
Effect of Different Vegetation Restoration on Recovery of Compaction-Induced Soil Degradation in Hyrcanian Mixed Forests: Influence on Soil C and N Pools and Enzyme Activities


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
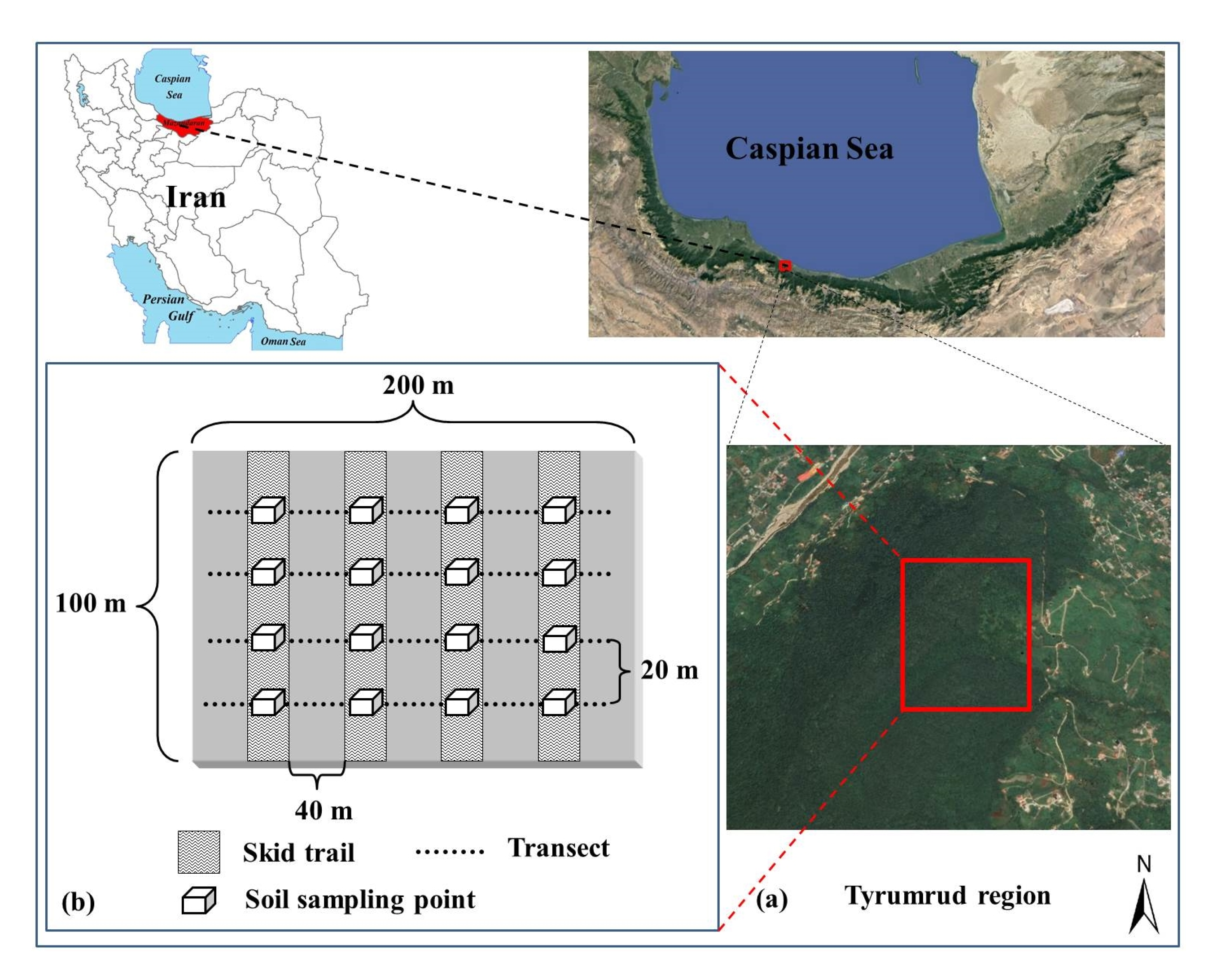
2.1. Site Description
2.2. Experimental Design
2.3. Data Collection and Laboratory Analysis
2.3.1. Litter Properties
2.3.2. Soil Physical Properties
2.3.3. Soil Chemical Properties
2.3.4. Soil Biochemical, Biological and Microbial Properties, and Enzyme Activity
2.4. Statistical Analyses
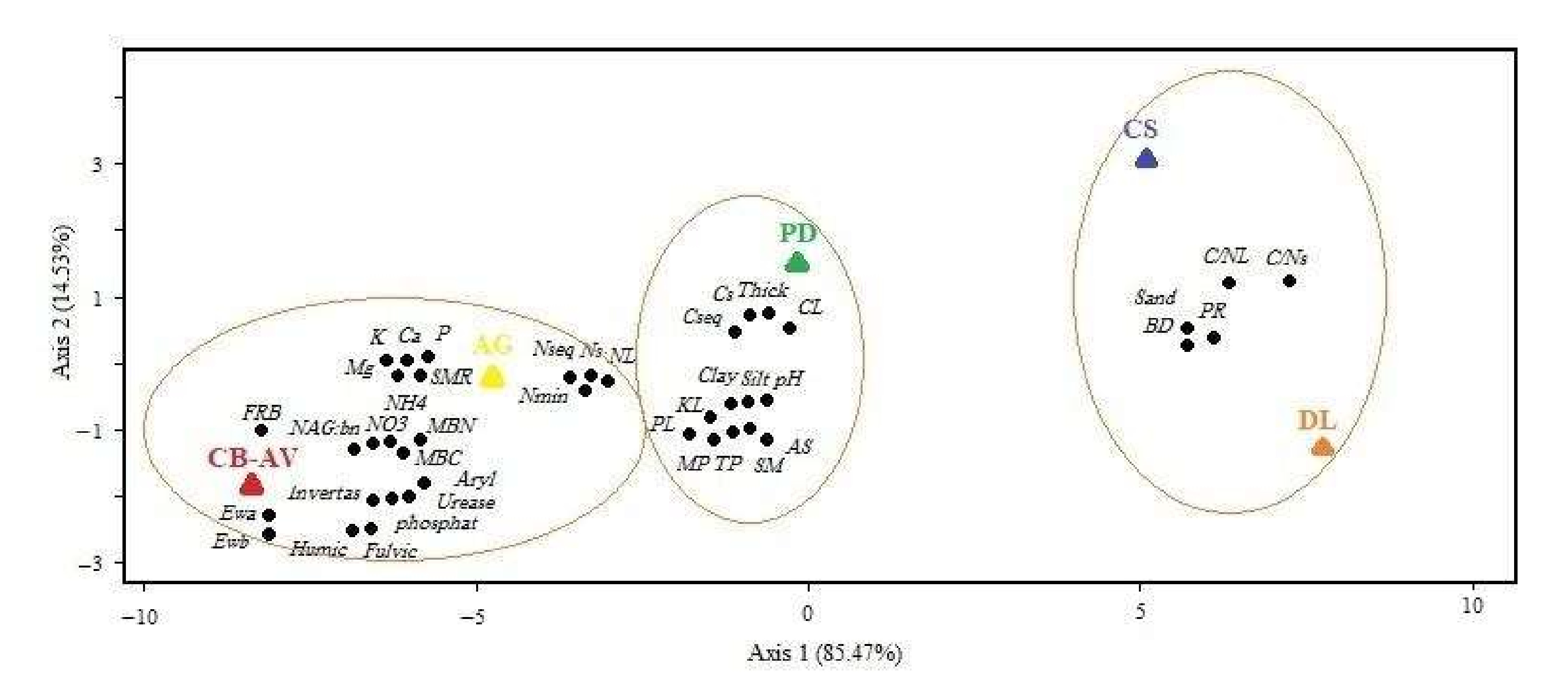
3. Results
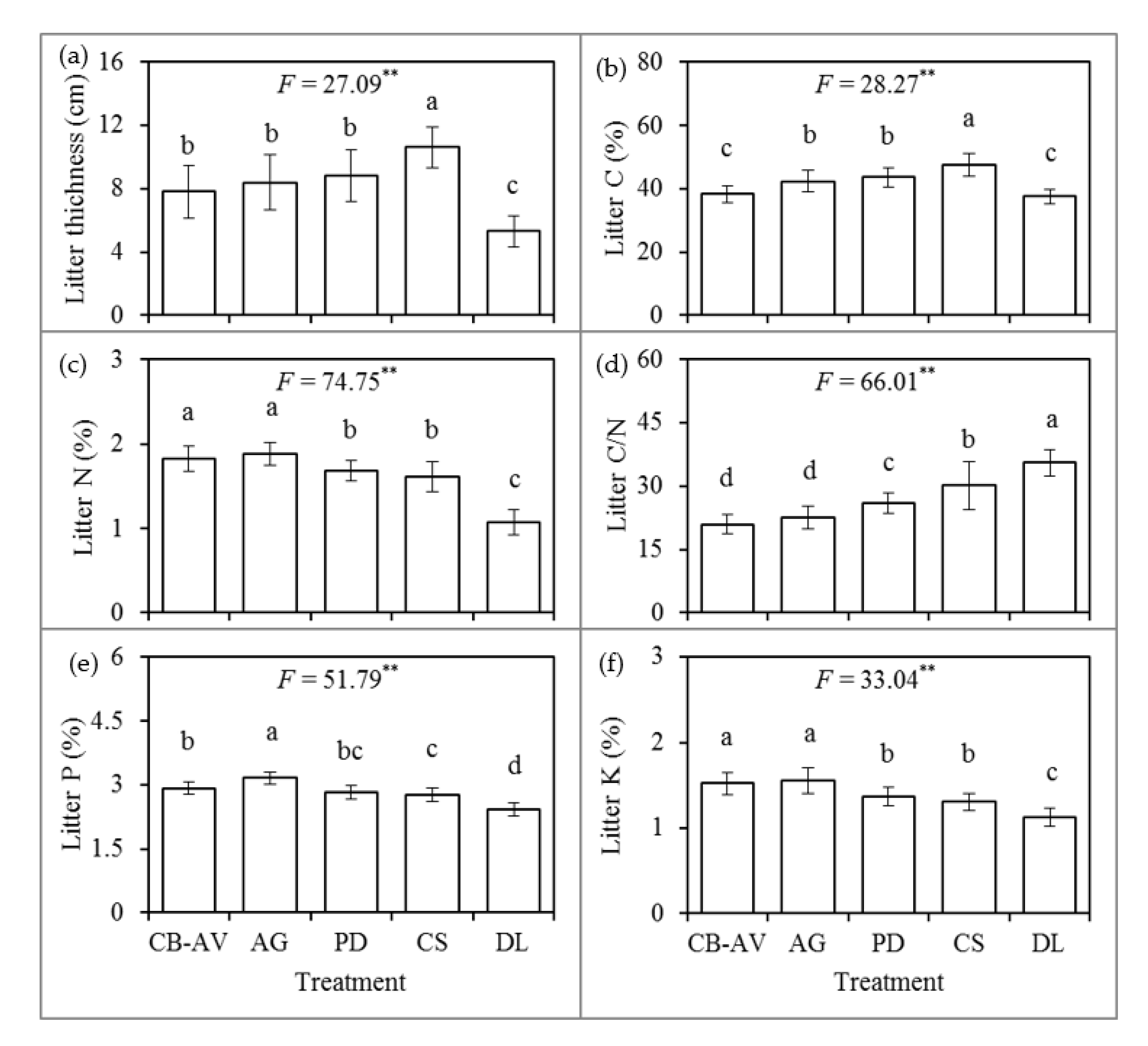
3.1. Litter Properties
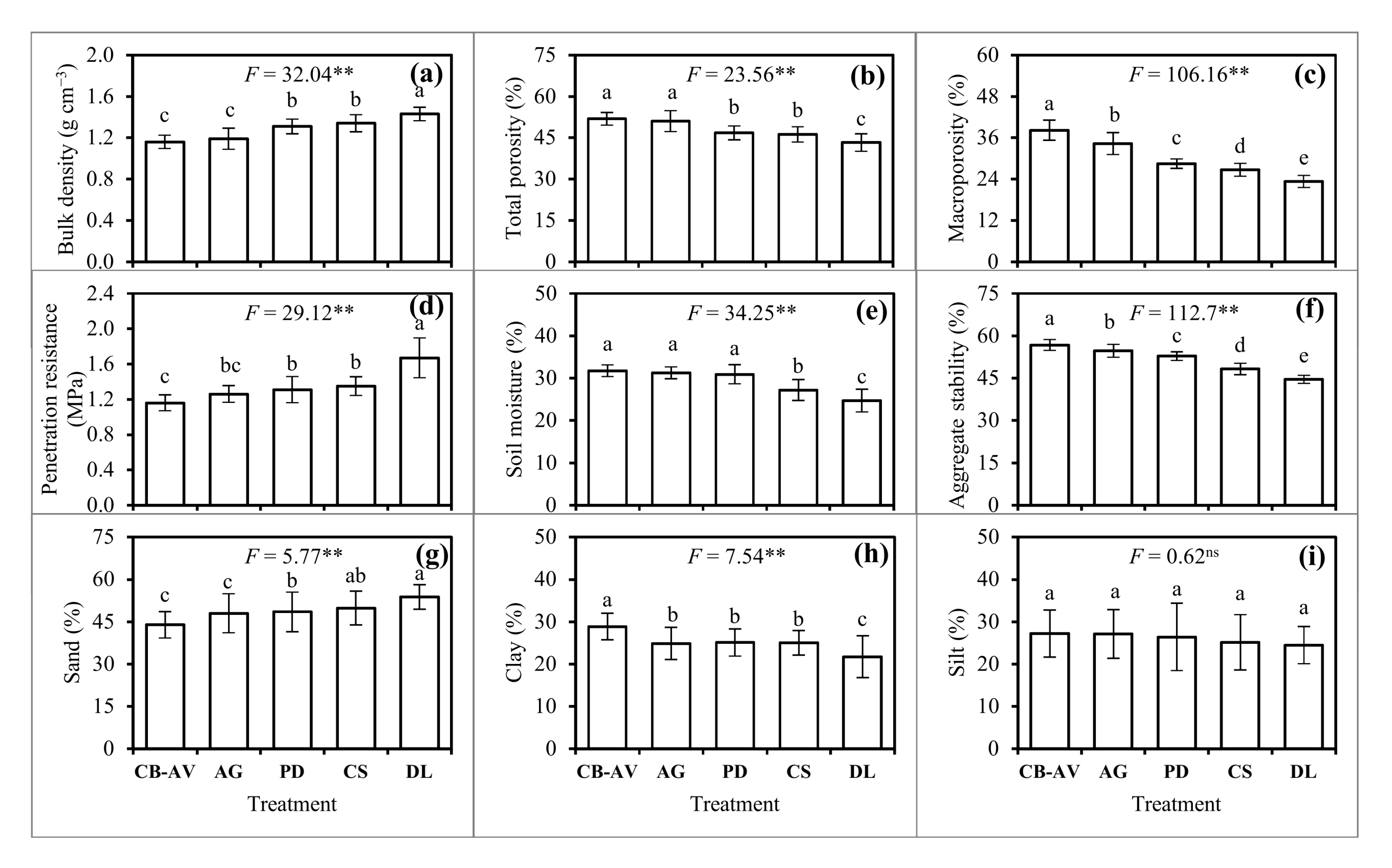
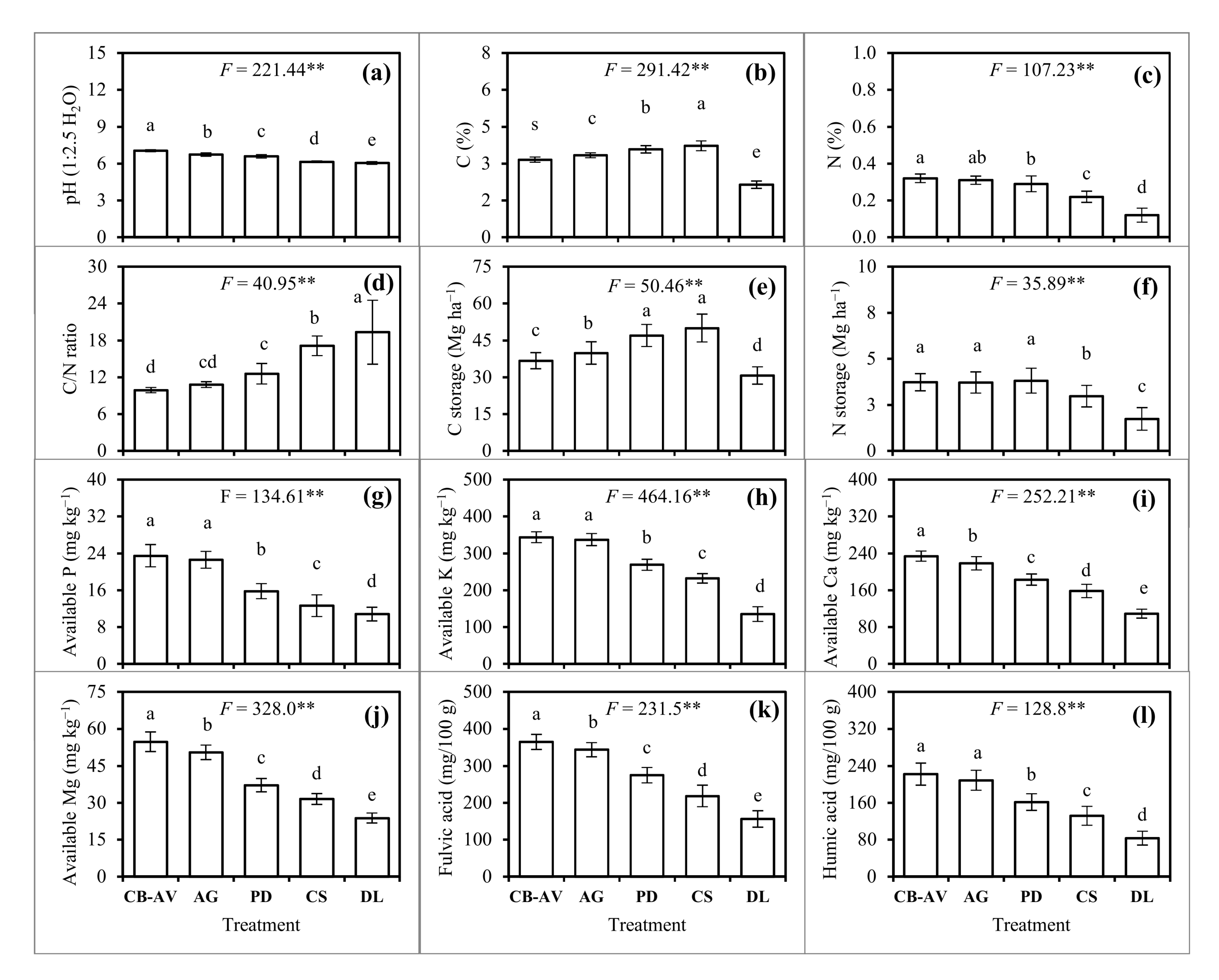
3.2. Soil Physio-Chemical Properties
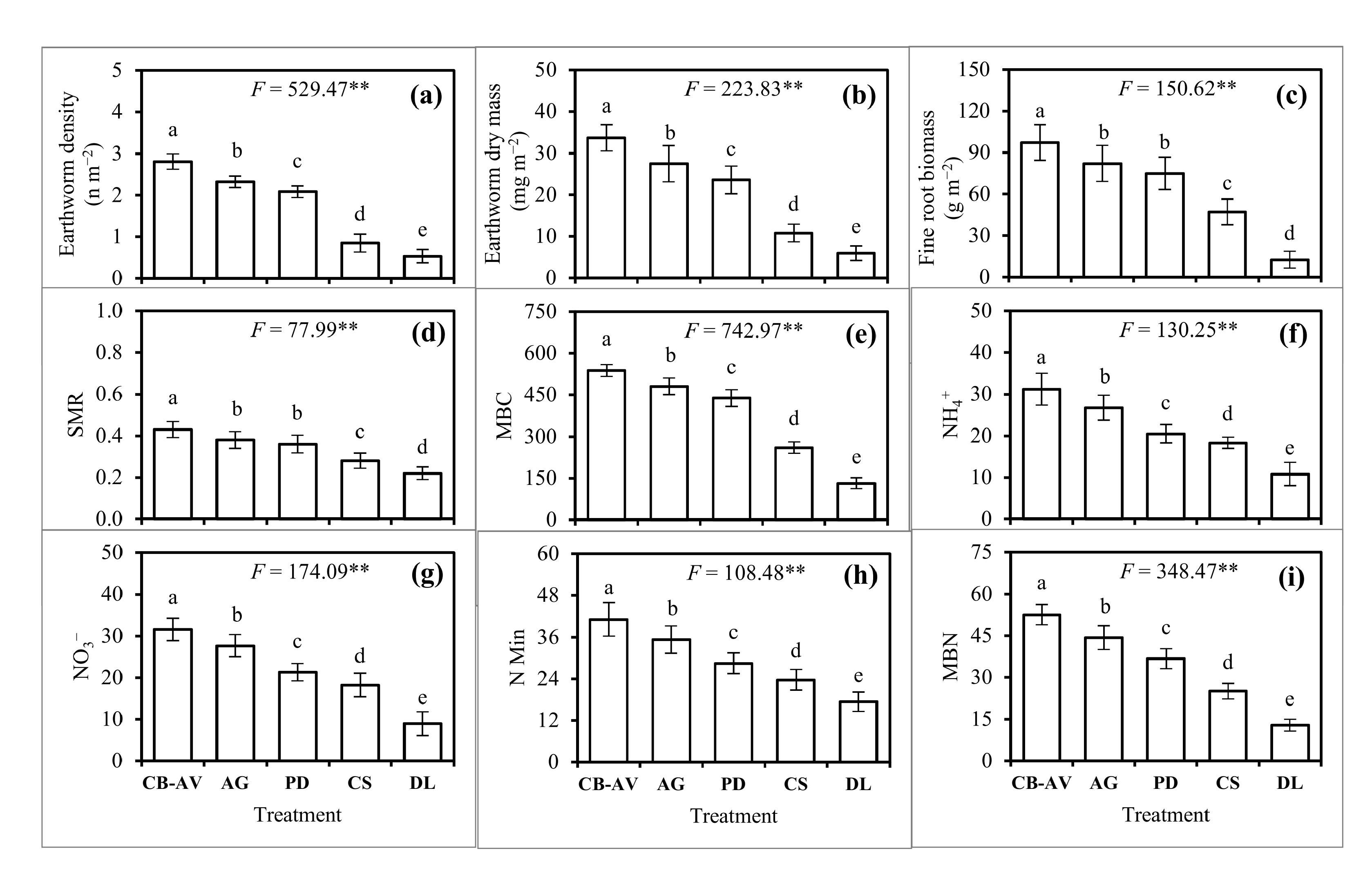
3.3. Soil Biochemical, Biological and Microbial Properties
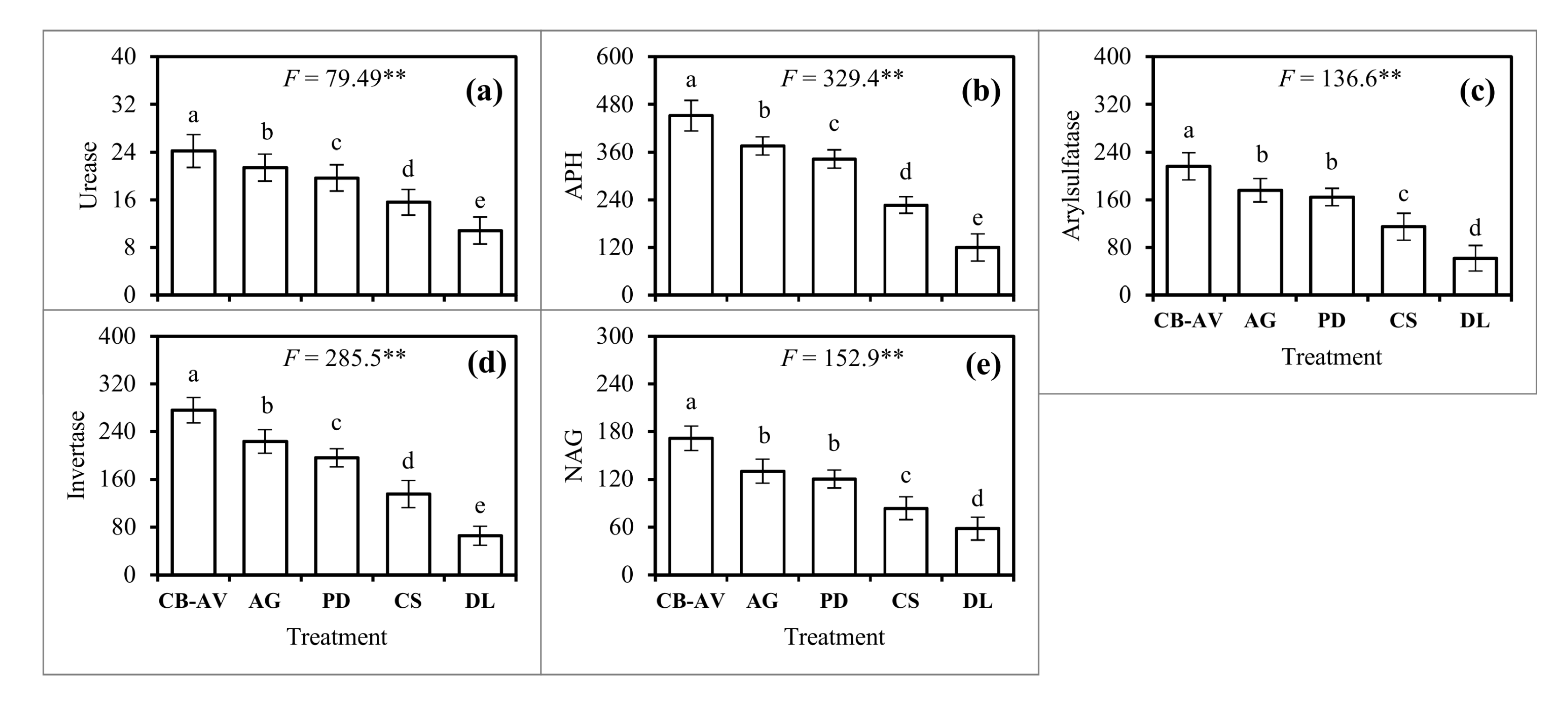
3.4. Enzyme Activitiy
4. Discussion
4.1. Litter Properties
4.2. Soil Physio-Chemical Properties
4.3. Soil Biochemical, Biological, and Microbial Properties and Enzyme Activity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marchi, E.; Chung, W.; Visser, R.; Abbas, D.; Nordfjell, T.; Mederski, P.S.; McEwan, A.; Brink, M.; Laschi, A. Sustainable Forest Operations (SFO): A new paradigm in a changing world and climate. Sci. Total Environ. 2018, 634, 1385–1397. [Google Scholar] [CrossRef] [Green Version]
- Mariotti, B.; Hoshika, Y.; Cambi, M.; Marra, E.; Feng, Z.; Paoletti, E.; Marchi, E. Vehicle-induced compaction of forest soil affects plant morphological and physiological attributes: A meta-analysis. For. Ecol. Manag. 2020, 462, 118004. [Google Scholar] [CrossRef]
- Labelle, E.R.; Hansson, L.; Högbom, L.; Jourgholami, M.; Laschi, A. Strategies to mitigate the effects of soil physical disturbances caused by forest machinery: A Comprehensive Review. Curr. For. Rep. 2022, 8, 20–37. [Google Scholar] [CrossRef]
- Marra, E.; Laschi, A.; Fabiano, F.; Foderi, C.; Neri, F.; Mastrolonardo, G.; Nordfjell, T.; Marchi, E. Impacts of wood extraction on soil: Assessing rutting and soil compaction caused by skidding and forwarding by means of traditional and innovative methods. Eur. J. For. Res. 2022, 141, 71–86. [Google Scholar] [CrossRef]
- Flores Fernández, J.L.; Hartmann, P.; Schäffer, J.; Pulhmann, H.; von Wilpert, K. Initial recovery of compacted soil—Planting and technical treatments decrease CO2 concentrations in soil and promote root growth. Ann. For. Sci. 2017, 74, 73. [Google Scholar] [CrossRef] [Green Version]
- Marra, E.; Wictorsson, R.; Bohlin, J.; Marchi, E.; Nordfjell, T. Remote measuring of the depth of wheel ruts in forest terrain using a drone. Int. J. For. Eng. 2021, 32, 224–234. [Google Scholar] [CrossRef]
- Picchio, R.; Jourgholami, M.; Zenner, E.K. Effects of forest harvesting on water and sediment yields: A Review toward better mitigation and rehabilitation strategies. Curr. For. Rep. 2021, 7, 214–229. [Google Scholar] [CrossRef]
- Sohrabi, H.; Jourgholami, M.; Labelle, E.R. The effect of forest floor on soil microbial and enzyme indices after forest harvesting operations in Hyrcanian deciduous forests. Eur. J. For. Res. 2022, 141, 1013–1027. [Google Scholar] [CrossRef]
- Cambi, M.; Certini, G.; Neri, F.; Marchi, E. The impact of heavy traffic on forest soils: A review. For. Ecol. Manag. 2015, 338, 124–138. [Google Scholar] [CrossRef]
- Cambi, M.; Hoshika, Y.; Mariotti, B.; Paoletti, E.; Picchio, R.; Venanzi, R.; Marchi, E. Compaction by a forest machine affects soil quality and Quercus robur L. seedling performance in an experimental field. For. Ecol. Manag. 2017, 384, 406–414. [Google Scholar] [CrossRef]
- Meyer, C.; Luscher, P.; Schulin, R. Enhancing the regeneration of compacted forest soils by planting black alder in skid lane tracks. Eur. J. For. Res. 2014, 133, 453–465. [Google Scholar] [CrossRef]
- Marra, E.; Cambi, M.; Fernandez-Lacruz, R.; Giannetti, F.; Marchi, E.; Nordfjell, T. Photogrammetric estimation of wheel rut dimensions and soil compaction after increasing numbers of forwarder passes. Scand. J. For. Res. 2018, 33, 613–620. [Google Scholar] [CrossRef]
- Mohieddinne, H.; Brasseur, B.; Spicher, F.; Gallet-Moron, E.; Buridant, J.; Kobaissi, A.; Horen, H. Physical recovery of forest soil after compaction by heavy machines, revealed by penetration resistance over multiple decades. For. Ecol. Manag. 2019, 449, 117472. [Google Scholar] [CrossRef]
- Horn, R.; Vossbrink, J.; Becker, S. Modern forestry vehicles and their impacts on soil physical properties. Soil Tillage Res. 2004, 79, 207–219. [Google Scholar] [CrossRef]
- Bottinelli, N.; Hallaire, V.; Goutal, N.; Bonnaud, P.; Ranger, J. Impact of heavy traffic on soil macroporosity of two silty forest soils: Initial effect and short-term recovery. Geoderma 2014, 217–218, 10–17. [Google Scholar] [CrossRef]
- Sohrabi, H.; Jourgholami, M.; Tavankar, F.; Venanzi, R.; Picchio, R. Post-harvest evaluation of soil physical properties and natural regeneration growth in steep-slope terrains. Forests 2019, 10, 1034. [Google Scholar] [CrossRef] [Green Version]
- Goutal, N.; Boivin, P.; Ranger, J. Assessment of the natural recovery rate of soil specific volume following forest soil compaction. Soil Sci. Soc. Am. J. 2012, 76, 1426–1435. [Google Scholar] [CrossRef]
- Flores Fernández, J.L.; Rubin, L.; Hartmann, P.; Puhlmann, H.; von Wilpert, K. Initial recovery of soil structure of a compacted forest soil can be enhanced by technical treatments and planting. For. Ecol. Manag. 2019, 431, 54–62. [Google Scholar] [CrossRef]
- Aponte, C.; García, L.V.; Marañón, T. Tree species effects on nutrient cycling and soil biota: A feedback mechanism favoring species coexistence. For. Ecol. Manag. 2013, 309, 36–46. [Google Scholar] [CrossRef] [Green Version]
- Lucas-Borja, M.E.; de Santiago, J.H.; Yang, Y.; Shen, Y.; Candel-Pérez, D. Nutrient, metal contents and microbiological properties of litter and soil along a tree age gradient in Mediterranean forest ecosystems. Sci. Total Environ. 2019, 650, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Langenbruch, C.; Helfrich, M.; Flessa, H. Effects of beech (Fagus sylvatica), ash (Fraxinus excelsior) and lime (Tilia spec.) on soil chemical properties in a mixed deciduous forest. Plant Soil 2012, 352, 389–403. [Google Scholar] [CrossRef] [Green Version]
- Diao, M.; Yang, K.; Zhu, J.; Li, M.; Xu, S. Native broad-leaved tree species play key roles on maintaining soil chemical and microbial properties in a temperate secondary forest, Northeast China. For. Ecol. Manag. 2020, 462, 117971. [Google Scholar] [CrossRef]
- Norris, M.; Avis, P.; Reich, P.; Hobbie, S. Positive feedbacks between decomposition and soil nitrogen availability along fertility gradients. Plant Soil 2013, 367, 347–361. [Google Scholar] [CrossRef]
- Prescott, C.E.; Vesterdal, L. Tree species effects on soils in temperate and boreal forests: Emerging themes and research needs. For. Ecol. Manag. 2013, 309, 1–3. [Google Scholar] [CrossRef]
- Jourgholami, M.; Ghassemi, T.; Labelle, E.R. Soil physio-chemical and biological indicators to evaluate the restoration of compacted soil following reforestation. Ecol. Indic. 2019, 101, 102–110. [Google Scholar] [CrossRef]
- Cools, N.; Vesterdal, L.; De Vos, B.; Vanguelova, E.; Hansen, K. Tree species is the major factor explaining C:N ratios in European forest soils. For. Ecol. Manag. 2014, 311, 3–16. [Google Scholar] [CrossRef]
- Marty, C.; Houle, D.; Gagnon, C.; Courchesne, F. The relationships of soil total nitrogen concentrations, pools and C:N ratios with climate, vegetation types and nitrate deposition in temperate and boreal forests of eastern Canada. Catena 2017, 152, 163–172. [Google Scholar] [CrossRef]
- Yang, K.; Shi, W.; Zhu, J.J. The impact of secondary forests conversion into larch plantations on soil chemical and microbiological properties. Plant Soil 2013, 368, 535–546. [Google Scholar] [CrossRef]
- Zhu, L.; Wang, X.; Chen, F.; Li, C.; Wu, L. Effects of the successive planting of Eucalyptus urophylla on soil bacterial and fungal community structure, diversity, microbial biomass, and enzyme activity. Land Degrad. Dev. 2019, 30, 636–646. [Google Scholar] [CrossRef]
- Yang, K.; Zhu, J.J. Impact of tree litter decomposition on soil biochemical properties obtained from a temperate secondary forest in Northeast China. J. Soils Sediments 2015, 15, 13–23. [Google Scholar] [CrossRef]
- Vauramo, S.; Setälä, H. Decomposition of labile and recalcitrant litter types under different plant communities in urban soils. Urban Ecosyst. 2011, 14, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Sagheb-Talebi, K.; Sajedi, T.; Pourhashemi, M. Forests of Iran: A Treasure from the Past, a Hope for the Future; Springer: Berlin, Germany, 2014; pp. 42–152. [Google Scholar]
- Mayer, M.; Prescott, C.E.; Abaker, W.E.A.; Augusto, L.; Cécillon, L.; Ferreira, G.W.D.; James, J.; Jandl, R.; Katzensteiner, K.; Laclau, J.-P.; et al. Tamm review: Influence of forest management activities on soil organic carbon stocks: A knowledge synthesis. For. Ecol. Manag. 2020, 466, 118127. [Google Scholar] [CrossRef]
- WRB IWG. World Reference Base for Soil Resources—A Framework for International Classification, Correlation and Communication; World Soil Resources Reports 103 International Union of Soil Sciences, ISRIC—World Soil Information and Food and Agriculture Organization of the United Nations: Rome, Italy, 2006. [Google Scholar]
- Hattenschwiler, S.; Jørgensen, H.B. Carbon quality rather than stoichiometry controls litter decomposition in a tropical rain forest. J. Ecol. 2010, 98, 754–763. [Google Scholar] [CrossRef]
- Homer, C.D.; Pratt, P.F. Methods of Analysis for Soils, Plants and Waters; University of California, Agricultural Sciences Publications: Berkeley, CA, USA, 1961; p. 309. [Google Scholar]
- Bower, C.A.; Reitemeier, R.F.; Fireman, M. Exchangeable cation analysis of saline and alkali soils. Soil Sci. 1952, 73, 251–261. [Google Scholar] [CrossRef]
- Kemper, W.D.; Rosenau, R.C. Aggregate stability and size distribution. In Methods of Soil Analysis, Part 1: Physical and Mineralogical Methods, 2nd ed.; Klute, A., Ed.; American Society of Agronomy: Madison, WI, USA, 1986; pp. 383–411. [Google Scholar]
- Thien, S.J.; Graveel, J.G. Laboratory Manual for Soil Science: Agricultural & Environmental Principles; Preliminary Edition; Kendall Hunt Publishing Company: Dubuque, IA, USA, 2003; p. 314. [Google Scholar]
- Gee, G.W.; Bauder, J.W. Particle-size analysis. In Methods of Soil Analysis, Part 1. Physical and Mineralogical Methods; Klute, A., Ed.; Soil Science Society of America: Madison, WI, USA, 1986; pp. 383–411. [Google Scholar]
- Danielson, R.E.; Southerland, P.L. Methods of Soil Analysis. Part I. Physical and Mineralogical Methods, 2nd ed.; ASA, SSSA: Madison, WI, USA, 1986; pp. 443–460. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter and a proposed modification of chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Kooch, Y.; Zaccone, C.; Lamersdorf, N.P.; Tonon, G. Pit and mound influence on soil features in an Oriental Beech (Fagus Orientalis Lipsky) forest. Eur. J. For. Res. 2014, 133, 347–354. [Google Scholar] [CrossRef]
- Neatrour, M.A.; Jones, R.H.; Golladay, S.W. Correlations between soil nutrient availability and fine-root biomass at two spatial scales in forested wetlands with contrasting hydrological regimes. Can. J. For. Res. 2005, 35, 2934–2941. [Google Scholar] [CrossRef]
- Alef, K. Soil respiration. In Methods in Applied Soil Microbiology and Biochemistry; Alef, K., Nanninipieri, P., Eds.; Academic Press: London, UK, 1995; pp. 214–219. [Google Scholar]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Yang, Y.; Geng, Y.; Zhou, H.; Zhao, G.; Wang, L. Effects of gaps in the forest canopy on soil microbial communities and enzyme activity in a Chinese pine forest. Pedobiologia 2017, 61, 51–60. [Google Scholar] [CrossRef]
- Robertson, G.P. Standard Soil Methods for Long-Term Ecological Research; Oxford University Press on Demand: Oxford, UK, 1999; Volume 2. [Google Scholar]
- Schinner, F.; von Mersi, W. Xylanase-, CM-cellulase- and invertase activity in soil: An improved method. Soil Biol. Biochem. 1990, 22, 511–515. [Google Scholar] [CrossRef]
- Picchio, R.; Mederski, P.S.; Tavankar, F. How and how much, do harvesting activities affect forest soil, regeneration and stands? Curr. For. Rep. 2020, 6, 115–118. [Google Scholar] [CrossRef] [Green Version]
- Kader, M.A.; Senge, M.; Mojid, M.A.; Ito, K. Recent advances in mulching materials and methods for modifying soil environment. Soil Tillage Res. 2017, 168, 155–166. [Google Scholar] [CrossRef]
- Zhao, L.; Hou, R.; Fang, Q. Differences in interception storage capacities of undecomposed broad-leaf and needle-leaf litter under simulated rainfall conditions. For. Ecol. Manag. 2019, 446, 135–142. [Google Scholar] [CrossRef]
- Sayer, E.J. Using experimental manipulation to assess the roles of leaf litter in the functioning of forest ecosystems. Biol. Rev. 2006, 81, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Giweta, M. Role of litter production and its decomposition, and factors affecting the processes in a tropical forest ecosystem: A review. J. Ecol. Environ. 2020, 44, 11. [Google Scholar] [CrossRef]
- Picchio, R.; Mercurio, R.; Venanzi, R.; Gratani, L.; Giallonardo, T.; Monaco, A.L.; Frattaroli, A.R. Strip clear-cutting application and logging typologies for renaturalization of pine afforestation—A case study. Forests 2018, 9, 366. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.C.; Li, Z.B.; Li, P. Fractal features of soil particle-size distribution and total soil nitrogen distribution in a typical watershed in the source area of the middle Dan River, China. Catena 2013, 101, 17–23. [Google Scholar] [CrossRef]
- Jourgholami, M.; Etehadi Abari, M. Effectiveness of sawdust and straw mulching on postharvest runoff and soil erosion of a skid trail in a mixed forest. Eco. Eng. 2017, 109, 15–24. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Cornlissen, J.H.C.; Amatangelo, K.; Dorrrepaal, E.; Eviner, V.T.; Godoy, O.; Hobbie, S.E.; Hoorens, B.; Kurokawa, H.; Pérez-Harguindeguy, N.; et al. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 2008, 11, 1065–1071. [Google Scholar] [CrossRef]
- Berg, B.; Laskowski, R. Litter decomposition: A guide to carbon and nutrient turnover. Adv. Ecol. Res. 2006, 38, 448. [Google Scholar]
- Krishna, M.P.; Mohan, M. Litter decomposition in forest ecosystems: A review. Energy Ecol. Environ. 2017, 2, 236–249. [Google Scholar] [CrossRef]
- Jourgholami, M.; Labelle, E.R. Effects of plot length and soil texture on runoff and sediment yield occurring on machine-trafficked soils in a mixed deciduous forest. Ann. For. Sci. 2020, 77, 19. [Google Scholar] [CrossRef]
- Hagen-Thorn, A.; Callesen, I.; Armolaitis, K.; Nihlgard, B. The impact of six European tree species on the chemistry of mineral topsoil in forest plantations on former agricultural land. For. Ecol. Manag. 2004, 195, 373–384. [Google Scholar] [CrossRef]
- Mueller, K.E.; Hobbie, S.E.; Chorover, J.; Reich, P.B.; Eisenhauer, N.; Castellano, M.J.; Chadwick, O.A.; Dobies, T.; Hale, C.M.; Jagodziński, A.M.; et al. Effects of litter traits, soil biota, and soil chemistry on soil carbon stocks at a common garden with 14 tree species. Biogeochemistry 2015, 123, 313–327. [Google Scholar] [CrossRef]
- Jourgholami, M.; Khajavi, S.; Labelle, E.R. Mulching and water diversion structures on skid trails: Response of soil physical properties six years after harvesting. Ecol. Eng. 2018, 123, 1–9. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C. Climatic environment. In Plant Litter; McClaugherty, C., Berg, B., Eds.; Springer: Berlin Heidelberg, Germany, 2014; pp. 143–170. [Google Scholar]
- García-Palacios, P.; Maestre, F.T.; Kattge, J.; Wall, D.H. Climate and litter quality differently modulate the effects of soil fauna on litter decomposition across biomes. Ecol. Lett. 2013, 16, 1045–1053. [Google Scholar] [CrossRef] [Green Version]
- Guo, D.; Xia, M.; Wei, X.; Chang, W.; Liu, Y.; Wang, Z. Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species. New Phytol. 2008, 180, 673–683. [Google Scholar] [CrossRef]
- Yang, X.; Shao, M.; Li, T. Effects of terrestrial isopods on soil nutrients during litter decomposition. Geoderma 2020, 376, 114546. [Google Scholar] [CrossRef]
- Parsons, S.A.; Valdez-Ramirez, V.; Congdon, R.A.; Williams, S.E. Contrasting patterns of litterfall seasonality and seasonal changes in litter decomposability in a tropical rainforest region. Biogeosciences 2014, 11, 5047–5056. [Google Scholar] [CrossRef] [Green Version]
- Chapman, S.K.; Koch, G.W. What type of diversity yields synergy during mixed litter decomposition in a natural forest ecosystem? Plant Soil 2007, 299, 153–162. [Google Scholar] [CrossRef]
- Chalker-Scott, L. Impact of mulches on landscape plants and the environment-a review. J. Environ. Hortic. 2007, 25, 239–249. [Google Scholar] [CrossRef]
- Salehi, M.; Zahedi Amiri, G.; Attarod, P.; Salehi, A.; Brunner, I.; Schleppi, P.; Thimonier, A. Seasonal variations of throughfall chemistry in pure and mixed stands of Oriental beech (Fagus orientalis Lipsky) in Hyrcanian forests (Iran). Ann. For. Sci. 2016, 73, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Lohse, K.A.; Brooks, P.D.; Mcintosh, J.C.; Meixner, T.; Huxman, T.E. Interactions between biogeochemistry and hydrologic systems. Annu. Rev. Environ. Resour. 2009, 34, 65–96. [Google Scholar] [CrossRef]
- Fang, S.; Xie, B.; Liu, D.; Liu, J. Effects of mulching materials on nitrogen mineralization, nitrogen availability and poplar growth on degraded agricultural soil. New For. 2011, 41, 147–162. [Google Scholar] [CrossRef]
- Zhang, D.; Zheng, H.; Ren, Z.; Zhai, C.; Shen, G.; Mao, Z.; Wang, P.; He, X. Effects of forest type and urbanization on carbon storage of urban forests in Changchun, Northeast China. Chin. Geogr. Sci. 2015, 25, 147–158. [Google Scholar] [CrossRef]
- Perie, C.; Ouimet, R. Organic carbon, organic matter and bulk density relationships in boreal forest soils. Can. J. Soil Sci. 2007, 88, 315–325. [Google Scholar] [CrossRef]
- Huang, W.; Spohn, M. Effects of long-term litter manipulation on soil carbon, nitrogen, and phosphorus in a temperate deciduous forest. Soil Biol. Biochem. 2015, 83, 12–18. [Google Scholar] [CrossRef]
- Xiong, Y.; Zeng, H.; Xia, H.; Guo, D. Interactions between leaf litter and soil organic matter on carbon and nitrogen mineralization in six forest litter-soil systems. Plant Soil 2014, 379, 217–229. [Google Scholar] [CrossRef]
- Hoogmoed, M.; Cunningham, S.C.; Baker, P.J.; Beringer, J.; Cavagnaro, T.R. Is there more soil carbon under nitrogen-fixing trees than under non-nitrogen-fixing trees in mixed-species restoration plantings? Agric. Ecosyst. Environ. 2014, 188, 80–84. [Google Scholar] [CrossRef]
- Guckland, A.; Jacob, M.; Flessa, H.; Thomas, F.M.; Leuschner, C. Acidity, nutrient stocks, and organic-matter content in soils of a temperate deciduous forest with different abundance of European beech (Fagus sylvatica L.). J. Plant Nutr. Soil Sci. 2009, 172, 500–511. [Google Scholar] [CrossRef]
- Augusto, L.; Ranger, J.; Binkley, D.; Rothe, A. Impact of several common tree species of European temperate forests on soil fertility. Ann. For. Sci. 2002, 59, 233–253. [Google Scholar] [CrossRef] [Green Version]
- Jourgholami, M.; Khajavi, S.; Labelle, E.R. Recovery of forest soil chemical properties following soil rehabilitation treatments: An assessment six years after machine impact. Croat. J. For. Eng. 2020, 41, 163–175. [Google Scholar] [CrossRef] [Green Version]
- Deng, H.; Shen, L.; Yang, J.; Mo, X. Stand stability of pure and mixed-eucalyptus forests of different tree species in a typhoon-prone area. Forests 2021, 12, 458. [Google Scholar] [CrossRef]
- Gao, Z.; Li, J.; Zhu, H.; Sun, L.; Du, Y.; Zhai, H. Using differential thermal analysis to analyze cold hardiness in the roots of grape varieties. Sci. Hortic. 2014, 174, 155–163. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Main Species | Slope (%) | Aspect | Tree Density (N ha−1) | Growing Stock (m3 ha−1) | Diameter at Breast Height (cm) |
|---|---|---|---|---|---|---|
| CB-AV | Hornbeam (Carpinus betulus L.)-Velvet maple (Acer velutinum Boiss.) | 15 ± 5 | Northeast | 364.1 ± 41.7 | 286.3 ± 45.2 | 52.6 ± 9.4 |
| AG | Black alder (Alnus glutinosa (L.) Gaertn.) | 17 ± 3 | Northeast | 614.8 ± 45.3 | 394.1 ± 33.6 | 25.3 ± 6.2 |
| PD | Eastern cottonwood (Populus deltoides L.) | 17 ± 2 | Northeast | 587.6 ± 43.8 | 362.5 ± 39.5 | 24.2 ± 5.3 |
| AC | Italian cypress (Cupressus sempervirens L. var. horizontalis (Mill.) Gord.) | 16 ± 3 | Northeast | 531.8 ± 52.6 | 269.4 ± 51.7 | 20.6 ± 7.4 |
| DL | Degraded land without trees | 18 ± 4 | Northeast | - | - | - |
| Layer | Group | Properties | Unit | Method | Reference for Method |
|---|---|---|---|---|---|
| Litter | Physical and chemical properties | Litter depth or thickness | cm | Tape measure | Sohrabi et al. [8] |
| C and N | % | The CN elemental analyzer | Hattenschwiler and Jørgensen [35] | ||
| P | % | Olsen method | Homer and Pratt [36] | ||
| K | % | Atomic absorption spectrophotometer | Bower et al. [37] | ||
| Soil | Physical properties | Soil bulk density | g cm−3 | Clod method | Kemper and Rosenau [38] |
| Soil moisture | % | By drying soil samples at 105 °C for 24 h | Thien and Graveel [39] | ||
| Soil particle size distribution | % | Hydrometer method | Gee and Bauder [40] | ||
| Soil particle density | g cm−3 | ASTM D854-00 2000 standard | Thien and Graveel [39] | ||
| Macroporosity | % | Water desorption method | Danielson and Sutherland [41] | ||
| Penetration resistance | MPa | Analog hand-held soil penetrometer | Sohrabi et al. [8] | ||
| Total porosity | % | was calculated using formula | Sohrabi et al. [8] | ||
| Aggregate stability | % | Yoder method | Kemper and Rosenau [38] | ||
| Chemical properties | pH | 1:2.5 H2O | Using an Orion Ionalyzer Model 901 pH meter | Sohrabi et al. [8] | |
| C | % | Walkley-Black technique | Walkley and Black [42] | ||
| N | % | The Kjeldahl method | Kooch et al. [43] | ||
| C and N storage | Mg ha−1 | calculated using the bulk density data, C and N concentrations | Kooch et al. [43] | ||
| Available P | mg kg−1 | Olsen method | Homer and Pratt [36] | ||
| Available K, Ca, Mg | mg kg−1 | Determined with an atomic absorption spectrophotometer | Bower et al. [37] | ||
| Fulvic and Humic acid | mg/100 g | The method of the International Humic Substances Society | Sparks and Bartels [43] |
| Layer | Group | Properties | Unit | Method | Reference for Method |
|---|---|---|---|---|---|
| Soil | Biological properties | Earthworm density | n m−2 | By hand sorting | Kooch et al. [43] |
| Earthworm dry mass | mg m−2 | oven dried at 60 °C for 24 h | Kooch et al. [43] | ||
| Fine root biomass | g m−2 | Oven dried at 70 °C to a constant mass | Neatrour et al. [44] | ||
| Soil microbial respiration | mg CO2-C g soil−1 day−1 | Determined by measuring the CO2 evolved in a 3-day incubation experiment at 25 °C | Alef [45] | ||
| C and N Microbial properties | Microbial biomass carbon (MBC) and nitrogen (MBN) | mg kg−1 | Measured by fumigation-extraction method | Brookes et al. [46] | |
| Ammonium and Nitrate | mg kg−1 | Colorimetric techniques were used to extract soil and NO3− via 2 M KCl solution (soil: solution, 1:5) | Yang et al. [47] | ||
| Nitrogen mineralization | mg N kg soil−1 | Aerobic incubation of the soils was applied | Robertson [48] | ||
| Enzyme activity | Urease | –Ng−1 2 h−1 | Analyzed using 200 µmol urea as substrate, incubated for 2 h at 37 °C | Yang et al. [47] | |
| Acid phosphatase (APH) | μg PNP g−1 h−1 | Determined in a MUB buffer (pH 6.5), incubated for 1 h at 37 °C | Yang et al. [47] | ||
| Arylsulfatase | μg PNP g−1 h−1 | A p-Nitrophenyl sulphate was used for incubation for 1 h at 37 °C | Schinner and von Mersi [49] | ||
| Invertase | μg Glucose g−1 3 h−1 | 1.2% sucrose solution was used for incubation at 3 h at 50 °C | Schinner and von Mersi [49] | ||
| ß-N-acetylglucosaminidase (NAG) | μg g−1 h−1 | Analyzed in 100 µmol acetate buffer at pH 5.5 | Yang et al. [47] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramineh, A.; Jourgholami, M.; Etemad, V.; Jafari, M.; Picchio, R. Effect of Different Vegetation Restoration on Recovery of Compaction-Induced Soil Degradation in Hyrcanian Mixed Forests: Influence on Soil C and N Pools and Enzyme Activities. Forests 2023, 14, 603. https://doi.org/10.3390/f14030603
Ramineh A, Jourgholami M, Etemad V, Jafari M, Picchio R. Effect of Different Vegetation Restoration on Recovery of Compaction-Induced Soil Degradation in Hyrcanian Mixed Forests: Influence on Soil C and N Pools and Enzyme Activities. Forests. 2023; 14(3):603. https://doi.org/10.3390/f14030603
Chicago/Turabian StyleRamineh, Alireza, Meghdad Jourgholami, Vahid Etemad, Mohammad Jafari, and Rodolfo Picchio. 2023. "Effect of Different Vegetation Restoration on Recovery of Compaction-Induced Soil Degradation in Hyrcanian Mixed Forests: Influence on Soil C and N Pools and Enzyme Activities" Forests 14, no. 3: 603. https://doi.org/10.3390/f14030603
APA StyleRamineh, A., Jourgholami, M., Etemad, V., Jafari, M., & Picchio, R. (2023). Effect of Different Vegetation Restoration on Recovery of Compaction-Induced Soil Degradation in Hyrcanian Mixed Forests: Influence on Soil C and N Pools and Enzyme Activities. Forests, 14(3), 603. https://doi.org/10.3390/f14030603