
Infestation Phases and Impacts of Dryocoetes confusus in Subalpine Fir Forests of Southern British Columbia


Abstract
:1. Introduction
- Stand and tree attributes of early-, mid-, and late-phase stands impacted by D. confusus;
- The progression of D. confusus attack and subalpine fir mortality in stands;
- The spatial-temporal distribution of live subalpine fir, other tree species, and D. confusus attacked subalpine fir.
2. Methods
2.1. Plot Selection and Establishment
- 70 mm photography of potential stands, carried out in 1997;
- Ground confirmation of D. confusus occurrence;
- The availability of ground access;
- Undisturbed stands with no plans for imminent harvest;
- A final rotary-wing reconnaissance flight to evaluate stand suitability.
- Six plots in the ESSFwc. The ESSFwc (wet, cold) rarely shows moisture deficits due to late snowmelt and frequent summer storms;
- Three plots in the ESSFxc. The ESSFxc (very dry, cold) is the driest and highest-elevation forested ESSF subzone in the southern interior. Snowpack rarely exceeds 80–100 cm, and frost can occur throughout the summer;
- Two plots in the ESSFmw. The ESSFmw (moist, warm) is the warmest ESSF subzone in the southern interior of British Columbia. The temperature regime is moderated by its proximity to the Pacific Ocean. This contributes to substantial winter precipitation and deep snowpack.
- Early-phase: new or building populations of D. confusus, characterized by scattered trees with bright red foliage (recent attack) and a low number of dead grey trees or old snags. The stands are typically of a higher density, with a closed canopy and minimal evidence of windthrow;
- Mid-phase: characterized by moderate to high levels of trees with bright red foliage (recent attack) and moderate levels of grey trees and snags;
- Late-phase: characterized by a few recent attacks (trees with bright red foliage) and high numbers of grey trees and snags. Stands typically have lower-density open canopies, with evidence of windthrow or fall-down.
2.2. Tree Assessment
2.3. Volume Estimation
- Height is tree height (m),
- DBH is tree diameter at breast height (cm),
- b0, b1, and b2 are regression coefficients,
- e is the base of the natural logarithm (2.71828) or Euler’s number.
2.4. Statistical Analysis
2.5. Temporal Patterns of Change in Stands
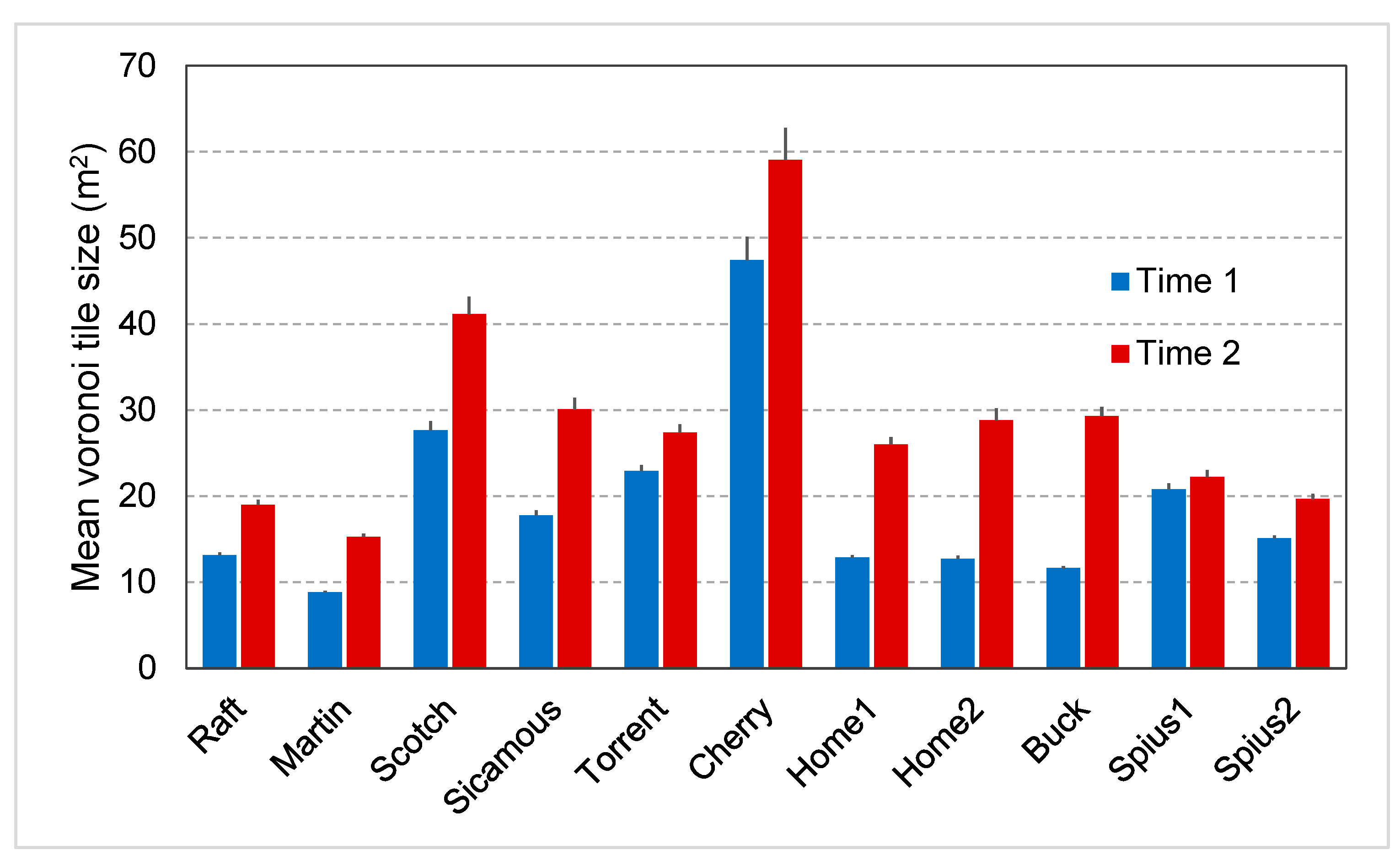
2.6. Spatial Analyses
3. Results
3.1. Infestation Progress
- Broken tops, affecting about 10% of subalpine fir;
- Two-year cycle budworm, Choristoneura biennis Freeman (Lepidoptera: Tortricidae), which, in some years, damaged most of the subalpine fir and spruce in a plot;
- Balsam bark weevil, P. striatulus, which affected about 2.6% of subalpine fir;
- Low levels of various secondary bark beetles;
- Pineus abietinus Underwood & Balch (Hemiptera; Adelgidae);
- Decay and rot (including Armillaria root disease);
- Windthrow, forking, and damage from other environmental factors, including frost, snow, ice, and lightning;
- Mechanical scarring from other trees;
- Localized animal damage caused by rodents feeding at the base of trees or scarring from bears.
3.2. Dryocoetes confusus Development
3.3. Stand Decline and Impact
3.4. Spatial Analyses
4. Discussion and Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, J.M.; Paritsis, J.; Veblen, T.T.; Chapman, T.B. Permanent forest plots show accelerating tree mortality in subalpine forests of the Colorado Front Range from 1982 to 2013. For. Ecol. Manag. 2015, 341, 8–17. [Google Scholar] [CrossRef]
- Maclauchlan, L.E. Quantification of Dryocoetes confusus caused mortality in subalpine fir forests of southern British Columbia. For. Ecol. Manag. 2016, 359, 210–220. [Google Scholar] [CrossRef]
- Reich, R.M.; Lundquist, J.E.; Hughes, K. Host-environment mismatches associated with subalpine fir decline in Colorado. Can. J. For. Res. 2016, 27, 1177–1189. [Google Scholar] [CrossRef]
- Lalande, B.M.; Hughes, K.; Jacobi, W.R.; Tinkham, W.T.; Reich, R.; Stewart, J.E. Subalpine fir mortality in Colorado is associated with stand density, warming climates and interactions among fungal diseases and the western balsam bark beetle. For. Ecol. Manag. 2020, 466, 118–133. [Google Scholar] [CrossRef]
- Howe, M.; Peng, L.; Carroll, A. Landscape predictions of western balsam bark beetle activity implicate warm temperatures, a longer growing season, and drought in widespread irruptions across British Columbia. For. Ecol. Manag. 2022, 508, 16. [Google Scholar] [CrossRef]
- Sinclair, W.A.; Hudler, G.W. Tree declines: Four concepts of causality. J. Arboric. 1988, 14, 29–35. [Google Scholar] [CrossRef]
- Furniss, R.L.; Carolin, V.M. Western Forest Insects; Misc. Pub. No. 1339; USDA Forest Service: Washington, DC, USA, 1977; p. 654. [CrossRef]
- Stock, A.J. The Western Balsam Bark Beetle, Dryocoetes confusus Swaine: Impact and Semiochemical-Based Management. Ph.D. Thesis, Simon Fraser University, Burnaby, BC, Canada, 1991. [Google Scholar]
- Garbutt, R. Western balsam bark beetle. Forest Pest Leaflet 64; Canadian Forest Service, Pacific Forestry Centre: Victoria, BC, Canada, 1992; p. 4.
- Bleiker, K.P.; Lindgren, B.S.; Maclauchlan, L.E. Characteristics of subalpine fir susceptible to attack by western balsam bark beetle (Coleoptera: Scolytidae). Can. J. For. Res. 2003, 33, 1538–1543. [Google Scholar] [CrossRef]
- McMillin, J.D.; Allen, K.K.; Long, D.F.; Harris, J.L.; Negrón, J.F. Effects of western balsam bark beetle on spruce-fir forests of north-central Wyoming. West. J. Appl. For. 2003, 184, 259–266. [Google Scholar] [CrossRef]
- Negrón, J.E.; Popp, J.B. The flight periodicity, attack patterns, and life history of Dryocoetes confusus Swaine (Coleoptera: Curculionidae: Scolytinae), the western balsam bark beetle, in North Central Colorado. West N. Am. Nat. 2009, 69, 447–458. [Google Scholar] [CrossRef]
- Maclauchlan, L.E.; Boone, C.; Robert, J. Declining Forests and a Forgotten Bark Beetle. In BC Forest Professional Magazine, Fall 2022; Association of BC Forest Professionals: Vancouver, BC, Canada, 2022. [Google Scholar]
- Winkler, R.; Spittlehouse, D.; Boon, S. Streamflow response to clear-cut logging on British Columbia’s Okanagan Plateau. Ecohydrology 2017, 10, 1836. [Google Scholar] [CrossRef]
- Van Mantgem, P.J.; Stephenson, N.L.; Byrne, J.C.; Daniels, L.D.; Franklin, J.F.; Fulé, P.Z.; Harmon, M.E.; Larson, A.J.; Smith, J.M.; Taylor, A.H.; et al. Widespread increase of tree mortality rates in the western United States. Science 2009, 323, 521–524. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Hicke, J.A.; Fisher, R.A.; Allen, C.D.; Aukema, J.; Bentz, B.; Hood, S.; Lichstein, J.W.; Macalady, A.K.; McDowell, N.; et al. Tree mortality from drought, insects, and their interactions in a changing climate. New Phytol. 2015, 208, 674–683. [Google Scholar] [CrossRef]
- Lloyd, D.; Angove, K.; Hope, G.; Thompson, C. A guide to site identification and interpretation for the Kamloops Forest Region. In BC Ministry of Forests Land Management Handbook 23 in 2 Parts; BC Ministry of Forests: Victoria, BC, Canada, 1990; p. 400. [Google Scholar]
- Meidinger, D.; Pojar, J. Ecosystems of British Columbia; Special Report Series No. 6; BC Ministry of Forests: Victoria, BC, Canada, 1991; p. 330. Available online: https://www.for.gov.bc.ca/hfd/pubs/docs/srs/srs06.htm (accessed on 1 December 2022).
- Parish, R.; Thomson, S. Tree Book: Learning to Recognize Trees of British Columbia; Natural Resources Canada, Canadian Forest Service, Pacific Forestry Centre: Victoria, BC, Canada; BC Ministry of Forests: Victoria, BC, Canada, 1994; p. 183.
- Klinka, K.; Worrall, J.J.; Skoda, L.; Varga, P. The Distribution and Synopsis of Ecological and Silvical Characteristics of Tree Species of British Columbia’s Forests; Canadian Cartographics Ltd.: Coquitlam, BC, Canada, 2000; p. 2. [Google Scholar] [CrossRef]
- Coupé, R.A.; Stewart, A.C.; Wikeem, B.M. Engelmann Spruce—Subalpine Fir Zone. In Ecosystems of British Columbia; Special Report Series 6; Meidinger, D.V., Pojar, J., Eds.; BC Ministry of Forests, Research Branch: Victoria, BC, Canada, 1991; pp. 223–236. [Google Scholar]
- DeLong, S.C.; Meidinger, D. Ecological variability of high elevation forests in central British Columbia. For. Chron. 2003, 79, 259–262. [Google Scholar] [CrossRef]
- Price, D.T.; Alfaro, R.I.; Brown, K.J.; Flannigan, M.D.; Fleming, R.A.; Hogg, E.H.; Girardin, M.P.; Lakusta, T.; Johnston, M.; McKenney, D.W.; et al. Anticipating the consequences of climate change for Canada’s boreal forest ecosystems. Environ. Rev. 2013, 21, 322–365. [Google Scholar] [CrossRef]
- Woods, A.J.; Coates, K.D.; Watts, M.; Foord, V.; Holtzman, E.I. Warning signals of adverse interactions between climate change and native stressors in British Columbia forests. Forests 2017, 8, 280. [Google Scholar] [CrossRef]
- Zhi, W.; Williams, K.H.; Carroll, R.W.; Brown, W.; Dong, W.; Kerins, D.; Li, L. Significant stream chemistry response to temperature variations in a high-elevation mountain watershed. Commun. Earth Environ. 2020, 1, 1–10. [Google Scholar] [CrossRef]
- Alexander, R.R. Ecology, silviculture, and management of the Engelmann spruce-subalpine fir type in the central and southern Rocky Mountains. In Agriculture Handbook 659; USDA Forest Service: Washington, DC, USA, 1987; p. 144. [Google Scholar]
- Griesbauer, H.; DeLong, S.C.; Rogers, B.; Foord, V. Growth sensitivity to climate varies with soil moisture regime in spruce–fir forests in central British Columbia. Trees 2021, 35, 649–669. [Google Scholar] [CrossRef]
- Peng, C.; Ma, Z.; Lei, X.; Zhu, Q.; Chen, H.; Wang, W.; Liu, S.; Li, W.; Fang, X.; Zhou, X. A drought-induced pervasive increase in tree mortality across Canada’s boreal forest. Nat. Clim. Change. 2011, 1, 467–471. [Google Scholar] [CrossRef]
- Williams, A.P.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D.; et al. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Change 2013, 3, 292–297. [Google Scholar] [CrossRef]
- Bentz, B.J.; Régnière, J.; Fettig, C.J.; Matthew, E.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.G.; Negrón, J.F.; Seybold, S.J. Climate change and bark beetles of the western United States and Canada: Direct and indirect effects. BioScience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Safranyik, L.; Wilson, B. The Mountain Pine Beetle: A Synthesis of Biology, Management, and Impacts on Lodgepole Pine; Natural Resources Canada, Canadian Forest Service, Pacific Forestry Centre: Victoria, BC, Canada, 2006; p. 304.
- Hlásny, T.; Krokene, P.; Liebhold, A.; Montagné-Huck, C.; Müller, J.; Qin, H.; Raffa, K.; Schelhaas, M.-J.; Seidl, R.; Svoboda, M.; et al. Living with bark beetles: Impacts, outlook and management options. In From Science to Policy 8; European Forest Institute: Joensuu, Finland, 2019; p. 52. [Google Scholar] [CrossRef]
- Kurz, W.A.; Dymond, C.C.; Stinson, G.; Rampley, G.J.; Neilson, E.T.; Carroll, A.L.; Ebata, T.; Safranyik, L. Mountain pine beetle and forest carbon feedback to climate change. Nature 2008, 452, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Safranyik, L.; Carroll, A.L.; Régnière, J.; Langor, D.W.; Riel, W.G.; Shore, T.L.; Peter, B.; Cooke, B.J.; Nealis, V.G.; Taylor, S.W. Potential for range expansion of mountain pine beetle into the boreal forest of North America. Can. Entomol. 2010, 142, 415–442. [Google Scholar] [CrossRef]
- Maclauchlan, L.E.; Brooks, J.E.; White, K.J. Impacts and susceptibility of young pine stands to the mountain pine beetle, Dendroctonus ponderosae, in British Columbia. J. Ecosys. Manage. 2015, 15, 1–18. [Google Scholar] [CrossRef]
- Maclauchlan, L.E.; Buxton, K. 2018 Overview of Forest Health Conditions in Southern British Columbia; Ministry of Forests, Range and Natural Resource Operations and Rural Development, Thopson Okanagan Region: Kamloops, BC, Canada, 2019; p. 73. Available online: https://www2.gov.bc.ca/assets/gov/environment/research–monitoring-and-reporting/monitoring/aerial-overview-survey-documents/2018_south_overview_report_web.pdf (accessed on 2 November 2022).
- Fettig, C.J.; Progar, R.A.; Paschke, J.; Sapio, F.J. Forest insects. In Disturbance and Sustainability in Forests of the Western United States; Robertson, G., Barrett, T., Eds.; Gen. Tech. Rep. PNW-GTR-992; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2021; pp. 81–121. [Google Scholar]
- Robbins, Z.J.; Xu, C.; Aukema, B.H.; Buotte, P.C.; Chitra-Tarak, R.; Fettig, C.J.; Goulden, M.L.; Goodsman, D.W.; Hall, A.D.; Koven, C.D.; et al. Warming increased bark beetle-induced tree mortality by 30% during an extreme drought in California. Global Change Biol. 2022, 28, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Maclauchlan, L.E. 1999 Overview of Forest Health in the Kamloops Forest Region; Ministry of Forests: Kamloops, BC, Canada, 2000; p. 23.
- Westfall, J.M.; Ebata, T. 2018 Summary of Forest Health Conditions in British Columbia; BC Ministry of Forests, Lands and Natural Resource Operations: Victoria, BC, Canada, 2019; p. 95.
- Stock, A.J.; Borden, J.H.; Pratt, T.L. Containment and concentration of infestations of the western balsam bark beetle, Dryocoetes confusus (Coleoptera: Scolytidae), using the aggregation pheromone exo-brevicomin. Can. J. For. Res. 1994, 24, 483–492. [Google Scholar] [CrossRef]
- Maclauchlan, L.E.; Brooks, J.E. Western balsam bark beetle, Dryocoetes confusus Swaine (Coleoptera: Curculionidae: Scolytinae), in situ development and seasonal flight periodicity in southern British Columbia. J. Entomol. Soc. B. C. 2017, 114, 1–16. [Google Scholar]
- Maclauchlan, L.E.; Brooks, J.E. Comparison of two treatment regimes for managing western balsam bark beetle. J. Ecosys. Manage. 2021, 21, 1–11. [Google Scholar] [CrossRef]
- Mathers, W.G. The biology of Canadian bark beetles: The seasonal history of Dryocoetes confusus Sw. Can. Entomol. 1931, 63, 247–248. [Google Scholar] [CrossRef]
- Gaylord, M.L.; Williams, K.K.; Hofstetter, R.W.; McMillin, J.D.; DeGomez, T.E.; Wagner, M.R. Influence of temperature on spring flight initiation for southwestern ponderosa pine bark beetles (Coleoptera: Curculionidae, Scolytinae). Environ. Entomol. 2008, 37, 57–69. [Google Scholar] [CrossRef]
- Hansen, E.M. Western balsam bark beetle, Dryocoetes confusus Swaine, flight periodicity in northern Utah. Great Basin Naturalist. 1996, 56, 348–359. [Google Scholar]
- Gibson, K.; Kegley, S.; Oakes, B. Western Balsam Bark Beetle Activity and Flight Periodicity in the Northern Region; Rep. 97-3; USDA Forest Service, Cooperative Forestry and Forest Health Protection: Missoula, MT, USA, 1997; p. 5.
- McMillin, J.D.; Allen, K.K.; Long, D.F. Evaluation of Western Balsam Bark Beetle Flight Periodicity on the Bighorn National Forest, Wyoming; Tech. Rep. R2-65; USDA Forest Service Rocky Mountain Region: Golden, CO, USA, 2001; p. 19.
- Stock, A.J.; Pratt, T.L.; Borden, J.H. Seasonal flight pattern of the western balsam bark beetle, Dryocoetes confusus Swaine (Coleoptera: Curculionidae), in central British Columbia. J. Entomol. Soc. B. C. 2013, 110, 27–34. [Google Scholar]
- Molnar, A.C. Pathogenic fungi associated with a bark beetle on alpine fir. Can. J. Bot. 1965, 43, 39–43. [Google Scholar] [CrossRef]
- Mattson, W.; Haack, R.A. Role of drought in outbreaks of plant-eating insects. BioScience 1987, 37, 110–118. [Google Scholar] [CrossRef]
- Bright, D.E., Jr. Bark beetles of the genus Dryocoetes (Coleoptera: Scolytidae) in North America. Ann. Entomol. Soc. Am. 1963, 56, 103–115. [Google Scholar] [CrossRef]
- Hansen, E.M.; Bentz, B.J. Comparison of reproductive capacity among univoltine, semivoltine, and re-emerged parent spruce beetles. Can. Entomol. 2003, 135, 697–712. [Google Scholar] [CrossRef]
- Bleiker, K.P.; Willsey, T. Experimental evidence supporting an obligate adult diapause for spruce beetle (Coleoptera: Curculionidae) from British Columbia. Environ. Entomol. 2020, 49, 98–103. [Google Scholar] [CrossRef]
- Antos, J.A.; Parish, R. Structure and dynamics of a nearly steady-state subalpine forest in south-central British Columbia, Canada. Oecologia 2002, 130, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Harvey, B.J.; Andrus, R.A.; Battaglia, M.A.; Negrón, J.F.; Orrego, A.; Veblen, T.T. Droughty times in mesic places: Factors associated with forest mortality vary by scale in a temperate subalpine region. Ecosphere 2021, 12, e03318. [Google Scholar] [CrossRef]
- Bleiker, K.P.; Lindgren, B.S.; Maclauchlan, L.E. Resistance of fast- and slow-growing subalpine fir to pheromone-induced attack by western balsam bark beetle (Coleoptera: Scolytinae). Agric. For. Entomol. 2005, 7, 237–244. [Google Scholar] [CrossRef]
- Maclauchlan, L.E.; Harder, L.; Borden, J.H.; Brooks, J.E. Impact of the western balsam bark beetle, Dryocoetes confusus Swaine (Coleoptera: Scolytidae), at the Sicamous Research site, and the potential for semiochemical based management in alternative silviculture systems. J. Entomol. Soc. B. C. 2003, 100, 27–41. [Google Scholar]
- Chapman, D.G. Statistical problems in population dynamics. In Proceedings of the Fourth Berkeley Symposium on Mathematical Statistics and Probability; Neyman, J., Ed.; University of California Press: Berkeley, CA, USA, 1961; Volume 4, pp. 153–186. [Google Scholar]
- R Core Team. R: A language and environment for statistical computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 1 December 2022).
- Kozak, A. My last words on taper equations. For. Chron. 2004, 80, 507–515. [Google Scholar] [CrossRef] [Green Version]
- Zar, J.H. Biostatistical Analysis, 2nd ed.; Prentice-Hall, Inc.: Englewood Cliffs, NJ, USA, 1984; p. 718. [Google Scholar]
- Churchill, D.J.; Larson, A.J.; Jeronimo, S.M.A.; Dalhgreen, M.C.; Franklin, J.F. The ICO Approach to Quantifying and Restoring Forest Spatial Pattern: Implementation Guide, Version 2.0.; Stewardship Forestry: Vashon, WA, USA, 2013; p. 68. [Google Scholar]
- Churchill, D.J.; Carnwath, G.C.; Larson, A.J.; Jeronimo, S.A. Historical Forest Structure, Composition, and Spatial Pattern in Dry Conifer Forests of the Western Blue Mountains, Oregon; Gen. Tech. Rep. PNW-GTR-956; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2017; p. 93. [CrossRef]
- Turner, R. _deldir: Delaunay Triangulation and Dirichlet (Voronoi) Tessellation. R package Ver. 1.0-6. 2021. Available online: https://CRAN.R-project.org/package=deldir (accessed on 1 December 2022).
- Brown, G.S. Point density in stems per acre. Forest Research Institute, New Zealand Forest Service. N. Z. For. Res. Notes 1965, 196, 11. [Google Scholar]
- Maclauchlan, L.E.; Borden, J.H. Spatial dynamics and impacts of Pissodes terminalis (Coleoptera: Curculionidae) in regenerating stands of lodgepole pine. For. Ecol. Manag. 1996, 82, 103–113. [Google Scholar] [CrossRef]
- Baddeley, A.; Rubak, E.; Turner, R. Spatial Point Patterns: Methodology and Applications with R; Chapman and Hall/CRC Press: London, UK, 2015. [Google Scholar]
- Statistics, version 7.0; SYSTAT® for Windows; SPSS Inc.: Chicago, IL, USA, 1997.
- Diggle, P.J. Statistical Analysis of Spatial Point Patterns; Oxford University Press: New York, NY, USA, 2003; p. 159. [Google Scholar]
- Lydersen, J.M.; North, M.P.; Knapp, E.E.; Collins, B.M. Quantifying spatial patterns of tree groups and gaps in mixed-conifer forests: Reference conditions and long-term changes following fire suppression and logging. For. Ecol. Manag. 2013, 304, 370–382. [Google Scholar] [CrossRef]
- Plotkin, J.B.; Chave, J.; Ashton, P.S. Cluster analysis of spatial patterns in Malaysian tree species. Am. Nat. 2002, 160, 629–644. [Google Scholar] [CrossRef]
- Larson, A.J.; Churchill, D.J. Spatial patterns of overstory trees in late successional conifer forests. Can. J. For. Res. 2008, 38, 2814–2825. [Google Scholar] [CrossRef]
- Larson, A.J.; Stover, K.C.; Keyes, C.R. Effects of restoration thinning on spatial heterogeneity in mixed conifer forest. Can. J. For. Res. 2012, 42, 1505–1517. [Google Scholar] [CrossRef]
- Larson, A.J.; Churchill, D.J. Tree spatial patterns in fire-frequent forests of western North America, including mechanisms of pattern formation and implications for designing fuel reduction and restoration treatments. For. Ecol. Manag. 2012, 267, 74–92. [Google Scholar] [CrossRef]
- Abella, S.R.; Denton, C.W. Spatial variation in reference conditions: Historical tree density and pattern on a Pinus ponderosa landscape. Can. J. For. Res. 2009, 39, 2391–2403. [Google Scholar] [CrossRef]
- Maclauchlan, L.E.; Brooks, J.E. The balsam bark weevil, Pissodes striatulus (Coleoptera: Curculionidae): Life history and occurrence in southern British Columbia. J. Entomol. Soc. B. C. 2020, 117, 3–19. [Google Scholar]
- DeLong, S.C.; Sutherland, G.D.; Daniels, L.D.; Heemskerk, B.H.; Storaunet, K.O. Temporal dynamics of snags and development of snag habitats in wet spruce–fir stands in east-central British Columbia. For. Ecol. Manag. 2008, 255, 3613–3620. [Google Scholar] [CrossRef]
- Government Province of B.C. Ministry of Forests. Aerial Overview Survey Data. 2022. Available online: https://www2.gov.bc.ca/gov/content/industry/forestry/managing-our-forest-resources/forest-health/aerial-overview-surveys/data-files (accessed on 23 September 2022).
- Kenkel, N.C.; Hoskins, J.A.; Hoskins, W.D. Edge effects in the use of area polygons to study competition. Ecology 1989, 70, 272–274. [Google Scholar] [CrossRef]
- Hyink, D.M.; Zedaker, S.M. Stand dynamics and the evaluation of forest decline. Tree Physiol. 1987, 3, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Fettig, C.; Klepzig, K.; Billings, R.; Munson, S.; Nebeker, T.; Negrón, J.; Nowak, J. The effectiveness of vegetation management practices for prevention and control of bark beetle infestations in coniferous forests of the western and southern United States. For. Ecol. Manag. 2007, 238, 24–53. [Google Scholar] [CrossRef]
- Flower, C.E.; Gonzalez-Meler, M.A. Responses of temperate forest productivity to insect and pathogen disturbances. Ann. Rev. Plant Biol. 2015, 66, 547–569. [Google Scholar] [CrossRef]
- Seidl, R.; Spies, T.A.; Peterson, D.L.; Stephens, S.L.; Hicke, J.A. Searching for resilience: Addressing the impacts of changing disturbance regimes on forest ecosystem services. J. Appl. Ecol. 2016, 53, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Rodman, K.C.; Andrus, R.A.; Carlson, A.R.; Carter, T.A.; Chapman, T.B.; Coop, J.D.; Fornwalt, P.J.; Gill, N.S.; Harvey, B.J.; Hoffman, A.E.; et al. Rocky Mountain forests are poised to recover following bark beetle outbreaks, but with altered composition. J. Ecol. 2022, 110, 2929–2949. [Google Scholar] [CrossRef]
- Lertzmen, K.P.; Krebs, C.J. Gap-phase structure of a subalpine old-growth forest. Can. J. For. Res. 1991, 21, 1730–1741. [Google Scholar] [CrossRef]
- Franklin, J.F.; Van Pelt, R. Spatial aspects of structural complexity in old-growth forests. J. For. 2004, 102, 22–28. [Google Scholar] [CrossRef]
- Boyden, S.; Binkley, D.; Shepperd, W. Spatial patterns in structure, regeneration and mortality of an old-growth ponderosa pine forest in the Colorado Front Range. For. Ecol. Manag. 2005, 219, 43–55. [Google Scholar] [CrossRef]
- Schneider, E.E.; Larson, A.J. Spatial aspects of structural complexity in Sitka spruce-western hemlock forests, including evaluation of a new canopy gap delineation method. Can. J. For. Res. 2017, 47, 1033–1044. [Google Scholar] [CrossRef]
- Purves, D.W.; Lichstein, J.W.; Pacala, S.W. Crown plasticity and competition for canopy space: A new spatially implicit model parameterized for 250 North American tree species. PLoS ONE 2007, 2, e870. [Google Scholar] [CrossRef]
- Thorpe, H.C.; Astrup, R.; Trowbridge, A.; Coates, K.D. Competition and tree crowns: A neighbourhood analysis of three boreal tree species. For. Ecol. Manag. 2010, 259, 1586–1596. [Google Scholar] [CrossRef]
- Canham, C.D.; LePage, P.T.; Coates, K.D. A neighbourhood analysis of canopy tree competition: Effects of shading versus crowding. Can. J. For. Res. 2004, 34, 778–787. [Google Scholar] [CrossRef]
- Alexander, R.R.; Shearer, R.C.; Shepperd, W.D. Silvical Characteristics of Subalpine Fir; Gen. Tech. Rep. RM-115; USDA Forest Service, Rocky Mountain Forest and Range Experimental Station: Fort Collins, CO, USA, 1984; p. 29. [CrossRef]
- Sánchez Meador, A.J.; Parysow, P.F.; Moore, M.M. A new method for delineating tree patches and assessing spatial reference conditions of ponderosa pine forests in northern Arizona. Restor. Ecol. 2011, 19, 490–499. [Google Scholar] [CrossRef]
- Parish, R.; Antos, J.A. Dynamics of an old-growth, fire-initiated, subalpine forest in southern interior British Columbia: Tree-ring reconstruction of 2 year cycle spruce budworm outbreaks. Can. J. For. Res. 2002, 38, 1947–1960. [Google Scholar] [CrossRef]
- Langs, L.E.; Petrone, R.M.; Pomeray, J.W. Subalpine forest water use behaviour and evapotranspiration during two hydrologically contrasting growing seasons in the Canadian Rockies. Hydrol. Proc. 2021, 35, e14158. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot No. and Location | Latitude | Longitude | BEC 1 Unit | Elevation (m) | Infestation Phase 2 | Establishment and Assessments (Years) 3 |
|---|---|---|---|---|---|---|
| 1. Raft River | 51°52’57.62” N | 119°33’11.35” W | ESSFwc | 1503 | Early to mid- | 2012, 2019 |
| 2. Martin Creek | 51°44’15.43” N | 119°39’0.20” W | ESSFwc | 1675 | Early | 2000–2004, 2009, 2013, 2019 * |
| 3. Scotch Creek | 51°9’58.52” N | 119°12’26.91” W | ESSFwc | 1575 | Mid- to late | 2002–2004, 2011, 2020 |
| 4. Sicamous Creek | 50°49’47.13” N | 118°49’51.19” W | ESSFwc | 1650 | Late | 1998–2004, 2009, 2013, 2019 |
| 5. Torrent Creek | 50°34’26.81” N | 118°34’9.97” W | ESSFwc | 1750 | Mid- to late | 1998–2004, 2008, 2013, 2019 |
| 6. Cherry Creek | 50°18’33.51” N | 118°29’8.74” W | ESSFwc | 1650 | Late | 1998–2004, 2009, 2013, 2019 |
| 7. Home Lake-1 | 50°4’0.28” N | 118°48’47.61” W | ESSFxc | 1800 | Mid- | 1999–2004, 2008, 2013 * |
| 8. Home Lake-2 | 50°5’57.15” N | 118°46’52.55” W | ESSFxc | 1750 | Mid- | 1999–2004, 2008, 2013, 2020 * |
| 9. Buck Mountain | 50°5’21.63” N | 118°52’55.69” W | ESSFxc | 1725 | Early to mid- | 1999–2004, 2009, 2013, 2020 |
| 10. Spius Creek-1 | 49°58’42.76” N | 121°10’35.36” W | ESSFmw | 1470 | Mid- | 2002–2004, 2008, 2013, 2019 |
| 11. Spius Creek-2 | 49°59’11.99” N | 121°8’39.54” W | ESSFmw | 1610 | Early | 2002–2004, 2008, 2013, 2019 |
| Tree Code | Tree Code Description |
|---|---|
| 0 | healthy, standing live tree |
| 1 | green foliage with current D. confusus attack |
| 2 | brick red foliage |
| 3 | faded, dull red foliage |
| 4 | grey with fine branches and a few red needles remaining on tree |
| 5 | grey without fine branches, just larger branches still intact |
| 6 | snag—losing bark, sapwood checking |
| 7 | dead—other damage agent or unknown cause |
| 10 | windthrow—previously attacked by D. confusus |
| 11 | windthrow—no D. confusus attack prior to falling |
| At Establishment | % Bl Dead (All Causes) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Plot | 1st to Final Assessment (# Years) | Year Cores Collected | No. Cores (Bl) | Avg. Age Bl (± S.E.) | Bl Age Range | Avg. Age Sx (± S.E.) and (N) | Bl (# Live & Dead) | Bl & Sx (# Live & Dead) | Time 1 | Time 2 |
| 1. Raft River | 7 | 2012 | 25 | 149 ± 10 | 97–248 | 273 ± 43 (5) | 570 | 870 | 24.9 | 60.0 |
| 2. Martin Creek | 19 | 2001 | 44 | 89 ± 3 | 74–164 | 85 ± 4 (10) | 1161 | 1417 | 25.0 | 71.0 |
| 3. Scotch Creek | 18 | 2003 | 26 | 114 ± 3 | 83–138 | 115 ± 6 (8) | 665 | 724 | 50.5 | 80.2 |
| 4. Sicamous Creek | 21 | 2001 | 21 | 136 ± 7 | 98–201 | 123 (1) | 732 | 930 | 50.1 | 78.4 |
| 5. Torrent Creek | 21 | 2013 | 19 | 126 ± 11 | 67–243 | 101 ± 2 (4) | 514 | 597 | 30.7 | 47.3 |
| 6. Cherry Creek | 21 | 2001 | 28 | 124 ± 5 | 71–209 | 197 ± 45 (4) | 425 | 496 | 58.6 | 70.2 |
| 7. Home Lake-1 | 14 | 2001 | 41 | 148 ± 5 | 95–259 | 995 | 1202 | 39.1 | 79.8 | |
| 8. Home Lake-2 | 14 | 2001 | 29 | 127 ± 4 | 82–212 | 155 ± 15 (9) | 1149 | 1313 | 44.1 | 79.0 |
| 9. Buck Mountain | 21 | 2001 | 32 | 122 ± 3 | 88–164 | 1215 | 1317 | 38.4 | 79.1 | |
| 10. Spius Creek-1 | 17 | 2002 | 42 | 184 ± 5 | 121–284 | 179 ± 9 (13) | 614 | 785 | 39.7 | 48.1 |
| 11. Spius Creek-2 | 17 | 2002 | 6 | 180 ± 9 | 153–211 | 674 | 841 | 16.9 | 40.4 | |
| % Bl at Establishment | % Bl at Final Assessment | Annual Rate of Mortality (%) | |||||
|---|---|---|---|---|---|---|---|
| Plot 1 | Dead Due to D. confusus | Dead Other Causes | Live | Dead Due to D. confusus | Dead Other Causes | D. confusus | All Causes |
| 1. Raft River | 13.3 | 11.6 | 40.0 | 34.0 | 26.0 | 3.0 | 5.0 |
| 2. Martin Creek | 13.4 | 11.5 | 29.0 | 42.6 | 28.5 | 1.5 | 2.4 |
| 3. Scotch Creek | 27.2 | 23.2 | 19.8 | 42.1 | 38.1 | 0.8 | 1.7 |
| 4. Sicamous Creek | 33.3 | 16.8 | 21.6 | 52.0 | 26.4 | 0.9 | 1.3 |
| 5. Torrent Creek | 16.9 | 13.8 | 52.7 | 27.0 | 20.3 | 0.5 | 0.8 |
| 6. Cherry Creek | 50.0 | 8.8 | 29.8 | 54.3 | 16.0 | 0.2 | 0.5 |
| 7. Home Lake-1 1 | 20.9 | 18.2 | 20.2 | 53.9 | 26.0 | 2.4 | 2.9 |
| 8. Home Lake-2 1 | 22.8 | 21.2 | 21.3 | 48.6 | 30.6 | 3.5 | 2.2 |
| 9. Buck Mountain | 14.5 | 24.0 | 20.9 | 44.6 | 34.5 | 1.4 | 1.9 |
| 10. Spius Creek-1 | 28.0 | 11.7 | 51.9 | 33.1 | 15.0 | 0.3 | 0.5 |
| 11. Spius Creek-2 | 11.5 | 5.3 | 59.6 | 27.6 | 12.8 | 0.9 | 1.4 |
| Phase | % sph Live Bl | % sph D. confusus-Killed |
|---|---|---|
| Early | ~80+% | ~10% |
| Mid | ~50%–70% | ~15%–30% |
| Late | <50% | >30% |
| Attack to Snag | # Years Dead to Down (Avg. ± S.E.) | ||||
|---|---|---|---|---|---|
| Plot | N | # Years (Avg. ± S.E.) | Median | D. confusus | Other Factors |
| 1. Raft River | 9 | 7 | 7 | - | 7 |
| 2. Martin Creek | 98 | 12.7 ± 0.4 | 13 | 14.1 ± 0.5 | 11.4 ± 1.1 |
| 3. Scotch Creek | 22 | 10.2 ± 0.7 | 9 | 14.2 ± 0.8 | 12.0 ± 0.9 |
| 4. Sicamous Creek | 52 | 14.4 ± 0.5 | 16 | 15.2 ± 0.9 | 13.2 ± 1.4 |
| 5. Torrent Creek | 6 | 15 | 15 | 13.8 ± 1.9 | 14.3 ± 1.6 |
| 6. Cherry Creek | 31 | 13.3 ± 0.6 | 14 | 16.3 ± 1.1 | 12.3 ± 2.2 |
| 7. Home Lake-1 1 | 95 | 11.5 ± 0.2 | 12 | 11.2 ± 0.4 | 7.0 ± 6.0 |
| 8. Home Lake-2 1 | 182 | 12.4 ± 0.2 | 13 | 12.8 ± 0.4 | 9.1 ± 1.6 |
| 9. Buck Mountain | 99 | 10.4 ± 0.3 | 11 | 15.1 ± 0.4 | 14.7 ± 0.6 |
| 10. Spius Creek-1 | 16 | 16.5 ± 0.4 | 17 | 5.0 ± 0.0 | 15 |
| 11. Spius Creek-2 | 13 | 10.8 ± 0.8 | 11 | 12.8 ± 0.8 | 9.3 ± 1.8 |
| All plots combined | 623 | 12.1 ± 0.1 | 12 | 14.0 ± 0.2 | 12.8 ± 0.4 2 |
| Merchantable Volume (m3) | |||||||
|---|---|---|---|---|---|---|---|
| Live | Dead | D. confusus Killed | |||||
| Plot | Species | Time 1 | Time 2 | Cumulative (all Causes) | At Time 1 | Time 1 to Time 2 | Avg. vol./Year |
| 1. Raft River | Bl | 140.5 | 76.0 | 163.8 | 29.5 | 83.8 | 10.5 |
| 1. Raft River | Sx | 115.8 | 135.8 | 6.6 | |||
| 1. Raft River | other | 31.4 | 42.7 | 0.3 | |||
| 2. Martin Creek | Bl | 159.4 | 81.7 | 264.3 | 39.8 | 169.5 | 8.5 |
| 2. Martin Creek | Sx | 56.8 | 70.6 | 16.3 | |||
| 3. Scotch Creek | Bl | 152.9 | 87.1 | 376.3 | 94.5 | 153.4 | 8.1 |
| 3. Scotch Creek | Sx | 38.1 | 54.5 | 30.5 | |||
| 3. Scotch Creek | other | 0.1 | 0.7 | 0.6 | |||
| 4. Sicamous Creek | Bl | 179.9 | 107.7 | 410.6 | 149.6 | 175.6 | 8.0 |
| 4. Sicamous Creek | Sx | 95.1 | 155.4 | 13.6 | |||
| 5. Torrent Creek | Bl | 177.8 | 208.3 | 272.3 | 95.1 | 96.4 | 4.4 |
| 5. Torrent Creek | Sx | 49.9 | 73.2 | 14.9 | |||
| 6. Cherry Creek | Bl | 169.8 | 157.9 | 377.8 | 239.7 | 79.7 | 3.6 |
| 6. Cherry Creek | Sx | 62.0 | 91.2 | 18.0 | |||
| 7. Home Lake-1 | Bl | 205.0 | 58.1 | 354.8 | 50.2 | 235.8 | 15.7 |
| 7. Home Lake-1 | Sx | 136.9 | 158.2 | 15.9 | |||
| 8. Home Lake-2 | Bl | 190.7 | 63.7 | 344.8 | 74.5 | 193.4 | 12.9 |
| 8. Home Lake-2 | Sx | 99.0 | 25.9 | 20.1 | |||
| 9. Buck Mountain | Bl | 173.7 | 86.0 | 379.9 | 61.2 | 205.9 | 9.4 |
| 9. Buck Mountain | Sx | 99.9 | 111.3 | 34.6 | |||
| 10. Spius Creek-1 | Bl | 142.6 | 137.0 | 203.7 | 92.5 | 73.6 | 4.1 |
| 10. Spius Creek-1 | Sx | 93.2 | 118.1 | 85.6 | |||
| 10. Spius Creek-1 | other | 0.4 | 0.8 | 3.4 | |||
| 11. Spius Creek-2 | Bl | 203.5 | 182.1 | 214.2 | 30.7 | 133.9 | 7.4 |
| 11. Spius Creek-2 | Sx | 63.2 | 89.4 | 136.6 | |||
| 11. Spius Creek-2 | other | 1.6 | 2.7 | 7.8 | |||
| Bins (Meters) | |||||||
|---|---|---|---|---|---|---|---|
| Time | (1,3] | (3,5] | (5,7] | (7,9] | (9,11] | (11,13] | (13,15] |
| 1 | 71.31 | 23.25 | 4.4 | 0.82 | 0.17 | 0.04 | 0 |
| 2 | 56.98 | 31.27 | 9.1 | 2.15 | 0.38 | 0.09 | 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maclauchlan, L.E.; Stock, A.J.; Brooks, J.E. Infestation Phases and Impacts of Dryocoetes confusus in Subalpine Fir Forests of Southern British Columbia. Forests 2023, 14, 363. https://doi.org/10.3390/f14020363
Maclauchlan LE, Stock AJ, Brooks JE. Infestation Phases and Impacts of Dryocoetes confusus in Subalpine Fir Forests of Southern British Columbia. Forests. 2023; 14(2):363. https://doi.org/10.3390/f14020363
Chicago/Turabian StyleMaclauchlan, Lorraine E., Arthur J. Stock, and Julie E. Brooks. 2023. "Infestation Phases and Impacts of Dryocoetes confusus in Subalpine Fir Forests of Southern British Columbia" Forests 14, no. 2: 363. https://doi.org/10.3390/f14020363
APA StyleMaclauchlan, L. E., Stock, A. J., & Brooks, J. E. (2023). Infestation Phases and Impacts of Dryocoetes confusus in Subalpine Fir Forests of Southern British Columbia. Forests, 14(2), 363. https://doi.org/10.3390/f14020363