
Stoichiometric Soil Microbial and Enzymatic Characteristics under Three Different Plantation Types in China’s Luya Mountain

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Soil Sample Collection and Measurements
2.3. Soil Enzyme Activities and Microbial Nutrient Elements
2.4. Microbial Sequencing Technology
2.5. Data Analyses
2.6. Statistical Analysis
3. Results
3.1. Soil Physiochemical Properties
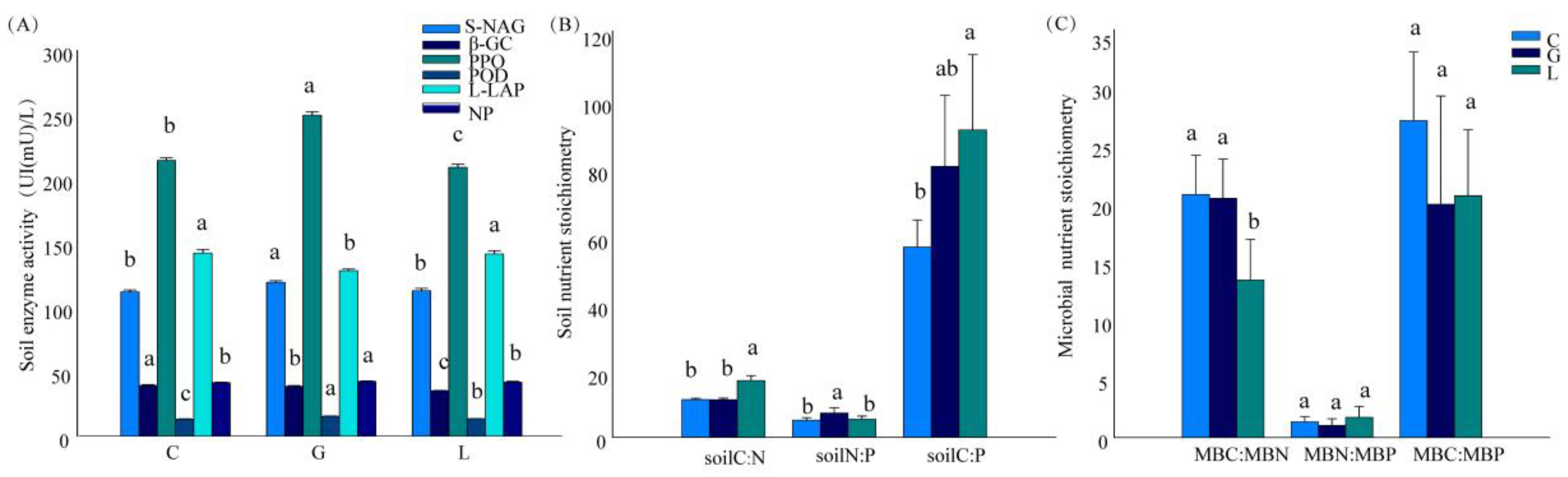
3.2. Variation in Soil Enzyme Activities
3.3. Stoichiometric Characteristics of Soil and Microbial Nutrients
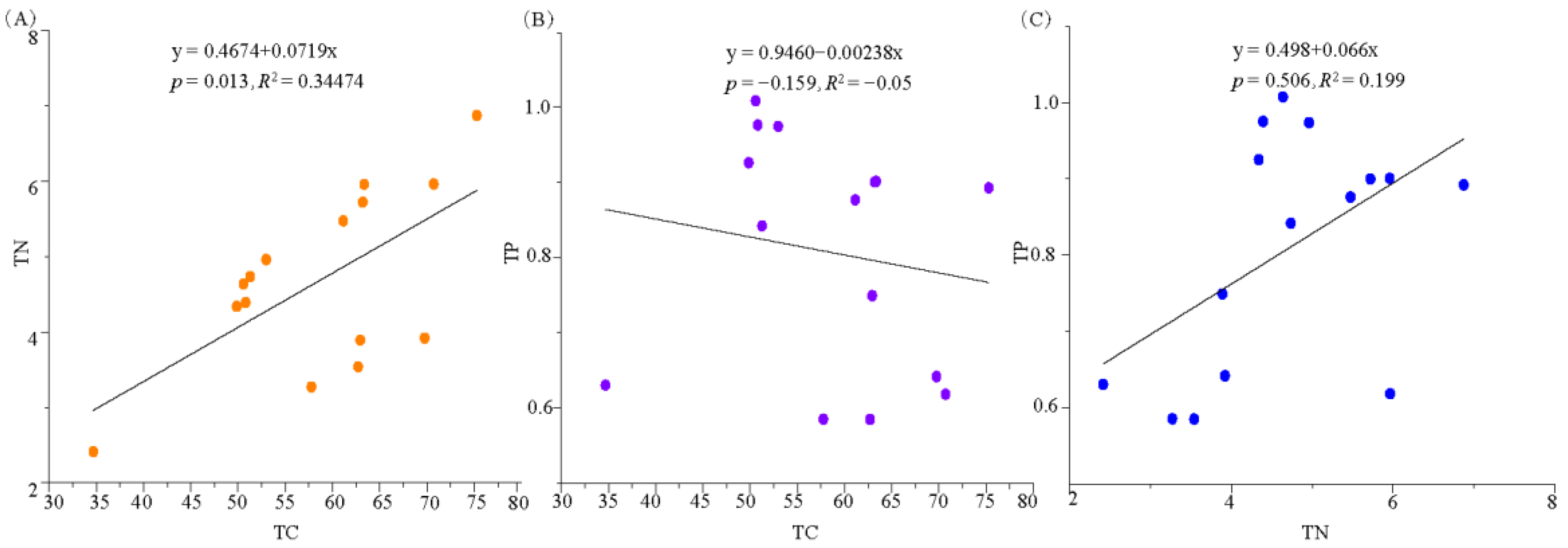
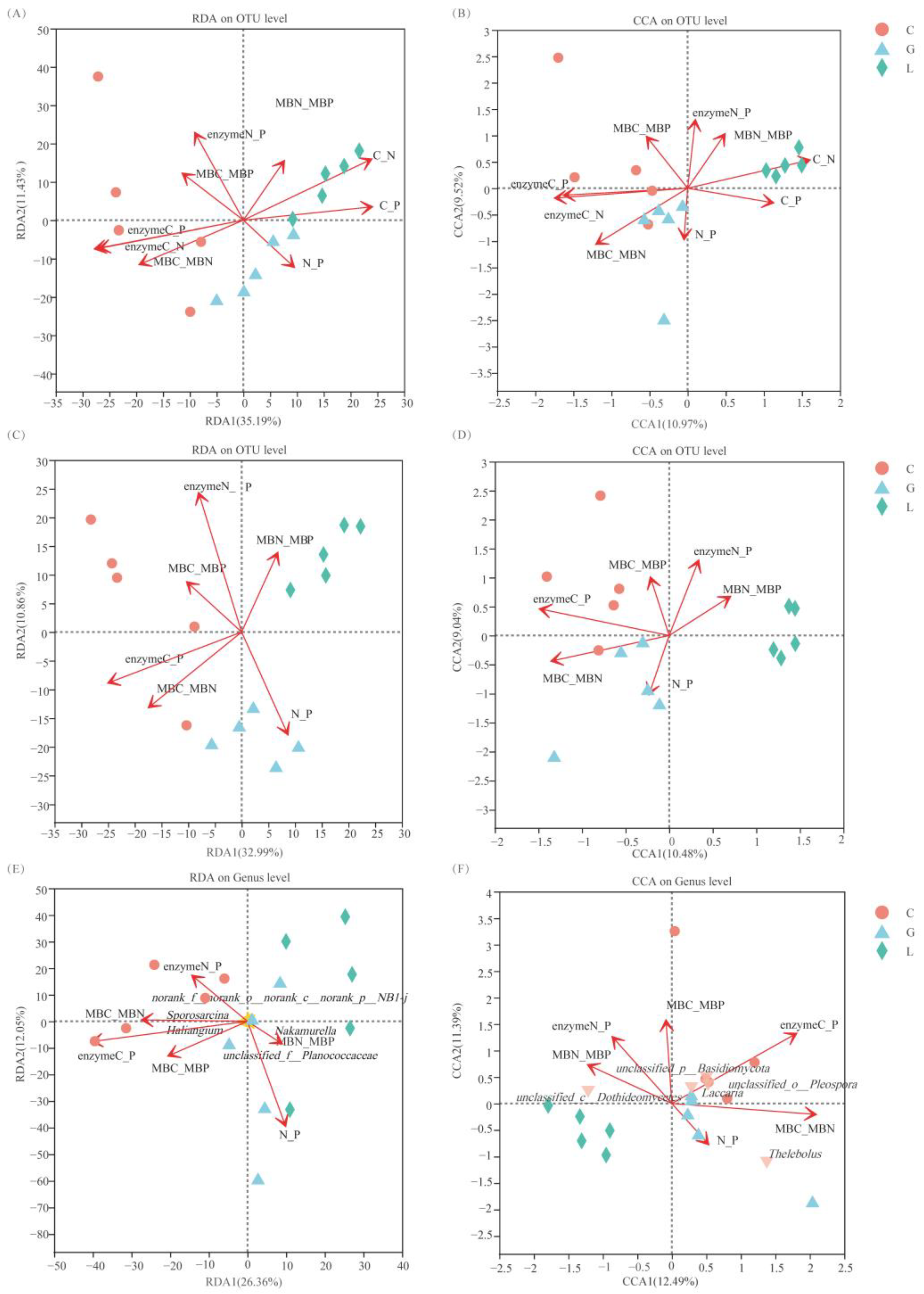
3.4. Correlation Analysis of Soil Extracellular Enzymes and Microbial Biomass Stoichiometry with Environmental Factors
3.5. Ecoenzymatic Stoichiometry
3.6. Characteristics of Soil Bacterial and Fungal Communities under the Different Plantation Types
3.6.1. Soil Bacterial and Fungal Community Diversity of the Different Plantation Types
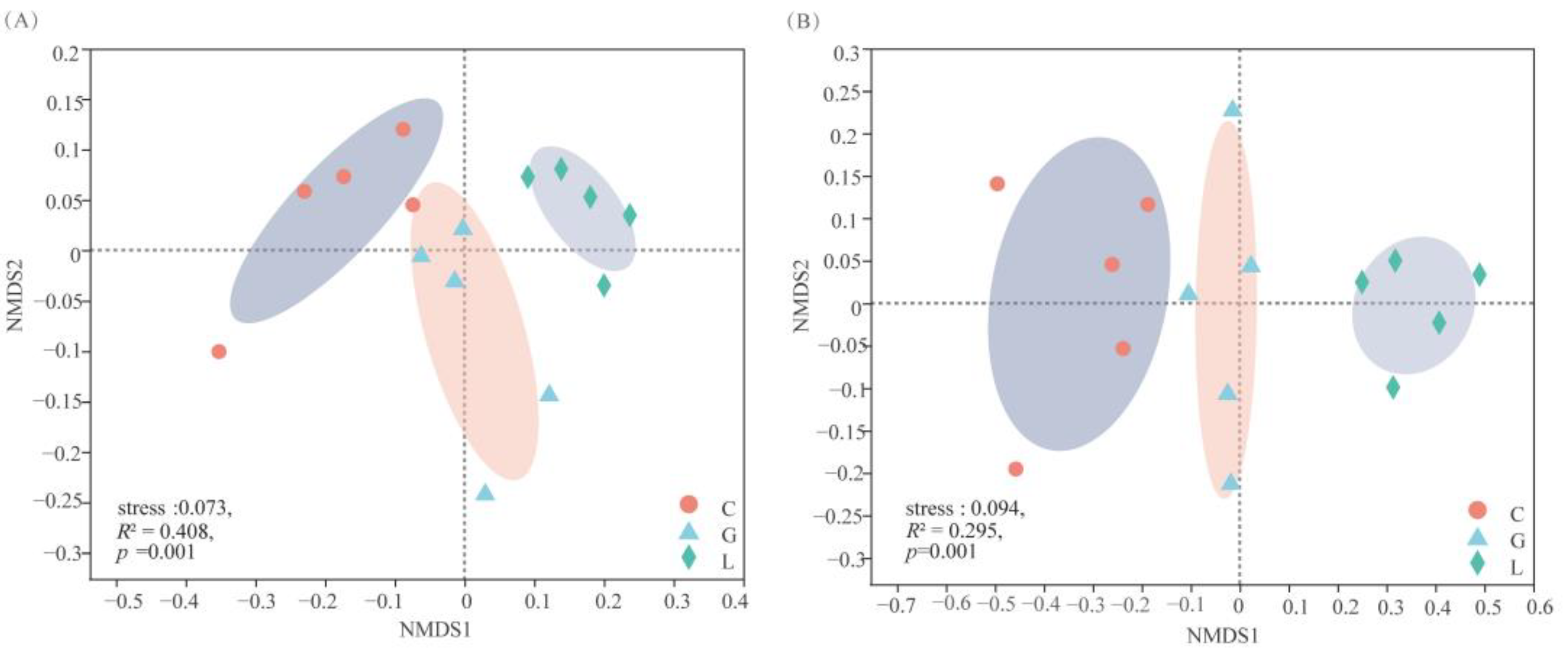
3.6.2. Soil Bacterial and Fungal Community Structure under the Different Plantation Types
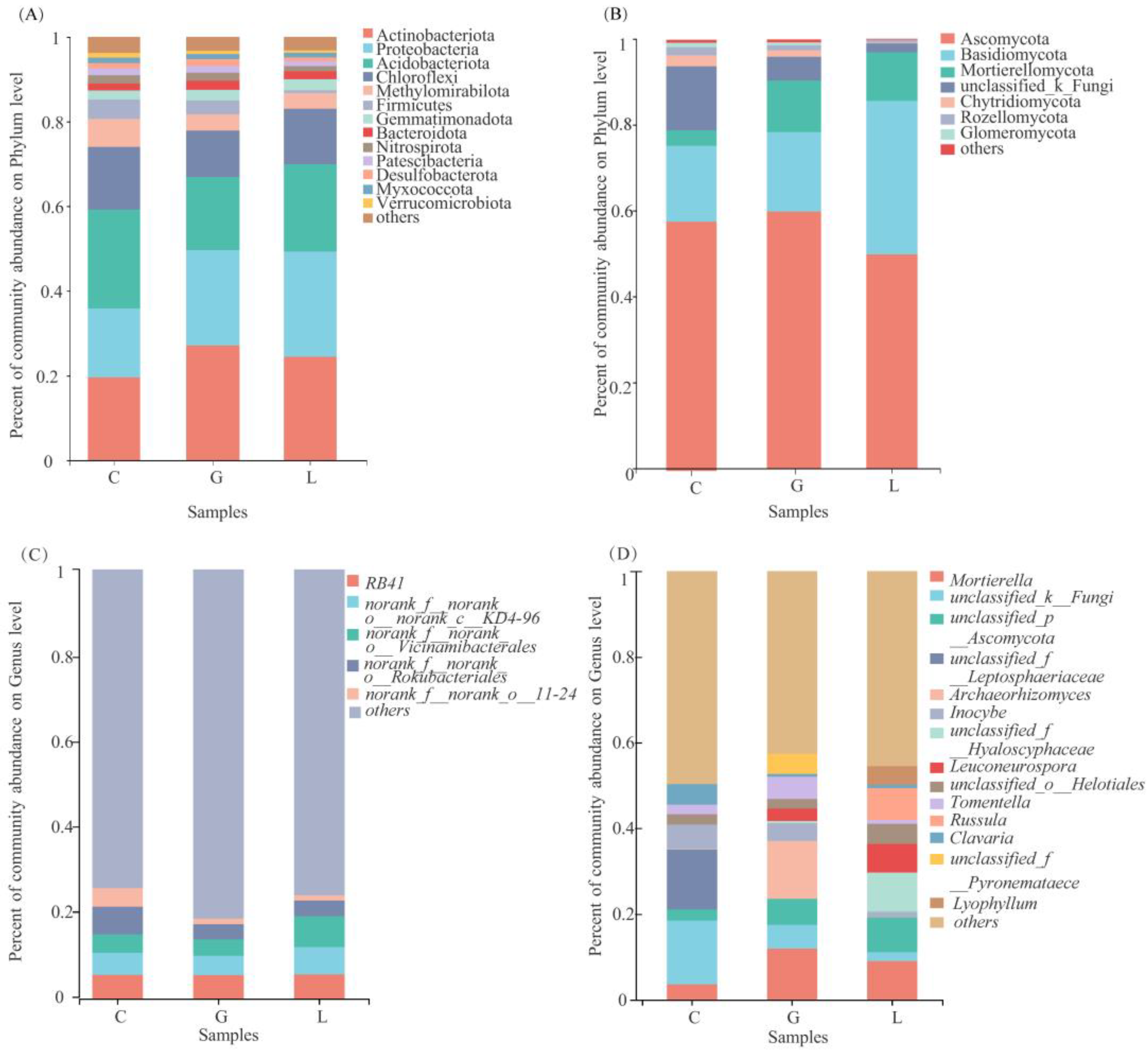
3.6.3. Soil Microbial Community Composition
3.6.4. Analysis of Bacterial and Fungal Community Composition in the Different Plantation Types
3.7. Correlation between Soil Enzyme Stoichiometry and Dominant Species
3.8. Stoichiometry Relative to Environmental Factors and Dominant Microbial Groups
4. Discussion
4.1. Effects of the Different Plantation Types on Soil Nutrients, Enzyme Activities, and Microbial Nutrients
4.2. Relationships among Soil Extracellular Enzymes, Microbial Nutrients, and Soil Nutrient Stoichiometry under the Different Plantation Types
4.3. Key Soil Microbial and Soil Extracellular Enzyme Stoichiometry Factors under Different Plantation Types
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yifan, R.; Mingxiang, X.; Binbin, L.; Luyang, M.; Shengmin, Z. Response of ecological stoichiometry characteristics of different soil-microorganism-plant systems to different fertility levels in the Loess hilly region. J. Northwest AF Univ. (Nat. Sci. Ed.) 2017, 45, 77–84. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chao, L.; Yunqiang, W.; Huan, C.; Shaoshan, A.; Scott, X.C. Soil extracellular enzyme stoichiometry reflects the shift from P- to N-limitation of microorganisms with grassland restoration. Soil Biol. Biochem. 2020, 149. [Google Scholar] [CrossRef]
- Zheng, H.; Vesterdal, L.; Schmidt, I.K.; Rousk, J. Ecoenzymatic stoichiometry can reflect microbial resource limitation, substrate quality, or both in forest soils. Soil Biol. Biochem. 2022, 167. [Google Scholar] [CrossRef]
- Lixin, Z.; Yuxi, D.; Bo, W.; Weifeng, W.; Xiaojing, L.; Jinjie, L. Characteristics of soil microorganisms and soil nutrients in different sand-fixation shrub plantations in Kubuqi Desert, China. Chin. J. Appl. Ecol. 2017, 28, 3871–3880. [Google Scholar] [CrossRef]
- Miaoping, X.; Chengjie, R.; Wei, Z.; Zhengxing, C.; Shuyue, F.; Weichao, L.; Gaihe, Y.; Xinhui, Y. Responses mechanism of C:N:P stoichiometry of soil microbial biomass and soil enzymes to climate change. Chin. J. Appl. Ecol. 2018, 29, 2445–2454. [Google Scholar] [CrossRef]
- Guisheng, Y.; Changchun, S.; Zhongmei, W.; Yanyu, S. Microbial activity in soils of Calamagrostis angustifolia wetlands in the Sanjiang Plain. Acta Sci. Circumstantiae 2010, 30, 1715–1721. [Google Scholar] [CrossRef]
- Qianyuan, L.; Xingliang, X.; Huimin, W.; Evgenia, B.; Yakov, K. Dominant extracellular enzymes in priming of SOM decomposition depend on temperature. Geoderma 2019, 343, 187–195. [Google Scholar] [CrossRef]
- Waring, B.G.; Weintraub, S.R.; Sinsabaugh, R.L. Ecoenzymatic stoichiometry of microbial nutrient acquisition in tropical soils. (Special Issue: Enzymes in biogeochemical cycles: Integrating experimental data, theory, and models). Biogeochemistry 2014, 117, 101–113. [Google Scholar] [CrossRef]
- Jingjing, W.; Gang, H.; Kun, L.; Yangui, S. Soil enzyme activity and stoichiometric characteristics of different vegetation types in tropical and subtropical forests. Chin. J. Appl. Environ. Biol. 2022, 1–12. [Google Scholar] [CrossRef]
- Xiaomeng, W.; Bahar, S.R.; Yajun, H.; Xingliang, X.; Zhenke, Z.; Yuhuai, L.; Yakov, K.; Yong, L.; Jinshui, W.; Tida, G. C/P stoichiometry of dying rice root defines the spatial distribution and dynamics of enzyme activities in root-detritusphere. Biol. Fertil. Soils 2019, 55, 251–263. [Google Scholar] [CrossRef]
- Liang, W.; Bahar, S.R.; Weiqi, W.; Zhenke, Z.; Shoulong, L.; Jinshui, W.; Yakov, K.; Tida, G. Labile carbon matters more than temperature for enzyme activity in paddy soil. Soil Biol. Biochem. 2019, 135, 134–143. [Google Scholar] [CrossRef]
- Wei, L.; Ge, T.; Zhu, Z.; Ye, R.; Peñuelas, J.; Li, Y.; Lynn, T.M.; Jones, D.L.; Jinshui, W.; Kuzyakov, Y. Paddy soils have a much higher microbial biomass content than upland soils: A review of the origin, mechanisms, and drivers. Agric. Ecosyst. Environ. 2022, 326, 107798. [Google Scholar] [CrossRef]
- Pingli, C.; Siyi, S.; Shengxu, L.; Jinxin, Z.; Xiaofei, L.; Jianfen, G. Response of Soil Extracellular Enzymes Activities to Throughfall Exclusion at Different Depths in a Subtropical Natural Castanopsis carlesii Forest. J. Soil Water Conserv. 2018, 36, 205–211. [Google Scholar] [CrossRef]
- Lei, H.; Zhishan, Z. Effect of rainfall pulses on plant growth and transpiration of two xerophytic shrubs in a revegetated desert area: Tengger Desert, China. Catena 2016, 137, 269–276. [Google Scholar] [CrossRef]
- Bell, C.W.; Acosta-Martinez, V.; McIntyre, N.E.; Cox, S.; Tissue, D.T.; Zak, J.C. Linking microbial community structure and function to seasonal differences in soil moisture and temperature in a Chihuahuan desert grassland. Microb. Ecol. 2009, 58, 827–842. [Google Scholar] [CrossRef]
- Zhenghu, Z.; Chuankuan, W.; Yiqi, L. Response of soil microbial communities to altered precipitation: A global synthesis. Glob. Ecol. Biogeogr. 2022, 27, 1121–1136. [Google Scholar] [CrossRef]
- He, Q.; Wu, Y.; Bing, H.; Zhou, J.; Wang, J. Vegetation type rather than climate modulates the variation in soil enzyme activities and stoichiometry in subalpine forests in the eastern Tibetan Plateau. Geoderma 2020, 374. [Google Scholar] [CrossRef]
- Xiangqian, X.; Haikuo, Z.; Yasi, F.; Jipeng, W.; Chenfei, L.; Youchao, C.; Gaodi, Z.; Yanjiang, C. Effects of plant residues on the stoichiometric characteristics of soil, microorganisms and extracellular enzymes C:N:P in an alpine meadow on the Tibetan Plateau. Chin. J. Appl. Ecol. 2022, 34, 58–66. [Google Scholar] [CrossRef]
- Chen, X.; Feng, J.; Ding, Z.; Tang, M.; Zhu, B. Changes in soil total, microbial and enzymatic C-N-P contents and stoichiometry with depth and latitude in forest ecosystems. Sci. Total Environ. 2021, 816. [Google Scholar] [CrossRef]
- Jianrong, G. Preliminary Report on Forest Ecosystem Services Evaluation in Shanxi Luya Mountain National Nature Reserve. For. Sci. Technol. 2021, 9, 68–71. [Google Scholar] [CrossRef]
- Xiaogang, W.; Jinping, G.; Xiuyun, Y.; Xuping, T. Soil organic carbon storage and profile inventory in the different vegetation types of Luya Mountai. Acta Ecol. Sin. 2011, 31, 3009–3019. [Google Scholar] [CrossRef]
- Jia, T.; Guo, T.Y.; Yao, Y.S.; Wang, R.H.; Chai, B.F. Seasonal Microbial Community Characteristic and Its Driving Factors in a Copper Tailings Dam in the Chinese Loess Plateau. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Spohn, M.; Klaus, K.; Wanek, W.; Richter, A. Microbial carbon use efficiency and biomass turnover times depending on soil depth—Implications for carbon cycling. Soil Biol. Biochem. 2016, 96, 74–81. [Google Scholar] [CrossRef]
- Carreiro, M.M.; Sinsabaugh, R.L.; Repert, D.A.; Parkhurst, D.F. Microbial enzyme shifts explain litter decay responses to simulated nitrogen deposition. Ecology 2000, 81, 2359–2365. [Google Scholar] [CrossRef]
- Jia, T.; Wang, Y.W.; Chai, B. Bacterial Community Characteristics and Enzyme Activities in Bothriochloa ischaemum Litter over Progressive Phytoremediation Years in a Copper Tailings Dam. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Jia, T.; Wang, R.; Chai, B. Effects of heavy metal pollution on soil physicochemical properties and microbial diversity over different reclamation years in a copper tailings dam. J. Soil Water Conserv. 2019, 74, 439–448. [Google Scholar] [CrossRef]
- Daryl, L.M.; Robert, L.S.; Brian, H.H.; Michael, N.W. Vector analysis of ecoenzyme activities reveal constraints on coupled C, N and P dynamics. Soil Biol. Biochem. 2016, 93. [Google Scholar] [CrossRef]
- Miaomiao, W.; Hongsong, C.; Tonggang, F.; Wei, Z.; Kelin, W. Differences of soil nutrients among different vegetation types and their spatial prediction in a small typical karst catchment. Chin. J. Appl. Ecol. 2016, 27, 1759–1766. [Google Scholar] [CrossRef]
- Mei, W.; Ziran, Y.; Ziwen, Z.; Yang, W.; Wenjing, C.; Yixiao, Y.; Guobin, L.; Sha, X. Variation Characteristics of Specific Soil Enzyme Activities During Vegetation Succession on the Loess Plateau. J. Soil Water Conserv. 2021, 35, 181–187. [Google Scholar] [CrossRef]
- Wang, H.Y.; Wu, J.Q.; Li, G.; Yan, L.J. Changes in soil carbon fractions and enzyme activities under different vegetation types of the northern Loess Plateau. Ecol. Evol. 2020, 10, 12211–12223. [Google Scholar] [CrossRef]
- Zhiwei, X.; Guirui, Y.; Xinyu, Z.; Nianpeng, H.; Qiufeng, W.; Shengzhong, W.; Ruili, W.; Ning, Z.; Yaolong, J.; Chunyan, W. Soil enzyme activity and stoichiometry in forest ecosystems along the North-South Transect in eastern China (NSTEC). Soil Biol. Biochem. 2017, 104, 152–163. [Google Scholar] [CrossRef]
- Xiaoqian, P.; Wei, W. Stoichiometry of soil extracellular enzyme activity along a climatic transect in temperate grasslands of northern China. Soil Biol. Biochem. 2016, 98, 74–84. [Google Scholar] [CrossRef]
- Jack, T.T.; Dudley, J.R. Use of Nitrogen to Phosphorus Ratios in Plant Tissue as an Indicator of Nutrient Limitation and Nitrogen Saturation. J. Appl. Ecol. 2003, 40, 523–534. [Google Scholar] [CrossRef]
- Wang, J.; Wu, Y.; Li, J.; He, Q.; Bing, H. Soil enzyme stoichiometry is tightly linked to microbial community composition in successional ecosystems after glacier retreat. Soil Biol. Biochem. 2021, 162. [Google Scholar] [CrossRef]
- Ge, C.Z.; Wang, R.Q.; Chai, Y.C.; Wang, H.Q.; Kan, M.M.; Liu, J. High Colonization Possibility of Some Species of Weeds in Suaeda salsa Community: From an Ecological Stoichiometry Perspective. PLoS ONE 2017, 12, e0170401. [Google Scholar] [CrossRef]
- Christian, L.L.; Michael, S.S.; Mark, A.B.; Noah, F. The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol. Biochem. 2008, 40, 2407–2415. [Google Scholar] [CrossRef]
- Jincheng, W.; Haiyan, L.; Jian, L. Composition Study of Soil Microorganism in Natural Chinese Pine Forest with Different Growth Years in Ziwuling Forest Region of the Loess Plateau. Bull. Soil Water Conserv. 2015, 35, 2481–2493. [Google Scholar] [CrossRef]
- Narendrula-Kotha, R.; Nkongolo, K.K. Microbial Response to Soil Liming of Damaged Ecosystems Revealed by Pyrosequencing and Phospholipid Fatty Acid Analyses. PLoS ONE 2017, 12, e0168497. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Zhigang, H.; Hanxi, X.; Youfeng, L.; Wanxia, P. Changes of soil microbial biomass carbon,nitrogen,and enzyme activities in East Dongting Lake wetlands at different water levels. Chin. J. Appl. Ecol. 2021, 32, 2958–2966. [Google Scholar] [CrossRef]
- Huiying, L.; Jiacong, Z.; Quanxin, Z.; Jun, S.; Huan, X.; Yuanyuan, L.; Kongcan, M.; Yue, W.; Xiaochun, Y.; Junmei, W.; et al. Soil enzyme stoichiometry revealed the changes of soil microbial carbon and phosphorus limitation along an elevational gradient in a Pinus taiwanensis forest of Wuyi Mountains,Southeast China. Chin. J. Appl. Ecol. 2022, 33, 33–41. [Google Scholar] [CrossRef]
- Petr, Č.; Robert, S.; Kirsten, S.H.; Ben, B.; Nancy, H. Apparent temperature sensitivity of soil respiration can result from temperature driven changes in microbial biomass. Soil Biol. Biochem. 2019, 135, 286–293. [Google Scholar] [CrossRef]
- Cui, Y.; Bing, H.; Fang, L.; Jiang, M.; Shen, G.; Yu, J.; Wang, X.; Zhu, H.; Wu, Y.; Zhang, X. Extracellular enzyme stoichiometry reveals the carbon and phosphorus limitations of microbial metabolisms in the rhizosphere and bulk soils in alpine ecosystems. Plant Soil 2019, 458, 7–20. [Google Scholar] [CrossRef]
- Donovan, P.G.; Kathleen, R.B.M.; Madeleine, M.S.; Steven, D.A. The Michaelis– Menten kinetics of soil extracellular enzymes in response to temperature: A cross-latitudinal study. Glob. Chang. Biol. 2012, 18, 1468–1479. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Type | Geographic Coordinates | Altitude/m | Aspect | Slope/(°) |
|---|---|---|---|---|
| Picea asperata forest | 111°54′14″ E 38°44′26″ N | 2280 | Semi-negative slope | 9.8° |
| Caragana jubata shrub | 111°50′22″ E 38°43′43″ N | 2750 | South slope | 0° |
| Carex lanceolata meadow | 111°50′22″ E 38°43′43″ N | 2750 | South slope | 0° |
| C | G | L | |
|---|---|---|---|
| SWC (%) | 0.028 ± 0.003 b | 0.039 ± 0.002 ab | 0.057 ± 0.026 a |
| pH | 6.528 ± 0.276 a | 6.715 ± 0.265 a | 6.505 ± 0.177 a |
| TN (g/kg) | 4.809 ± 0.566 b | 5.801 ± 0.782 a | 3.406 ± 0.617 c |
| TC (g/kg) | 53.493 ± 5.56 a | 64.356 ± 9.249 a | 57.573 ± 13.487 a |
| SOC (g/kg) | 51.689 ± 5.46 a | 62.238 ± 9.25 a | 55.524 ± 12.96 a |
| TP (g/kg) | 0.956 ± 0.043 a | 0.826 ± 0.118 b | 0.638 ± 0.067 c |
| SOM (g/kg) | 89.111 ± 9.414 a | 107.299 ± 15.947 a | 95.723 ± 22.343 a |
| NO3−-N (mg/kg) | 1.234 ± 0.426 a | 1.245 ± 0.462 a | 1.136 ± 0.084 a |
| NH4+-N (mg/kg) | 31.645 ± 4.511 a | 24.915 ± 3.326 b | 32.96 ± 4.3 a |
| Vector L | Vector A | |
|---|---|---|
| C | 1.19 ± 0.006 a | 33.93 ± 0.097 b |
| G | 1.18 ± 0.005 b | 34.24 ± 0.030 a |
| L | 1.15 ± 0.005 c | 34.01 ± 0.075 b |
| C | G | L | ||
|---|---|---|---|---|
| Sobs | bacteria | 1808.000 ± 135.220 a | 1903.200 ± 98.210 a | 1801.200 ± 89.256 a |
| fungi | 299.800 ± 89.433 b | 424.200 ± 122.608 ab | 524.600 ± 59.622 a | |
| Shannon | bacteria | 6.000 ± 0.093 b | 6.238 ± 0.104 a | 6.120 ± 0.108 ab |
| fungi | 3.638 ± 1.094 a | 3.704 ± 0.969 a | 3.589 ± 0.390 a | |
| Simpson | bacteria | 0.007 ± 0.001 a | 0.005 ± 0.001 c | 0.006 ± 0.001 b |
| fungi | 0.131 ± 0.196 a | 0.124 ± 0.155 a | 0.072 ± 0.019 a | |
| Ace | bacteria | 2843.900 ± 504.220 a | 2691.909 ± 104.928 a | 2622.585 ± 173.886 a |
| fungi | 311.215 ± 91.264 b | 451.525 ± 144.021 b | 595.417 ± 58.499 a | |
| Chao | bacteria | 2623.274 ± 267.273 a | 2690.841 ± 71.938 a | 2539.597 ± 115.243 a |
| fungi | 313.440 ± 92.766 b | 457.557 ± 149.277 ab | 593.096 ± 57.298 a | |
| Coverage | bacteria | 0.966 ± 0.004 a | 0.966 ± 0.001 a | 0.968 ± 0.001 a |
| fungi | 1.000 ± 0 a | 0.999 ± 0.001 a | 0.998 ± 0.001 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Zheng, M.; Zhang, Y.; Chen, Y.; Zhao, L.; Chai, B.; Jia, T. Stoichiometric Soil Microbial and Enzymatic Characteristics under Three Different Plantation Types in China’s Luya Mountain. Forests 2023, 14, 558. https://doi.org/10.3390/f14030558
Wang X, Zheng M, Zhang Y, Chen Y, Zhao L, Chai B, Jia T. Stoichiometric Soil Microbial and Enzymatic Characteristics under Three Different Plantation Types in China’s Luya Mountain. Forests. 2023; 14(3):558. https://doi.org/10.3390/f14030558
Chicago/Turabian StyleWang, Xuerong, Mengyao Zheng, Yue Zhang, Ying Chen, Lijuan Zhao, Baofeng Chai, and Tong Jia. 2023. "Stoichiometric Soil Microbial and Enzymatic Characteristics under Three Different Plantation Types in China’s Luya Mountain" Forests 14, no. 3: 558. https://doi.org/10.3390/f14030558
APA StyleWang, X., Zheng, M., Zhang, Y., Chen, Y., Zhao, L., Chai, B., & Jia, T. (2023). Stoichiometric Soil Microbial and Enzymatic Characteristics under Three Different Plantation Types in China’s Luya Mountain. Forests, 14(3), 558. https://doi.org/10.3390/f14030558