Exogenous Application of Salicylic Acid Improves Physiological and Biochemical Attributes of Morus alba Saplings under Soil Water Deficit

 , , ,
, , ,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Planting Material and Treatment
2.2. Growth and Dry Biomass Production
2.3. The Physiological Parameters, Chlorophyll a, b and Carotenoid Contents
2.4. The Gas Exchange Measurements
2.5. Proline, Soluble Sugar, Soluble Protein and Total Phenolic Contents
2.6. Lipid Peroxidation, MDA Contents and Electrolyte Leakage (EL %)
2.7. Oxidants, Hydrogen Peroxide (H2O2) and Superoxide Radical (O2−) Measurements
2.8. Antioxidants’ Enzyme Activities
2.9. Statistical Analysis
3. Results
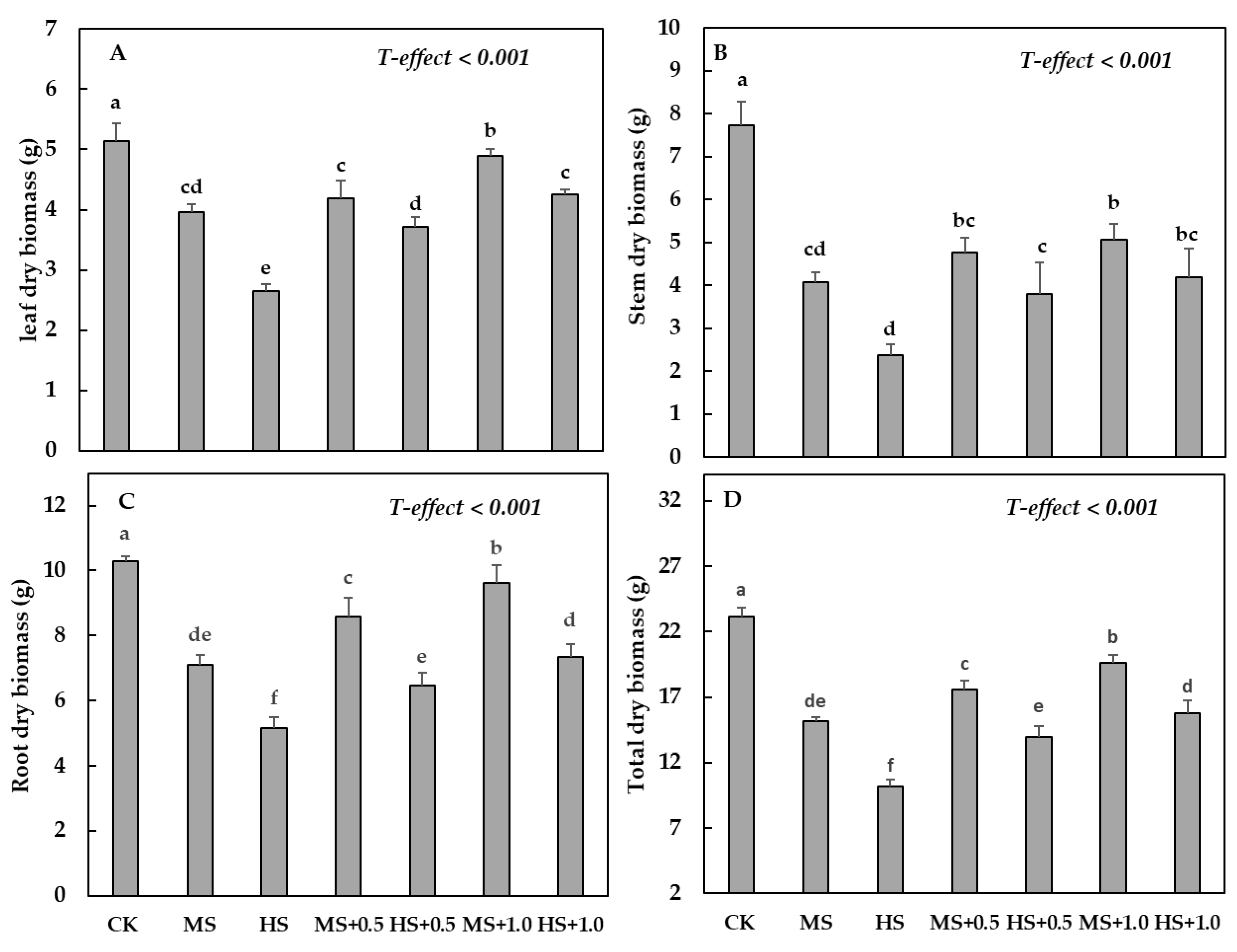
3.1. Water Deficit Treatments and Application of SA on Morphological Attributes
3.2. Water Deficit Treatments and Application of SA on Photosynthetic Pigments
3.3. Water Deficit Treatments and Application of SA on Gas Exchange Attributes
3.4. Water Deficit Treatments and Effect of SA on Osmolyte Accumulation

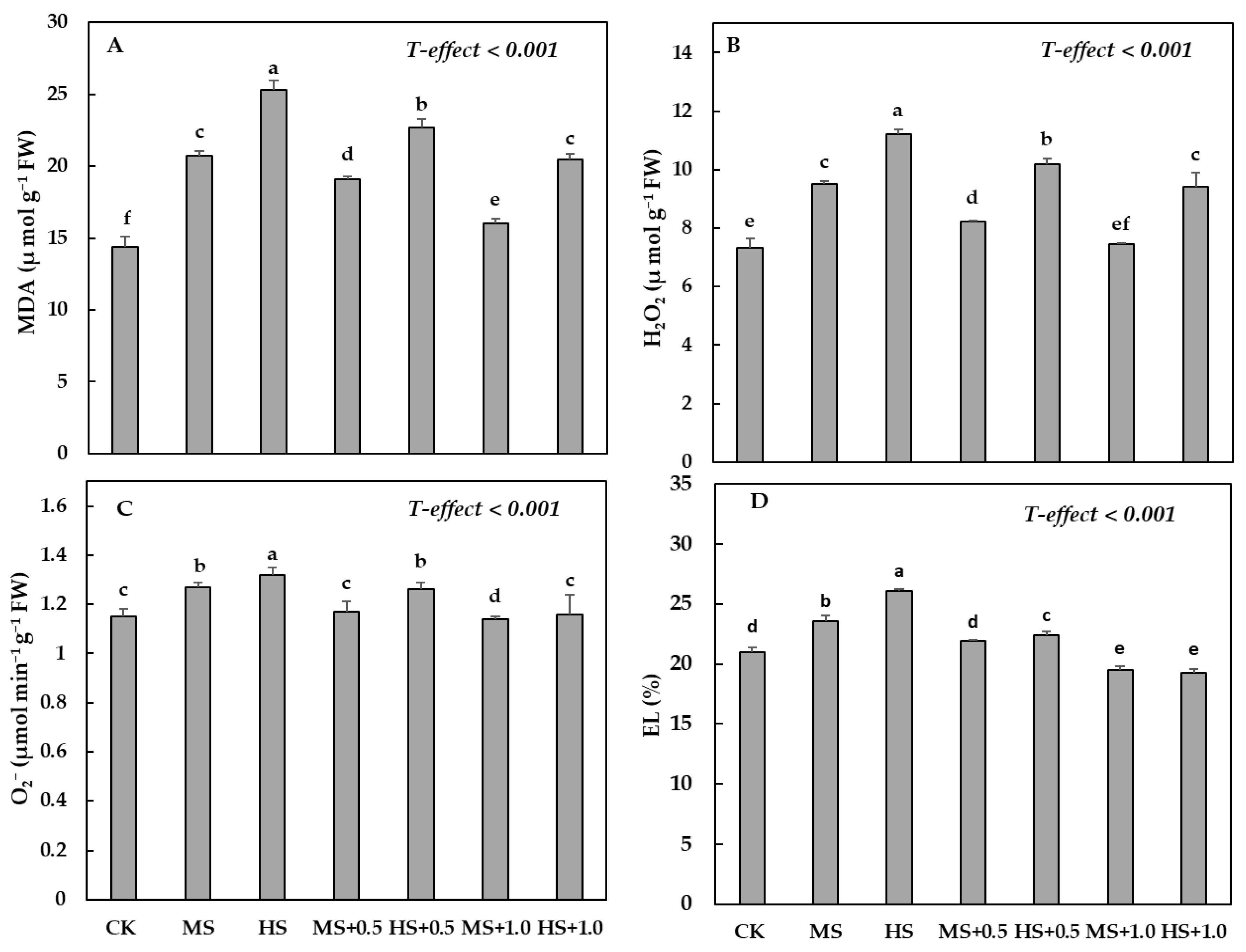
3.5. Water Deficit Treatments and Effect of SA on MDA, H2O2, O2− and EL%
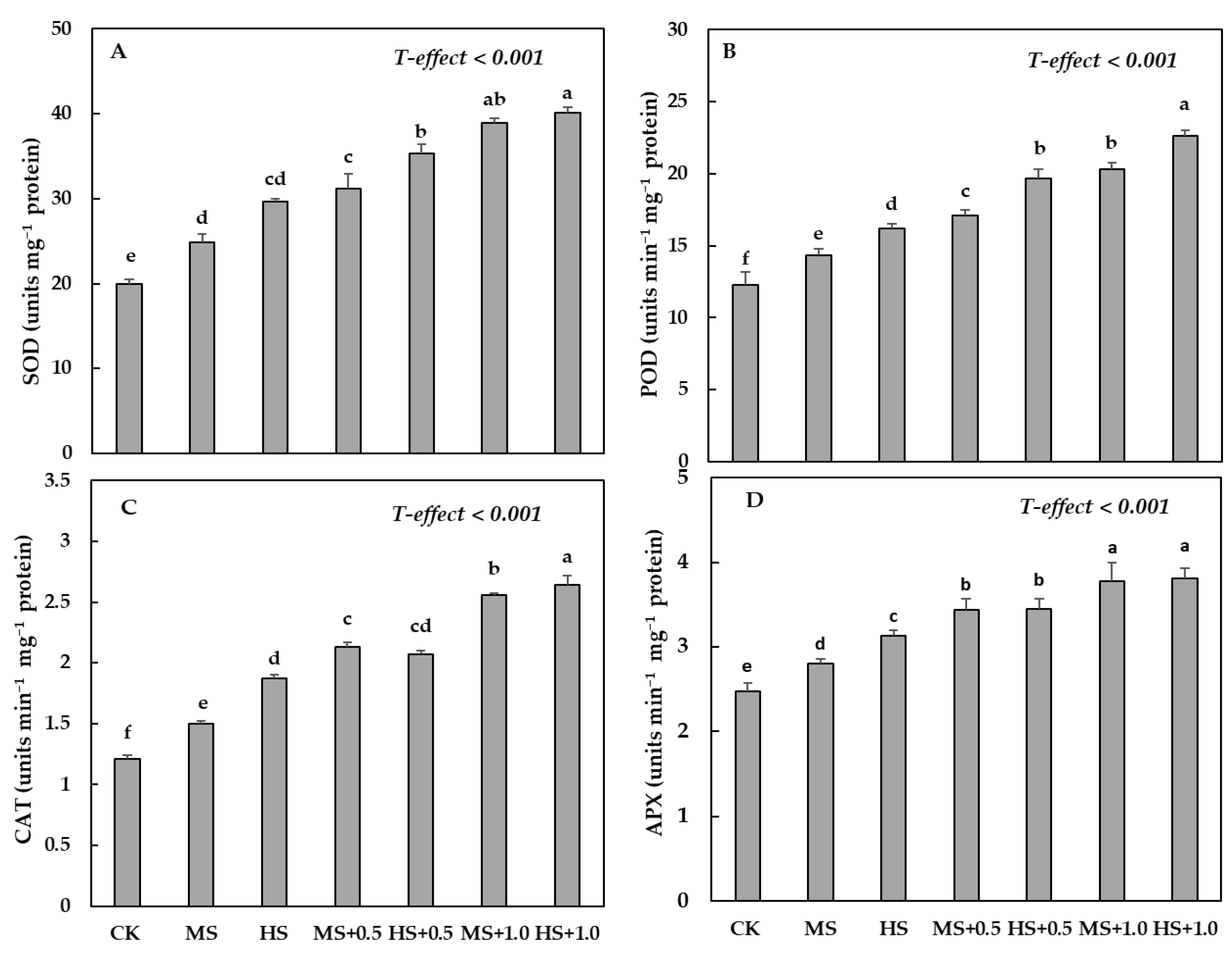
3.6. Water Deficit Treatments and Effect of SA on Antioxidant Enzymes Activities
4. Discussion
4.1. Plant Growth and Dry Biomass Production
4.2. The Physiological Parameters, Chlorophyll a, b and Carotenoid Contents
4.3. The Osmolyte Accumulation
4.4. H2O2, O2−, along with MDA and EL% and Antioxidants
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought-from genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef] [PubMed]
- Stuart, M.E.; Gooddy, D.C.; Bloomfield, J.P.; Williams, A.T. A review of the impact of climate change on future nitrate concentrations in groundwater of the UK. Sci. Total Environ. 2011, 409, 2859–2873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IPCC. Executive Summary of the Intergovernmental Panel on Climate Change, February 2007. Available online: www.ipcc.com.ch (accessed on 21 June 2022).
- Liu, C.; Liu, Y.; Guo, K.; Fan, D.; Li, G.; Zheng, Y.; Yu, L.; Yang, R. Effect of drought on pigments, osmotic adjustment and antioxidant enzymes in six woody plant species in karst habitats of southwestern China. Environ. Exp. Bot. 2011, 71, 174–183. [Google Scholar] [CrossRef]
- Eswaran, H.; Reich, P.; Beinroth, F. Global desertification tension zones. In Proceedings of the 10th International Soil Conservation Organization Meeting, West Lafayette, IN, USA, 24–29 May 2001. [Google Scholar]
- Aryal, J.P.; Sapkota, T.B.; Khurana, R.; Khatri-Chhetri, A.; Rahut, D.B.; Jat, M.L. Climate change and agriculture in South Asia: Adaptation options in smallholder production systems. Environ. Dev. Sustain. 2020, 22, 5045–5075. [Google Scholar] [CrossRef] [Green Version]
- Anjum, S.; Saleem, M.; Cheema, M.; Bilal, M.; Khaliq, T. An assessment to vulnerability, extent, characteristics and severity of drought hazard in Pakistan. Pak. J. Sci. 2012, 64, 85–96. [Google Scholar]
- Park Williams, A.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D.; et al. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Chang. 2013, 3, 292–297. [Google Scholar] [CrossRef]
- Scholze, M.; Knorr, W.; Arnell, N.W.; Prentice, I. A climate-change risk analysis for world ecosystems. Proc. Natl. Acad. Sci. USA 2006, 103, 13116–13120. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.T.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 4, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Rice, K.J.; Matzner, S.L.; Byer, W.; Brown, J.R. Patterns of tree dieback in Queensland, Australia: The importance of drought stress and the role of resistance to cavitation. Oecologia 2004, 139, 190–198. [Google Scholar] [CrossRef]
- Hoffmann, W.A.; Marchin, R.M.; Abit, P.; Lau, O.L. Hydraulic failure and tree dieback are associated with high wood density in a temperate forest under extreme drought. Glob. Chang. Biol. 2011, 17, 2731–2742. [Google Scholar] [CrossRef]
- Sperry, J.S.; Sullivan, J.E.M. Xylem embolism in response to freeze thaw cycles and water stress in ringporous, diffuse-porous, and conifer species. Plant Physiol. 1992, 100, 605–613. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.G.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.E.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Badel, E.; Burlett, R.; Delzon, S.; Cochard, H.; Jansen, S. Non-invasive measurement of vulnerability to drought-induced embolism by X-Ray microtomography. Plant Physiol. 2016, 170, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, C.B.; Rouina, B.; Sensoy, S.; Boukhris, M.; Abdallah, F.B. Changes in gas exchange, proline accumulation and antioxidative enzyme activities in three olive cultivars under contrasting water availability regimes. Environ. Exp. Bot. 2009, 67, 345–352. [Google Scholar] [CrossRef]
- Rasheed, F.; Gondal, A.; Kudus, K.A.; Zafar, Z.; Nawaz, M.F.; Khan, W.R.; Abdullah, M.; Ibrahim, F.H.; Depardieu, C.; Pazi, A.M.M.; et al. Effects of soil water deficit on three tree species of the arid environment: Variations in growth, physiology, and antioxidant enzyme activities. Sustainability. 2021, 13, 3336. [Google Scholar] [CrossRef]
- Bacelar, E.A.; Santos, D.L.; Moutinho-Pereira, J.M.; Lopes, J.I.; Goncalves, B.C.; Ferreira, T.C.; Correia, C.M. Physiological behaviour, oxidative damage and antioxidative protection of olive trees grown under different irrigation regimes. Plant Soil 2007, 292, 1. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Penuelas, J. Photo and antioxidative protection, and a role for salicylic acid during drought and recovery in field grown Phillyrea angustifolia plants. Planta 2003, 217, 758–766. [Google Scholar] [CrossRef]
- Zarafshar, M.; Akbarinia, M.; Askari, H.; Hosseini, S.M.; Rahaie, M.; Struve, D.; Striker, G.G. Morphological, physiological and biochemical responses to soil water deficit in seedlings of three populations of wild pear (Pyrus boisseriana). Biotechnol. Agron. Soc. Environ. 2014, 18, 353–366. [Google Scholar]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Arbona, V.; Manzi, M.; Cd, O.; Gómez-Cadenas, A. Metabolomics as a tool to investigate abiotic stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 4885–4911. [Google Scholar] [CrossRef]
- Zafar, Z.; Rasheed, F.; Atif, R.M.; Javed, M.A.; Maqsood, M.; Gailing, O. Foliar application of salicylic acid improves water stress tolerance in Conocarpus erectus L. and Populus deltoides L. saplings: Evidence from morphological, physiological, and biochemical changes. Plants 2021, 10, 1242. [Google Scholar] [CrossRef] [PubMed]
- Zafar, Z.; Rasheed, F.; Atif, R.M.; Maqsood, M.; Gailing, O. Salicylic acid-induced morpho-physiological and biochemical changes triggered water deficit tolerance in Syzygium cumini L. saplings. Forests 2021, 12, 491. [Google Scholar] [CrossRef]
- Zafar, Z.; Rasheed, F.; Abdullah, M.; Salam, M.M.A.; Mohsin, M. Effects of water deficit on growth and physiology of young Conocarpus erectus L. and Ficus benjamina L. saplings. Bangladesh J. Bot. 2019, 48, 1215–1221. [Google Scholar] [CrossRef]
- Noctor, G.; Reichheld, J.P.; Foyer, C.H. ROS related redox regulation and signaling in plants. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2018; Volume 80, pp. 3–12. [Google Scholar]
- Laxa, M.; Michael, L.; Wilena, T.; Kamel, C.; Karl, J.D. The role of the plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and drought stresses in crop plants Implications, cross talk, and potential management opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.R.; Fatma, M.; Per, T.S.; Anjum, N.A.; Khan, N.A. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front. Plant Sci. 2015, 6, 462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayat, S.; Hasan, S.A.; Fariduddin, Q.; Ahmad, A. Growth of tomato (Lycopersicon esculentum) in response to salicylic acid under water stress. J. Plant Interact. 2008, 3, 297–304. [Google Scholar] [CrossRef]
- Horvath, E.; Szalai, G.; Janda, T. Induction of abiotic stress tolerance by salicylic acid signaling. J. Plant Growth Regul. 2007, 26, 290–300. [Google Scholar] [CrossRef]
- Belkadhi, A.; De Haro, A.; Obregon, S.; Chaıbi, W.; Djebali, W. Positive effects of salicylic acid pretreatment on the composition of flax plastidial membrane lipids under cadmium stress. Environ. Sci. Pollut. Res. 2015, 22, 1457–1467. [Google Scholar] [CrossRef]
- Janda, T.; Gondor, O.K.; Yordanova, R.; Szalai, G.; Pál, M. Salicylic acid and photosynthesis: Signaling and effects. Acta Physiol. Plant. 2014, 36, 2537–2546. [Google Scholar] [CrossRef] [Green Version]
- Reddy, A.R.; Chaitanya, K.V.; Jutur, P.P.; Sumithra, K. Differential antioxidative responses to water stress among five mulberry (Morus alba L.) cultivars. Environ. Exp. Bot. 2004, 52, 33–42. [Google Scholar] [CrossRef]
- Dhanyalakshmi, K.H.; Nataraja, K.N. Mulberry (Morus spp.) has the features to treat as a potential perennial model system. Plant Signal. Behav. 2018, 13, 1491267. [Google Scholar] [CrossRef] [PubMed]
- Butt, M.S.; Nazir, A.; Sultan, M.T.; Schroën, K. Morus alba L. nature’s functional tonic. Trends Food Sci. Tech. 2008, 19, 505–512. [Google Scholar] [CrossRef]
- Papanastasis, V.P.; Yiakoulaki, M.D.; Decandia, M.; Dini-Papanastasi, O. Integrating woody species into livestock feeding in the Mediterranean areas of Europe. Anim. Feed. Sci. Technol. 2008, 140, 1–17. [Google Scholar] [CrossRef]
- Rasheed, F.; Dreyer, E.; Richard, B.; Brignolas, F.; Brendel, O.; Thiec, D.L. Vapour pressure deficit during growth has little impact on genotypic differences of transpiration efficiency at leaf and whole-plant level: An example from Populus nigra L. Plant Cell Environ. 2015, 38, 670–684. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Valentovic, P.; Luxova, M.; Kolarovic, L.; Gasparikova, O. Effect of osmotic stress on compatible solutes content, membrane stability and water relations in two maize cultivars. Plant Soil Environ. 2006, 52, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Nayyar, H. Accumulation of osmolytes and osmotic adjustment in water-stressed wheat (Triticum aestivum) and maize (Zea mays) as affected by calcium and its antagonists. Environ. Exp. Bot. 2003, 50, 253–264. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Bai, T.; Li, C.; Ma, F.; Feng, F.; Shu, H. Responses of growth and antioxidant system to root-zone hypoxia stress in two Malus species. Plant Soil 2010, 327, 95–105. [Google Scholar] [CrossRef]
- Bayer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Maehly, A.C.; Chance, B. The assay of catalases and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Knörzer, O.C.; Burner, J.; Boger, P. Alterations in the antioxidative system of suspension-cultured soybean cells (Glycine max) induced by oxidative stress. Physiol. Plant. 1996, 97, 388–396. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Sabir, M.A.; Rasheed, F.; Zafar, Z.; Khan, I.; Nawaz, M.F.; Haq, I.U.; Bilal, M. A consistent CO2 assimilation rate and an enhanced root development drives the tolerance mechanism in Ziziphus jujuba under soil water deficit. Arid. Land Res. Manag. 2020, 34, 392–404. [Google Scholar] [CrossRef]
- Petrík, P.; Petek, A.; Konôpková, A.; Bosela, M.; Fleischer, P.; Frýdl, J.; Kurjak, D. Stomatal and leaf morphology response of European Beech (Fagus sylvatica L.) provenances transferred to contrasting climatic conditions. Forests 2020, 11, 1359. [Google Scholar] [CrossRef]
- Muller, B.; Pantin, F.; Genard, M.; Turc, O.; Freixes, S.; Piques, M.; Gibon, Y. Water deficits uncouple growth from photosynthesis, increase C content, and modify the relationships between C and growth in sink organs. J. Exp. Bot. 2011, 62, 1715–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, C.; Hu, Y.; Du, X.; Li, T.; Tang, H.; Wu, J. Salicylic acid induces physiological and biochemical changes in Torreya grandis cv. Merrillii seedlings under drought stress. Trees 2014, 28, 961–970. [Google Scholar]
- Brito, C.; Dinis, L.; Ferreira, H.; Moutinho-Pereira, J.; Correia, C. The role of nighttime water balance on Olea europaea plants subjected to contrasting water regimes. J. Plant Physiol. 2018, 226, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Jesus, C.; Meijón, M.; Monteiro, P.; Correia, B.; Amaral, J.; Escandón, M.; Cañal, J.M.; Pinto, G. Salicylic acid application modulates physiological and hormonal changes in Eucalyptus globulus under water deficit. Environ. Exp. Bot. 2015, 118, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Sakhabutdinova, A.R.; Fatkhutdinova, D.R.; Bezrukova, M.V.; Shakiova, F.M. Salicylic acid prevents the damaging action of stress factors on wheat plants. Bulg. J. Plant Physiol. 2003, 29, 314–319. [Google Scholar]
- Flexas, J.; Medrano, H. Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef]
- Saheri, F.; Barzin, G.; Pishkar, L.; Boojar, M.M.A.; Babaeekhou, L. Foliar spray of salicylic acid induces physiological and biochemical changes in purslane (Portulaca oleracea L.) under drought stress. Biologia 2020, 75, 2189–2200. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Sharkey, T.K. Stomatal conductance and photosynthesis. Ann. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Blum, A. Plant water relations, plant stress and plant production. In Plant Breeding for Water-Limited Environments; Springer: New York, NY, USA, 2011; pp. 11–52. [Google Scholar]
- He, F.; Sheng, M.; Tang, M. Effects of Rhizophagus irregularis on photosynthesis and antioxidative enzymatic system in Robinia pseudoacacia L. under drought stress. Front. Plant Sci. 2017, 8, 183. [Google Scholar] [CrossRef] [Green Version]
- Tahjib-Ul-Arif, M.; Siddiqui, M.N.; Sohag, A.A.M.; Sakil, M.A.; Rahman, M.M.; Polash, M.A.S.; Mostofa, M.G.; Tran, L.S.P. Salicylic acid-mediated enhancement of photosynthesis attributes and antioxidant capacity contributes to yield improvement of maize plants under salt stress. J. Plant Growth Regul. 2018, 37, 1318–1330. [Google Scholar] [CrossRef]
- White, D.A.; Turner, N.C.; Galbraith, J.H. Leaf water relations and stomatal behavior of four allopatric Eucalyptus species planted in Mediterranean southwestern Australia. Tree Physiol. 2000, 20, 1157–1165. [Google Scholar] [CrossRef]
- Cotrozzi, L.; Remorini, D.; Pellegrini, E.; Landi, M.; Massai, R.; Nali, C.; Guidi, L.; Lorenzini, G. Variations in physiological and biochemical traits of oak seedlings grown under drought and ozone stress. Physiol. Plant. 2016, 157, 69–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghorbani, A.; Razavi, S.M.; Omran, V.O.G.; Pirdashti, H. Piriformospora indica alleviates salinity by boosting redox poise and antioxidative potential of tomato. Russ. J. Plant Physiol. 2018, 65, 898–907. [Google Scholar] [CrossRef]
- Idrees, M.; Khan, M.M.A.; Aftab, T.; Naeem, M.; Hashmi, N. Salicylic acid-induced physiological and biochemical changes in lemongrass varieties under water stress. J. Plant Interact. 2010, 5, 293–303. [Google Scholar] [CrossRef]
- Chavoushi, M.; Najafi, F.; Salimi, A.; Angaji, S.A. Improvement in drought stress tolerance of safflower during vegetative growth by exogenous application of salicylic acid and sodium nitroprusside. Ind. Crop. Prod. 2019, 134, 168–176. [Google Scholar] [CrossRef]
- Zhu, J.J.; Zhang, J.L.; Liu, H.C.; Cao, K.F. Photosynthesis, non-photochemical pathways and activities of antioxidant enzymes in a resilient evergreen oak under different climatic conditions from a valley-savanna in Southwest China. Physiol. Plant. 2009, 135, 62–72. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Dianat, M.; Saharkhiz, M.J.; Tavassolian, I. Salicylic acid mitigates drought stress in Lippia citriodora L. effects on biochemical traits and essential oil yield. Biocatal. Agric. Biotechnol. 2016, 8, 286–293. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Plant Height (cm) | Stem Diameter (mm) | No. of Leaves | Root/Shoot Ratio | Chlorophyll a (mg g−1 FW) | Chlorophyll b (mg g−1 FW) | Carotenoids (mg g−1 FW) | |
|---|---|---|---|---|---|---|---|
| CK | 53.0 ± 1.60 b | 4.49 ± 0.24 a | 40.4 ± 0.50 a | 0.64 ± 0.04 b,c | 1.43 ± 0.06 a | 1.57 ± 0.10 a | 0.97 ± 0.07 a |
| MS | 40.3 ± 1.34 d | 3.67 ± 0.15 c | 20.0 ± 0.70 f | 0.75 ± 0.04 a | 1.16 ± 0.02 e | 1.07 ± 0.02 f | 0.72 ± 0.03 f |
| HS | 31.7 ± 1.37 e | 3.02 ± 0.06 f | 26.0 ± 0.94 e | 0.66 ± 0.05 c | 0.92 ± 0.04 g | 0.96 ± 0.01 g | 0.66 ± 0.03 g |
| MS + 0.5 | 51.0 ± 1.29 b | 3.84 ± 0.30 b,c | 33.3 ± 0.86 d | 0.69 ± 0.05 b | 1.23 ± 0.00 d | 1.17 ± 0.01 d | 0.75 ± 0.03 e |
| HS + 0.5 | 47.2 ± 0.76 c | 3.29 ± 0.10 e | 34.2 ± 1.78 c,d | 0.63 ± 0.09 c | 1.13 ± 0.02 f | 1.13 ± 0.01 e | 0.78 ± 0.01 d |
| MS + 1.0 | 58.6 ± 1.47 a | 3.91 ± 0.08 b | 35.6 ± 0.87 c | 0.74 ± 0.06 a,b | 1.34 ± 0.01 b | 1.26 ± 0.01 b | 0.80 ± 0.01 c |
| HS + 1.0 | 56.1 ± 1.65 ab | 3.43 ± 0.12 d | 37.0 ± 0.70 b | 0.70 ± 0.04 b | 1.31 ± 0.02 c | 1.23 ± 0.03 c | 0.84 ± 0.02 b |
| T-effect | p < 0.001 | p = 0.066 | p = 0.689 | p = 0.109 | p < 0.001 | p < 0.001 | p < 0.001 |
| Proline (µ mol g−1 FW) | Soluble Sugar (mg g−1 FW) | Total Phenolic Content (mg g−1 FW) | Soluble Protein (mg g−1 FW) | CO2 Assimilation Rate (μmoL m−2 s−1) | Stomatal Conductance (moL m−2 s−1) | WUEi (μmoL m−1) | |
|---|---|---|---|---|---|---|---|
| CK | 15.8 ± 0.44 g | 70.6 ± 0.99 e | 1.46 ± 0.02 f | 25.3 ± 0.47 d | 10.0 ± 0.02 a | 0.54 ± 0.01 a | 18.5 ± 1.09 c |
| MS | 19.7 ± 0.97 f | 76.4 ± 0.65 d | 1.63 ± 0.01 e | 29.7 ± 1.21 c,d | 9.22 ± 0.02 b,c | 0.44 ± 0.02 b,c | 20.9 ± 1.31 b,c |
| HS | 25.3 ± 0.41 e | 80.2 ± 0.29 c | 1.81 ± 0.02 c | 33.4 ± 0.14 b,c | 8.34 ± 0.06 d | 0.36 ± 0.01 d | 23.1 ± 1.11 a |
| MS + 0.5 | 27.0 ± 0.77 d | 79.3 ± 0.47 c,d | 1.77 ± 0.03 d | 31.6 ± 0.81 c | 9.44 ± 0.09 a,b,c | 0.44 ± 0.03 b,c | 21.4 ± 1.22 a,b |
| HS + 0.5 | 30.6 ± 0.63 c | 83.2 ± 0.49 b | 1.98 ± 0.02 b | 35.2 ± 0.26 a,b,c | 8.65 ± 0.08 c,d | 0.37 ± 0.01 d | 23.3 ± 1.23 a |
| MS + 1.0 | 33.8 ± 0.60 b | 82.4 ± 0.41 b,c | 2.11 ± 0.04 a,b | 36.5 ± 0.37 a,b | 9.66 ± 0.09 a,b | 0.45 ± 0.02 b | 21.4 ± 1.12 a,b |
| HS + 1.0 | 35.4 ± 0.48 a | 85.4 ± 0.37 a | 2.28 ± 0.01 a | 37.4 ± 0.85 a | 8.71 ± 0.01 c | 0.39 ± 0.01 c | 22.3 ± 1.18 a,b |
| T-effect | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zafar, Z.; Rasheed, F.; Mushtaq, N.; Khan, M.U.; Mohsin, M.; Irshad, M.A.; Summer, M.; Raza, Z.; Gailing, O. Exogenous Application of Salicylic Acid Improves Physiological and Biochemical Attributes of Morus alba Saplings under Soil Water Deficit. Forests 2023, 14, 236. https://doi.org/10.3390/f14020236
Zafar Z, Rasheed F, Mushtaq N, Khan MU, Mohsin M, Irshad MA, Summer M, Raza Z, Gailing O. Exogenous Application of Salicylic Acid Improves Physiological and Biochemical Attributes of Morus alba Saplings under Soil Water Deficit. Forests. 2023; 14(2):236. https://doi.org/10.3390/f14020236
Chicago/Turabian StyleZafar, Zikria, Fahad Rasheed, Naveed Mushtaq, Muhammad Usman Khan, Muhammad Mohsin, Muhammad Atif Irshad, Muhammad Summer, Zohaib Raza, and Oliver Gailing. 2023. "Exogenous Application of Salicylic Acid Improves Physiological and Biochemical Attributes of Morus alba Saplings under Soil Water Deficit" Forests 14, no. 2: 236. https://doi.org/10.3390/f14020236
APA StyleZafar, Z., Rasheed, F., Mushtaq, N., Khan, M. U., Mohsin, M., Irshad, M. A., Summer, M., Raza, Z., & Gailing, O. (2023). Exogenous Application of Salicylic Acid Improves Physiological and Biochemical Attributes of Morus alba Saplings under Soil Water Deficit. Forests, 14(2), 236. https://doi.org/10.3390/f14020236