Pipe Model Can Accurately Estimate Crown Biomass of Larch (Larix olgensis) Plantation Forest in Northeast China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Data
2.2. Estimation of the Stem Cross-Sectional Area at the Crown Base (AB)
2.3. Estimation of Crown Biomass
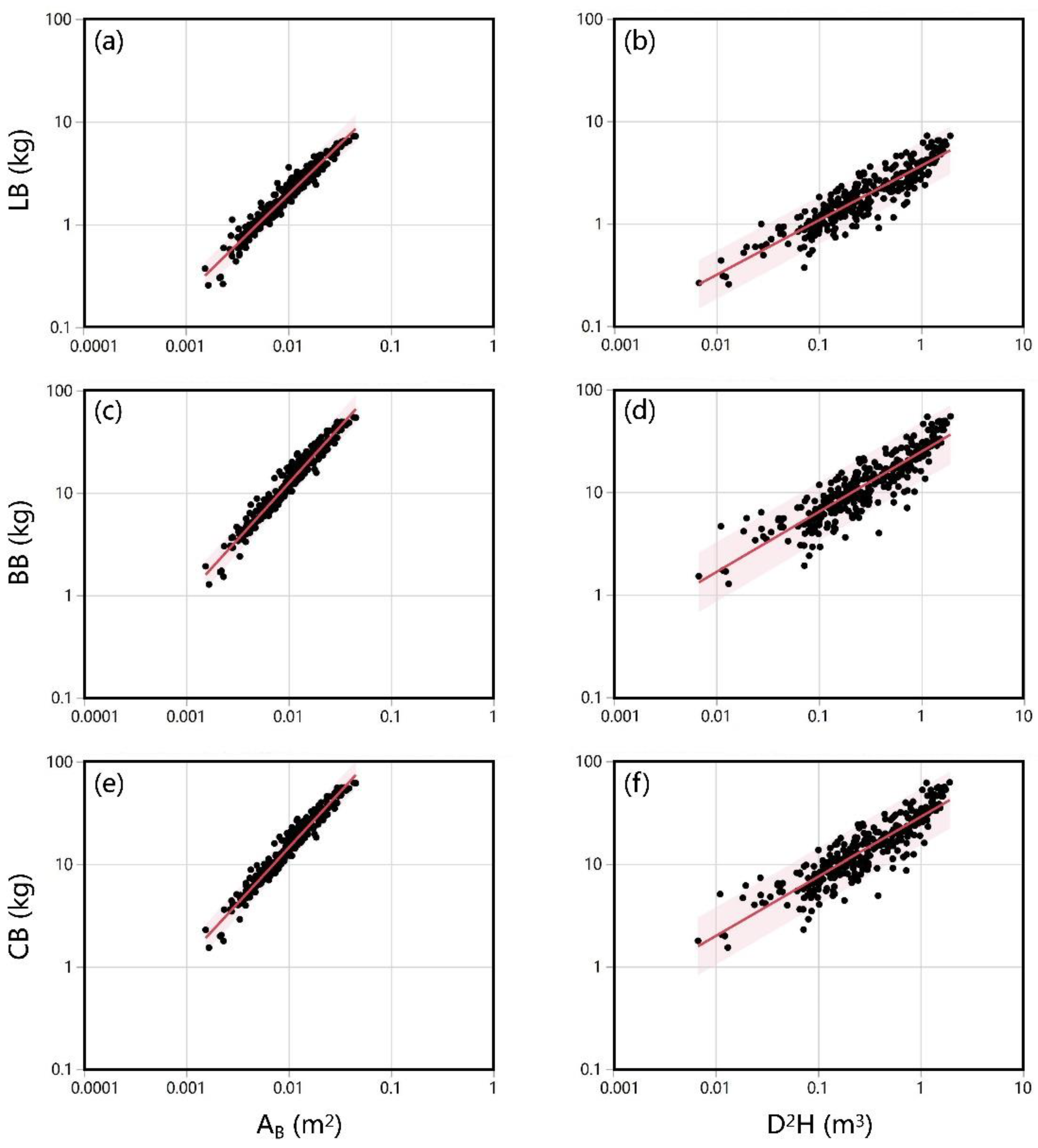
3. Results
3.1. Estimation of the Stem Cross-Sectional Area at the Crown Base (AB)
3.2. Estimation of Crown Biomass
4. Discussion
4.1. Estimation AB Based on the Pipe Model
4.2. Positive Proportional Relationship between Biomass and AB
4.3. Effects of Site, Stand Density and Age on Biomass
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zou, W.T.; Zeng, W.S.; Zhang, L.J.; Zeng, M. Modeling crown biomass for four pine species in China. Forests 2015, 6, 433–449. [Google Scholar] [CrossRef]
- Clark, D.A.; Brown, S.; Kicklighter, D.W.; Chambers, J.Q.; Thomlinson, J.R.; Ni, J. Measuring net primary production in forests: Concepts and field methods. Ecol. Appl. 2001, 11, 356–370. [Google Scholar] [CrossRef]
- Meng, S.; Yang, F.; Hu, S.; Wang, H.; Wang, H. Generic additive allometric models and biomass allocation for two natural oak species in northeastern China. Forests 2021, 12, 715. [Google Scholar] [CrossRef]
- Chen, D.; Li, F. The characteristics and biomass distribution in crown of Larix olgensis in Northeastern China. J. Korean Soc. For. Sci. 2010, 99, 204–212. [Google Scholar]
- Ogawa, K.; Adu-Bredu, S.; Yokota, T.; Hagihara, A. Leaf biomass changes with stand development in hinoki cypress (Chamaecyparis obtusa [Sieb. et Zucc.] Endl.). Plant Ecol. 2010, 211, 79–88. [Google Scholar] [CrossRef]
- Ochał, W.; Wertz, B.; Orzeł, S. Above-ground biomass allocation and potential carbon sink of black pine–a case study from southern Poland. Ann. For. Res. 2022, 65, 71–90. [Google Scholar]
- Dong, L.; Zhang, L.; Li, F. Additive biomass equations based on different dendrometric variables for two dominant species (Larix gmelini Rupr. and Betula platyphylla Suk.) in natural forests in the Eastern Daxing’an Mountains, Northeast China. Forests 2018, 9, 261. [Google Scholar] [CrossRef]
- Huxley, J.S. Problems of relative growth (Methuen & Co. Ltd, London). Nature 1932, 129, 775–777. [Google Scholar]
- Zianis, D.; Radoglou, K. Comparison between empirical and theoretical biomass allometric models and statistical implications for stem volume predictions. Forestry 2006, 79, 477–487. [Google Scholar] [CrossRef]
- Dutcă, I.; McRoberts, R.E.; Naesset, E.; Blujdea, V.N. A practical measure for determining if diameter (D) and height (H) should be combined into D2H in allometric biomass models. For. Int. J. For. Res. 2019, 92, 627–634. [Google Scholar] [CrossRef]
- Henry, M.; Picard, N.; Trotta, C.; Manlay, R.; Valentini, R.; Bernoux, M.; Saint-André, L. Estimating tree biomass of sub-Saharan African forests: A review of available allometric equations. Silva Fenn. 2011, 45, 477–569. [Google Scholar] [CrossRef]
- Hosoda, K.; Iehara, T. Aboveground biomass equations for individual trees of Cryptomeria japonica, Chamaecyparis obtusa and Larix kaempferi in Japan. J. For. Res. 2010, 15, 299–306. [Google Scholar] [CrossRef]
- Pérez-Cruzado, C.; Rodríguez-Soalleiro, R. Improvement in accuracy of aboveground biomass estimation in Eucalyptus nitens plantations: Effect of bole sampling intensity and explanatory variables. For. Ecol. Manag. 2011, 261, 2016–2028. [Google Scholar] [CrossRef]
- Inagaki, Y.; Nakanishi, A.; Tange, T. A simple method for leaf and branch biomass estimation in Japanese cedar plantations. Trees 2020, 34, 349–356. [Google Scholar] [CrossRef]
- Shinozaki, K.; Yoda, K.; Hozumi, K.; Kira, T. A quantitative analysis of plant form-the pipe model theory: I. Basic analyses. Jpn. J. Ecol. 1964, 14, 97–105. [Google Scholar]
- Shinozaki, K.; Yoda, K.; Hozumi, K.; Kira, T. A quantitative analysis of plant form-the pipe model theory: II. Further evidence of the theory and its application in forest ecology. Jpn. J. Ecol. 1964, 14, 133–139. [Google Scholar]
- Hu, M.; Lehtonen, A.; Minunno, F.; Mäkelä, A. Age effect on tree structure and biomass allocation in Scots pine (Pinus sylvestris L.) and Norway spruce (Picea abies [L.] Karst.). Ann. For. Sci. 2020, 77, 1–15. [Google Scholar] [CrossRef]
- Sumida, A.; Nakai, T.; Yamada, M.; Ono, K.; Uemura, S.; Hara, T. Ground-based estimation of leaf area index and vertical distribution of leaf area density in a Betula ermanii forest. Silva Fenn. 2009, 43, 799–816. [Google Scholar] [CrossRef]
- Berninger, F.; Coll, L.; Vanninen, P.; Mäkelä, A.; Palmroth, S.; Nikinmaa, E. Effects of tree size and position on pipe model ratios in Scots pine. Can. J. For. Res. 2005, 35, 1294–1304. [Google Scholar] [CrossRef]
- Shelburne, V.B.; Hedden, R.L.; Allen, R.M. The effect of site, stand density, and sapwood permeability on the relationship between leaf area and sapwood area in loblolly pine (Pinus taeda L.). For. Ecol. Manag. 1993, 58, 193–209. [Google Scholar] [CrossRef]
- McTague, J.P.; Weiskittel, A. Evolution, history, and use of stem taper equations: A review of their development, application, and implementation. Can. J. For. Res. 2021, 51, 210–235. [Google Scholar] [CrossRef]
- Valentine, H.T.; Baldwin, V.C., Jr.; Gregoire, T.G.; Burkhart, H.E. Surrogates for foliar dry matter in loblolly pine. For. Sci. 1994, 40, 576–585. [Google Scholar]
- Pan, L.; Mei, G.; Wang, Y.; Saeed, S.; Chen, L.; Cao, Y.; Sun, Y. Generalized Nonlinear Mixed-Effect Model of Individual TREE Height to Crown Base for Larix olgensis Henry in Northeast China. J. Sustain. For. 2020, 39, 827–840. [Google Scholar] [CrossRef]
- Zeng, W.; Duo, H.; Lei, X.; Chen, X.; Wang, X.; Pu, Y.; Zou, W. Individual tree biomass equations and growth models sensitive to climate variables for Larix spp. in China. Eur. J. For. Res. 2017, 136, 233–249. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, Y.; Zhang, Z.; Xie, L.; Li, F. Comparison of tree biomass modeling approaches for larch (Larix olgensis Henry) trees in Northeast China. Forests 2020, 11, 202. [Google Scholar] [CrossRef]
- Zhou, L.; Zhu, D.; Li, Z.; Lv, X.; Zhai, X. Research and establishment of the aboveground biomass model of Robinia pseudoacacia L. Acta Agric. Shanghai 2016, 32, 138–142. [Google Scholar] [CrossRef]
- Li, F. Modeling crown profile of Larix olgensis trees. Sci. Silvae Sin. 2004, 40, 16–24. [Google Scholar] [CrossRef]
- Zeng, W.; Tang, S. Goodness evaluation and precision analysis of tree biomass equations. Sci. Silvae Sin. 2011, 47, 106–113. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, L.; Li, F. Evaluation of stand biomass estimation methods for major forest types in the eastern Da Xing’an Mountains, Northeast China. Forests 2019, 10, 715. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, L.; Li, F. Developing additive systems of biomass equations for nine hardwood species in Northeast China. Trees 2015, 29, 1149–1163. [Google Scholar] [CrossRef]
- Zhao, D.; Kane, M.; Markewitz, D.; Teskey, R.; Clutter, M. Additive tree biomass equations for midrotation loblolly pine plantations. For. Sci. 2015, 61, 613–623. [Google Scholar] [CrossRef]
- Baskerville, G.L. Use of logarithmic regression in the estimation of plant biomass. Can. J. For. Res. 1972, 2, 49–53. [Google Scholar] [CrossRef]
- Schlund, M.; Boehm, H.D. Assessment of linear relationships between TanDEM-X coherence and canopy height as well as aboveground biomass in tropical forests. Int. J. Remote Sens. 2021, 42, 3405–3425. [Google Scholar] [CrossRef]
- António, N.; Tomé, M.; Tomé, J.; Soares, P.; Fontes, L. Effect of tree, stand, and site variables on the allometry of Eucalyptus globulus tree biomass. Can. J. For. Res. 2007, 37, 895–906. [Google Scholar] [CrossRef]
- Forrester, D.I.; Tachauer, I.H.H.; Annighoefer, P.; Barbeito, I.; Pretzsch, H.; Ruiz-Peinado, R.; Stark, H.; Vacchiano, G.; Zlatanov, T.; Chakraborty, T.; et al. Generalized biomass and leaf area allometric equations for European tree species incorporating stand structure, tree age and climate. For. Ecol. Manag. 2017, 396, 160–175. [Google Scholar] [CrossRef]
- Sumida, A.; Miyaura, T.; Torii, H. Relationships of tree height and diameter at breast height revisited: Analyses of stem growth using 20-year data of an even-aged Chamaecyparis obtusa stand. Tree Physiol. 2013, 33, 106–118. [Google Scholar] [CrossRef]
- Zapatacuartas, M.; Sierra, C.A.; Alleman, L. Probability distribution of allometric coefficients and Bayesian estimation of aboveground tree biomass. For. Ecol. Manag. 2012, 277, 173–179. [Google Scholar] [CrossRef]
- Rubilar, R.A.; Allen, H.L.; Alvarez, J.S. Silvicultural manipulation and site effect on above and belowground biomass equations for young Pinus radiata. Biomass Bioenerg. 2010, 34, 1825–1837. [Google Scholar] [CrossRef]
- Lehtonen, A.; Heikkinen, J.; Petersson, H.; Ťupek, B.; Liski, E.; Mäkelä, A. Scots pine and Norway spruce foliage biomass in Finland and Sweden—Testing traditional models vs. the pipe model theory. Can. J. For. Res. 2020, 50, 146–154. [Google Scholar] [CrossRef]
- Kantola, A.; Mäkelä, A. Development of biomass proportions in Norway spruce (Picea abies [L.] Karst.). Trees 2006, 20, 111–121. [Google Scholar] [CrossRef]
- Ilomäki, S.; Nikinmaa, E.; Mäkelä, A. Crown rise due to competition drives biomass allocation in silver birch. Can. J. For. Res. 2003, 33, 2395–2404. [Google Scholar] [CrossRef]
- Wassihun, A.N.; Hussin, Y.A.; Van Leeuwen, L.M. Effect of forest stand density on the estimation of above ground biomass/carbon stock using airborne and terrestrial LIDAR derived tree parameters in tropical rain forest, Malaysia. Environ. Syst. Res. 2019, 8, 1–15. [Google Scholar] [CrossRef]
- Dahlhausen, J.; Uhl, E.; Heym, M. Stand density sensitive biomass functions for young oak trees at four different European sites. Trees 2017, 31, 1811–1826. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Dyderski, M.K.; Gęsikiewicz, K. How do tree stand parameters affect young Scots pine biomass?—Allometric equations and biomass conversion and expansion factors. For. Ecol. Manag. 2018, 409, 74–83. [Google Scholar] [CrossRef]
- Thompson Dan, C. The effect of stand structure and stand density on the leaf area–sapwood area relationship of lodgepole pine. Can. J. For. Res. 1989, 19, 392–396. [Google Scholar] [CrossRef]
- Robichaud, E.; Methven, I.R. The applicability of the pipe model theory for the prediction of foliage biomass in trees from natural, untreated black spruce stands. Can. J. For. Res. 1992, 22, 1118–1123. [Google Scholar] [CrossRef]
- Köstner, B.; Falge, E.; Tenhunen, J.D. Age-related effects on leaf area/sapwood area relationships, canopy transpiration and carbon gain of Norway spruce stands (Picea abies) in the Fichtelgebirge, Germany. Tree Physiol. 2002, 22, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, K. Mathematical analysis of age-related changes in leaf biomass in forest stands. Can. J. For. Res. 2012, 42, 356–363. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Plot Number | Area (ha) | Density (n/ha) | Age (Year) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Max | Min | Mean | Max | Min | Mean | Max | Min | Mean | ||
| Hailin | 50 | 0.1225 | 0.0100 | 0.0735 | 7800 | 475 | 1653.36 | 40 | 6 | 27.37 |
| Dahailin | 48 | 0.1225 | 0.0400 | 0.0635 | 4525 | 542 | 1822.14 | 40 | 12 | 20.80 |
| Linkou | 51 | 0.1200 | 0.0225 | 0.0805 | 3967 | 500 | 1567.28 | 40 | 6 | 20.20 |
| Huanan | 50 | 0.1200 | 0.0100 | 0.0738 | 7800 | 408 | 1843.89 | 40 | 9 | 26.59 |
| Total | 199 | 0.1225 | 0.0100 | 0.0730 | 7800 | 408 | 1719.88 | 40 | 6 | 23.76 |
| Site | Plot Number | Sample Trees Number | DBH (cm) | Height (m) | HB (m) | CW (m) | CL (m) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Max | Min | Mean ± Std | CV | Max | Min | Mean ± Std | CV | Max | Min | Mean ± Std | CV | Max | Min | Mean ± Std | CV | Max | Min | Mean ± Std | CV | |||
| Hailin | 50 | 100 | 27.8 | 5.8 | 14.7 ± 5.46 | 0.37 | 25.95 | 3.30 | 14.62 ± 5.45 | 0.37 | 15.90 | 0.21 | 6.63 ± 4.75 | 0.72 | 6.35 | 1.25 | 3.42 ± 1.01 | 0.29 | 11.90 | 2.35 | 7.99 ± 1.97 | 0.25 |
| Dahailin | 48 | 96 | 26.7 | 8.1 | 14.0 ± 4.46 | 0.32 | 25.33 | 9.68 | 14.41 ± 3.90 | 0.27 | 17.80 | 1.10 | 6.58 ± 3.51 | 0.53 | 7.55 | 0.83 | 3.19 ± 1.18 | 0.37 | 12.85 | 2.92 | 7.82 ± 1.90 | 0.24 |
| Linkou | 51 | 102 | 25.8 | 5.0 | 16.3 ± 4.40 | 0.27 | 24.45 | 2.70 | 15.65 ± 3.75 | 0.24 | 16.70 | 0.98 | 7.47 ± 3.23 | 0.43 | 6.00 | 1.00 | 2.84 ± 1.14 | 0.40 | 14.30 | 1.65 | 8.18 ± 2.12 | 0.26 |
| Huanan | 50 | 100 | 29.6 | 5.0 | 16.8 ± 6.07 | 0.36 | 24.40 | 3.80 | 16.55 ± 5.49 | 0.33 | 16.50 | 0.50 | 7.86 ± 4.63 | 0.59 | 5.90 | 1.25 | 3.32 ± 1.10 | 0.33 | 14.70 | 2.10 | 8.69 ± 2.49 | 0.29 |
| Total | 199 | 398 | 29.6 | 5.0 | 15.5 ± 5.27 | 0.34 | 25.95 | 2.70 | 15.32 ± 4.80 | 0.31 | 17.80 | 0.21 | 7.14 ± 4.12 | 0.58 | 7.55 | 0.83 | 3.19 ± 1.13 | 0.35 | 14.70 | 1.65 | 8.17 ± 2.16 | 0.26 |
| Site | Plot Number | Sample Trees Number | Leaf Biomass (kg) | Branch Biomass (kg) | Crown Biomass (kg) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Max | Min | Mean ± Std | CV | Max | Min | Mean ± Std | CV | Max | Min | Mean ± Std | CV | |||
| Hailin | 50 | 100 | 5.87 | 0.44 | 2.16 ± 1.26 | 0.58 | 46.72 | 2.91 | 14.49 ± 9.67 | 0.67 | 52.59 | 3.46 | 16.65 ± 10.92 | 0.66 |
| Dahailin | 48 | 96 | 6.14 | 0.37 | 1.92 ± 1.12 | 0.59 | 49.11 | 1.91 | 12.45 ± 8.91 | 0.72 | 55.25 | 2.28 | 14.36 ± 10.02 | 0.70 |
| Linkou | 51 | 102 | 7.21 | 0.26 | 2.49 ± 1.28 | 0.51 | 54.08 | 1.51 | 16.18 ± 9.56 | 0.59 | 61.28 | 1.78 | 18.67 ± 10.81 | 0.58 |
| Huanan | 50 | 100 | 7.20 | 0.26 | 2.69 ± 1.63 | 0.60 | 54.71 | 1.27 | 18.55 ± 12.73 | 0.69 | 61.90 | 1.53 | 21.24 ± 14.35 | 0.68 |
| Total | 199 | 398 | 7.21 | 0.26 | 2.32 ± 1.37 | 0.59 | 54.71 | 1.27 | 15.45 ± 10.57 | 0.68 | 61.90 | 1.53 | 17.77 ± 11.93 | 0.67 |
| X | Slope(a) | Intercept(b) | R2 | R2adj | ||
|---|---|---|---|---|---|---|
| a | 95% CI | b | 95% CI | |||
| Leaf biomass | ||||||
| AB | 0.974 | (0.948, 1.001) | 5.156 | (5.032, 5.280) | 0.944 | 0.944 |
| D2H | 0.533 | (0.505, 0.560) | 1.298 | (1.252, 1.343) | 0.825 | 0.825 |
| Branch biomass | ||||||
| AB | 1.102 | (1.075, 1.129) | 7.591 | (7.462, 7.720) | 0.952 | 0.952 |
| D2H | 0.587 | (0.553, 0.620) | 3.208 | (3.151, 3.264) | 0.790 | 0.789 |
| Crown biomass | ||||||
| AB | 1.084 | (1.057, 1.110) | 7.654 | (7.529, 7.779) | 0.953 | 0.953 |
| D2H | 0.579 | (0.547, 0.612) | 3.347 | (3.292, 3.401) | 0.797 | 0.796 |
| Effect Factors | Leaf Biomass/AB (kg m−2) | Branch Biomass/AB (kg m−2) | Crown Biomass/AB (kg m−2) |
|---|---|---|---|
| Site | |||
| Hailin | 196.08 ± 28.58 a | 1284.35 ± 259.29 a | 1480.43 ± 281.28 a |
| Dahailin | 198.72 ± 27.98 a | 1284.35 ± 259.29 a | 1415.60 ± 213.92 a |
| Linkou | 199.58 ± 43.64 a | 1220.54 ± 169.18 a | 1420.12 ± 192.07 a |
| Huanan | 196.56 ± 28.60 a | 1278.90 ± 239.23 a | 1475.46 ± 261.45 a |
| Stand density (stems ha−1) | |||
| <1000 | 197.31 ± 44.65 a | 1246.10 ± 264.89 a | 1443.41 ± 288.95 a |
| 1000–2000 | 197.99 ± 27.76 a | 1262.81 ± 214.84 a | 1460.80 ± 235.37 a |
| >2000 | 197.61 ± 25.97 a | 1222.75 ± 174.90 a | 1420.36 ± 189.26 a |
| Age | |||
| <20 | 208.09 ± 37.66 a | 1231.08 ± 152.27 a | 1439.17 ± 170.51 a |
| 20–30 | 193.69 ± 23.63 b | 1260.02 ± 208.20 a | 1453.71 ± 225.90 a |
| >30 | 189.21 ± 31.57 b | 1260.30 ± 300.96 a | 1449.51 ± 327.88 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, C.; Zhang, Y.; Chen, L.; Zhuang, L.; Zhang, Y.; Sang, W. Pipe Model Can Accurately Estimate Crown Biomass of Larch (Larix olgensis) Plantation Forest in Northeast China. Forests 2023, 14, 400. https://doi.org/10.3390/f14020400
Huang C, Zhang Y, Chen L, Zhuang L, Zhang Y, Sang W. Pipe Model Can Accurately Estimate Crown Biomass of Larch (Larix olgensis) Plantation Forest in Northeast China. Forests. 2023; 14(2):400. https://doi.org/10.3390/f14020400
Chicago/Turabian StyleHuang, Chenyu, Yuanyuan Zhang, Lu Chen, Liwen Zhuang, Yanliang Zhang, and Weiguo Sang. 2023. "Pipe Model Can Accurately Estimate Crown Biomass of Larch (Larix olgensis) Plantation Forest in Northeast China" Forests 14, no. 2: 400. https://doi.org/10.3390/f14020400
APA StyleHuang, C., Zhang, Y., Chen, L., Zhuang, L., Zhang, Y., & Sang, W. (2023). Pipe Model Can Accurately Estimate Crown Biomass of Larch (Larix olgensis) Plantation Forest in Northeast China. Forests, 14(2), 400. https://doi.org/10.3390/f14020400