
How Climate Change Impacts the Distribution of Lithocarpus hancei (Fagaceae), a Dominant Tree in East Asian Montane Cloud Forests

Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Area
2.2. Data Collection
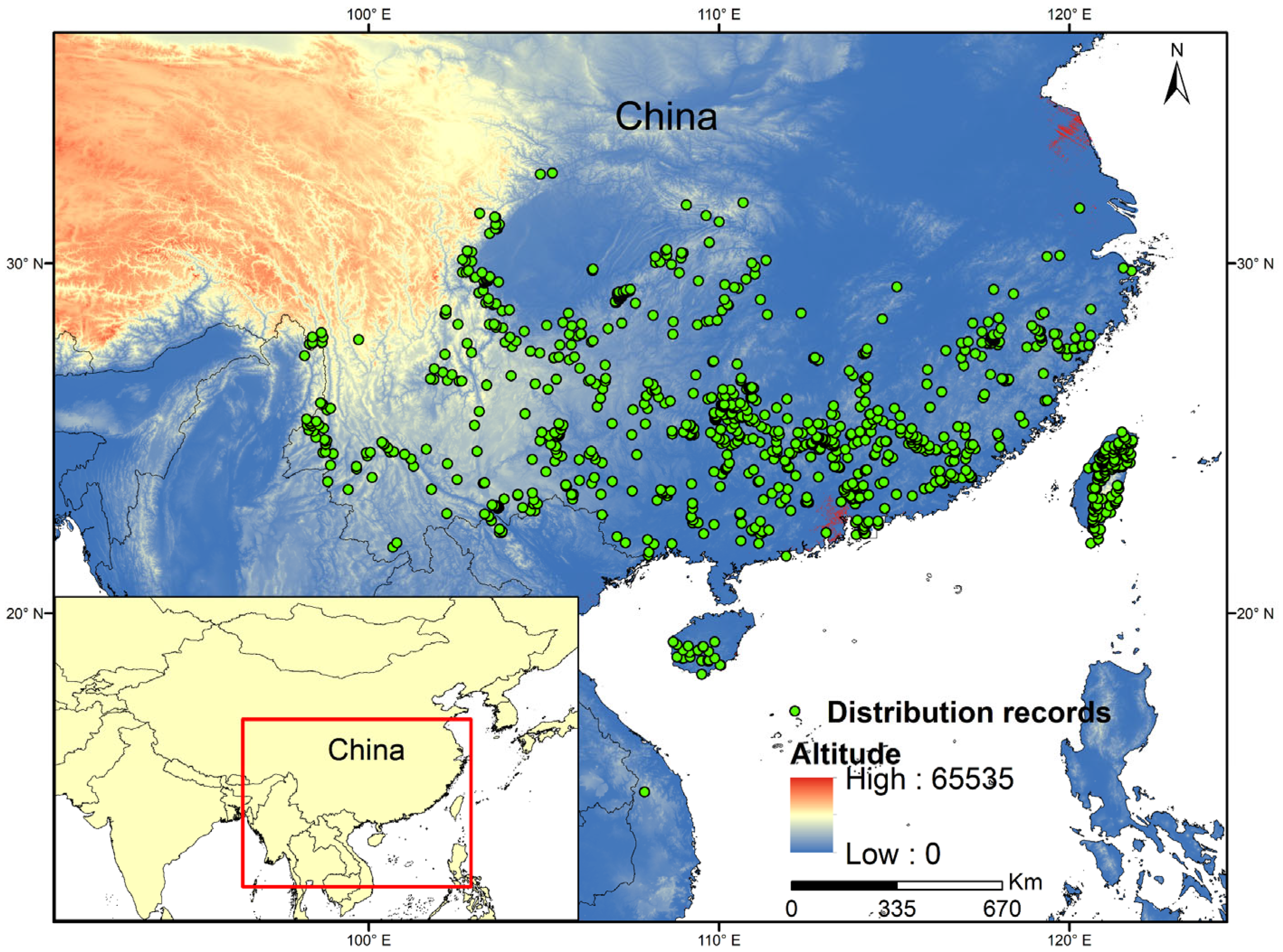
2.2.1. Occurrence Data and Data Filtering Process
2.2.2. Environmental Variables
2.3. Variable Selection
2.4. Model Evaluation and Modeling Simulation
2.5. Analysis of Habitat Change and Range Contraction or Expansion
2.6. Centroid Change
3. Results
3.1. Model Performance
3.2. Environmental Variable Contribution and Response Curves
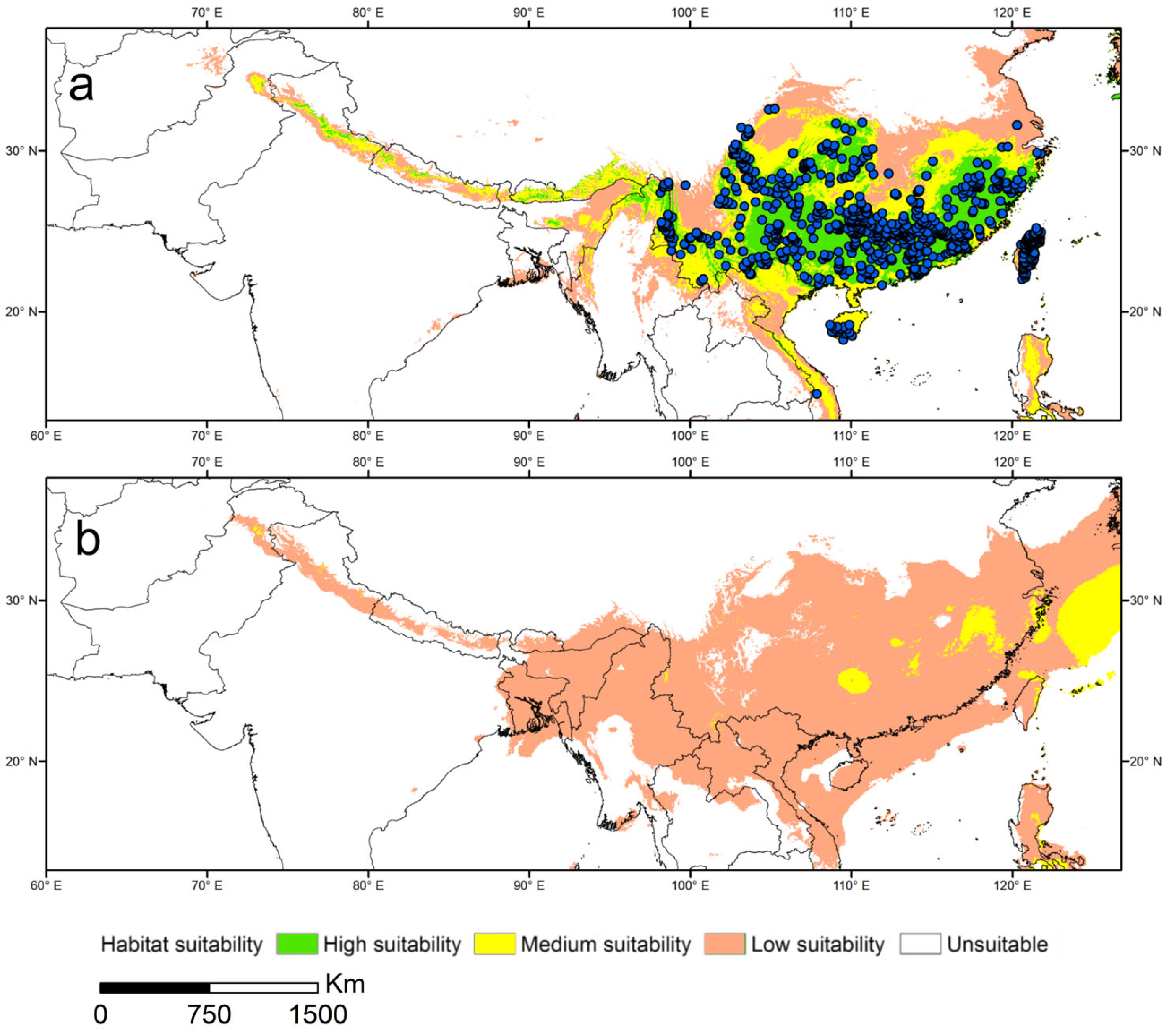
3.3. Current Potential Distribution of Suitable Areas for L. hancei
3.4. Potential Distribution of Suitable Areas for L. hancei during the LGM
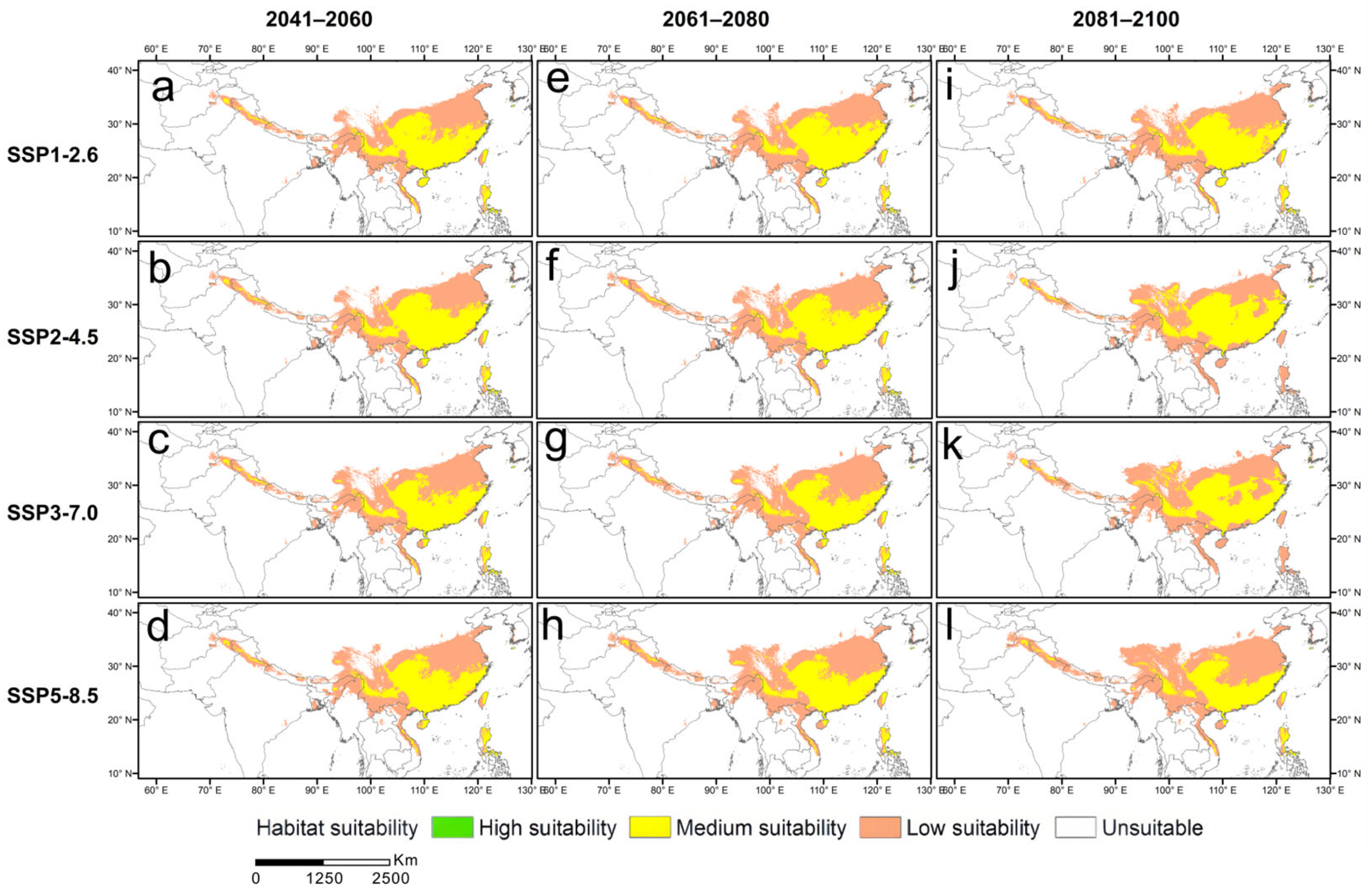
3.5. Potential Distribution of Suitable Areas for L. hancei in the Future
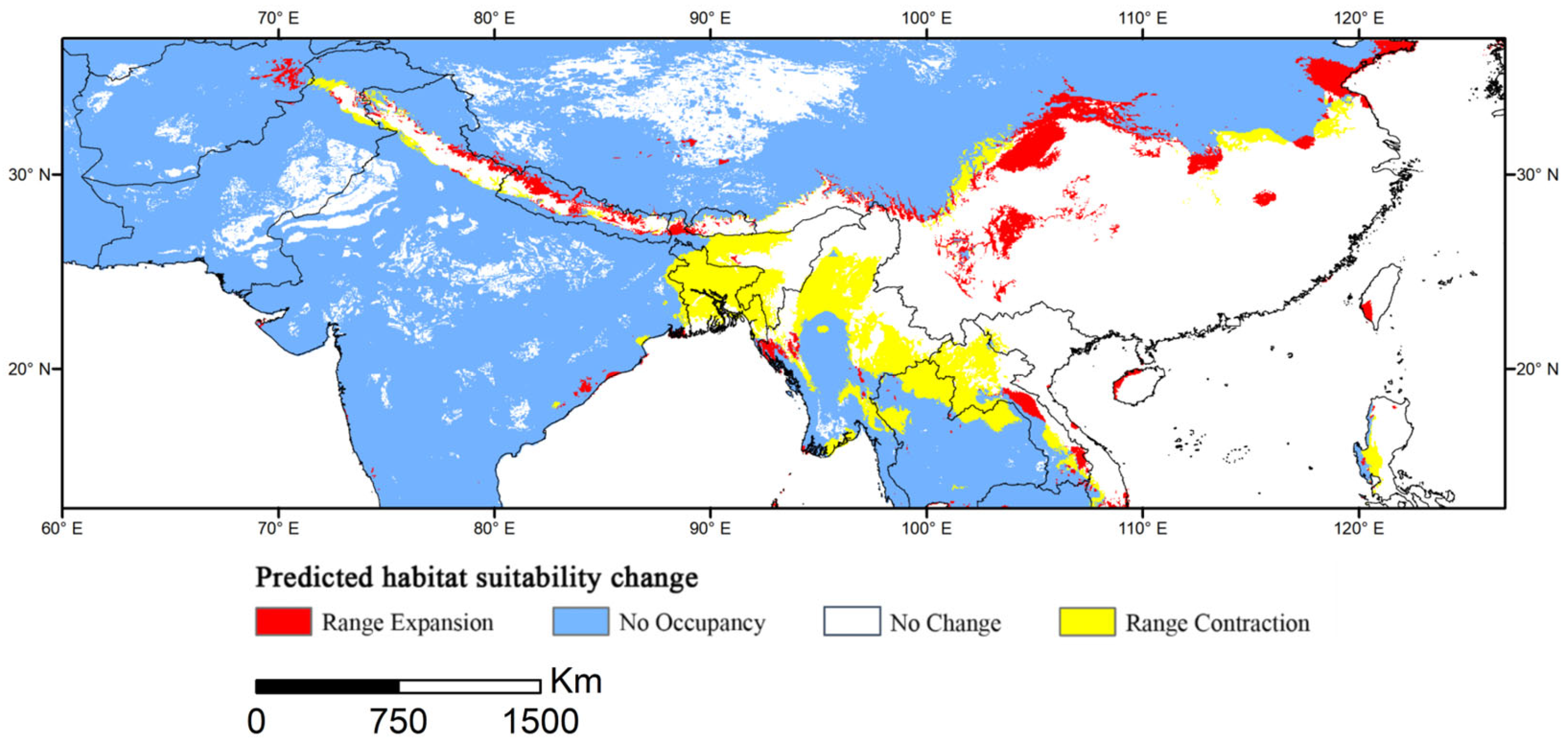
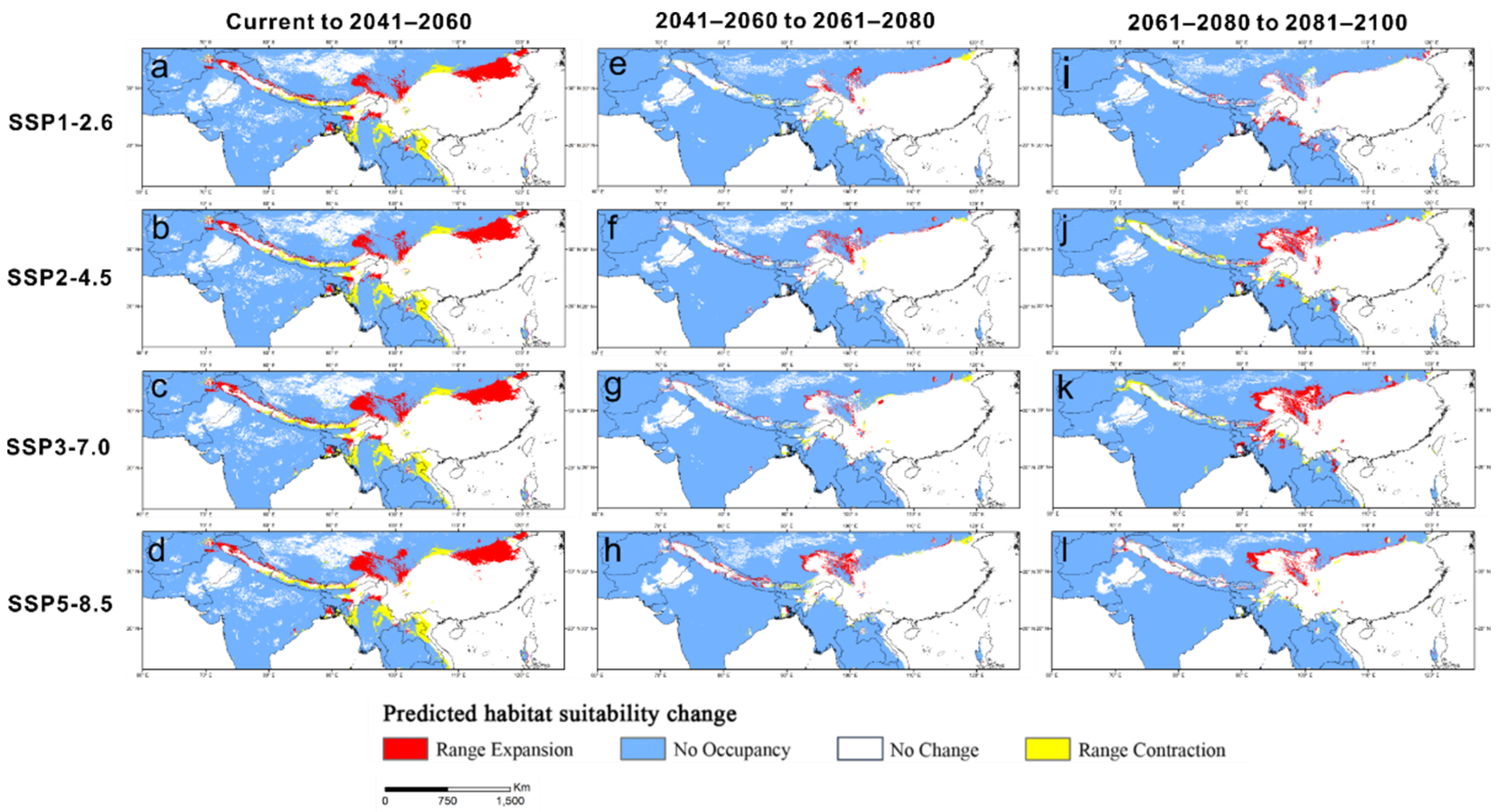
3.6. Spatial Delineation of Range Contraction or Expansion
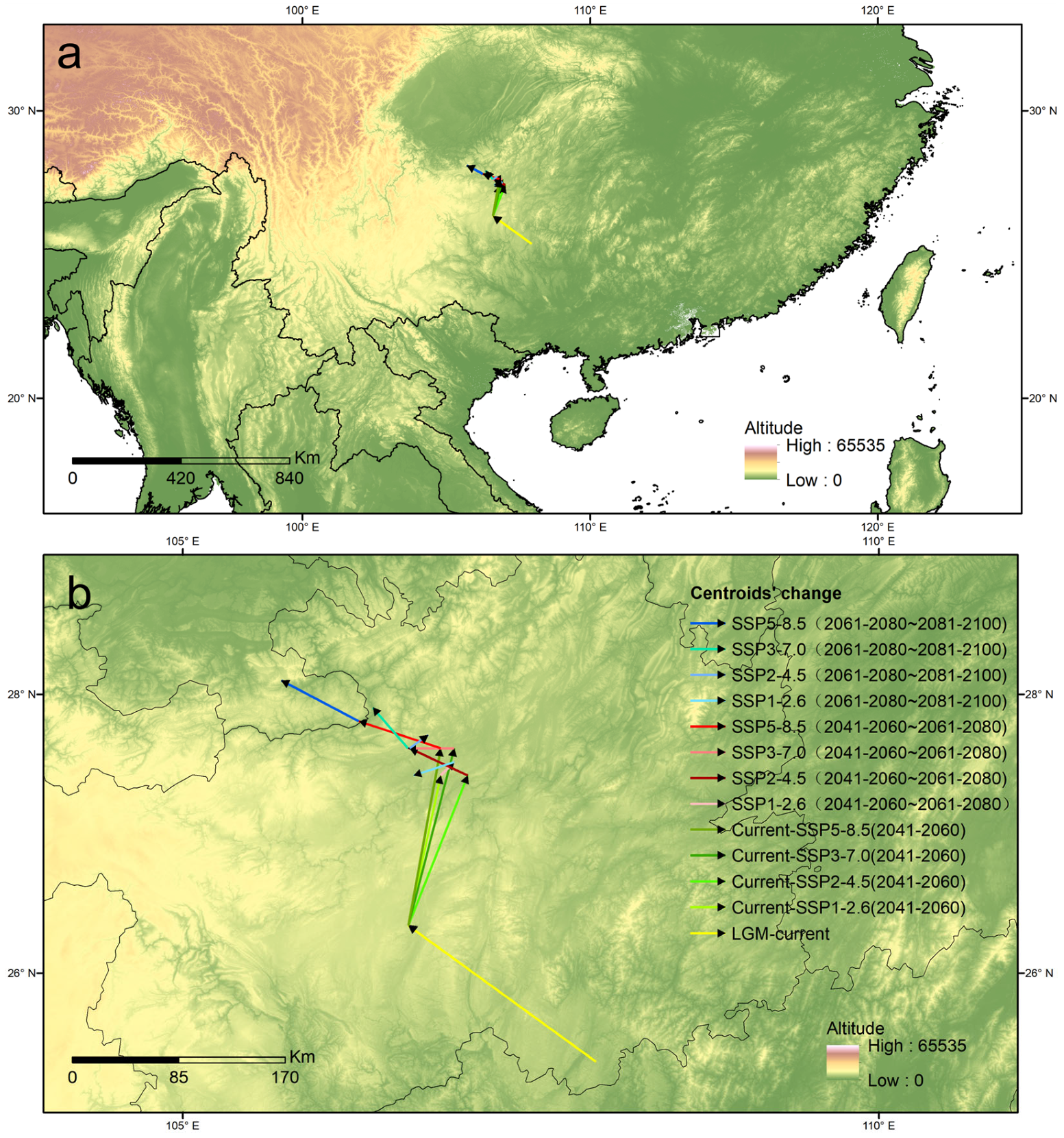
3.7. Shifts in the Distribution Centroid
4. Discussion
4.1. The Main Environmental Variables Restricting the Distribution of L. hancei
4.2. Suitable Habitats for L. hancei under the Current Climate
4.3. Changes in the Suitable Habitats of L. hancei under Future Climate Scenarios
4.4. Conservation and Forestry Management of East Asia MMEBFs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fang, J.Y.; Ohsawa, M.; Kira, T. Vertical vegetation zones along 30° N latitude in humid East Asia. Vegetatio 1996, 126, 135–149. [Google Scholar] [CrossRef]
- Wang, X.H.; Kent, M.; Fang, X.F. Evergreen broad-leaved forest in Eastern China: Its ecology and conservation and the importance of resprouting in forest restoration. For. Ecol. Manag. 2007, 245, 76–87. [Google Scholar] [CrossRef]
- Wu, Z.Y.; Zhu, Y.C.; Jiang, H.Q. Vegetation of Yunnan; Science Press: Beijing, China, 1987; pp. 275–279. [Google Scholar]
- Song, L.; Yang, B.; Liu, L.L.; Mo, Y.X.; Liu, W.J.; Meng, X.J.; Lu, H.Z.; Li, Y.; Zakari, S.; Tan, Z.H.; et al. Spatial-temporal differentiations in water use of coexisting trees from a subtropical evergreen broadleaved forest in Southwest China. Agric. For. Meteorol. 2022, 316, 108862. [Google Scholar] [CrossRef]
- Huang, C.C.; Chang, Y.T.; Bartholomew, B. Fagaceae. In Flora of China; Wu, C.Y., Raven, P.H., Eds.; Science Press and Missouri Botanical Garden Press: Beijing, China, 1999; Volume 22, pp. 176–180. [Google Scholar]
- Cao, K.F. Morphology and growth of deciduous and evergreen broad-leaved saplings under different light conditions in a Chinese beech forest with dense bamboo undergrowth. Ecol. Res. 2001, 16, 509–517. [Google Scholar] [CrossRef]
- The Editorial Committee of Vegetation of China. Vegetation of China; Science Press: Beijing, China, 1980; pp. 316–318. [Google Scholar]
- Li, G.C.; Han, X.G.; Huang, J.H. Dry-season dynamics of soil inorganic nitrogen pools in primary Lithocarpus xylocarpus forest and degraded vegetation in Ailao Mountain, Yunnan province. Chin. J. Plant Ecol. 2001, 25, 210–217. [Google Scholar]
- Song, L.; Liu, W.Y.; Nadkarni, N.M. Response of non-vascular epiphytes to simulated climate change in a montane moist evergreen broad-leaved forest in southwest China. Biol. Conserv. 2012, 152, 127–135. [Google Scholar] [CrossRef]
- Tang, C.Q. The Subtropical Vegetation of Southwestern China: Plant Distribution, Diversity and Ecology; Springer: Dordrecht, The Netherlands, 2015; pp. 339–342. [Google Scholar]
- Williams-Linera, G. Tree species richness complementarity, disturbance and fragmentation in a Mexican tropical montane cloud forest. Biodivers. Conserv. 2002, 11, 1825–1843. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Svenning, J.C.; Fitzpatrick, M.C.; Normand, S.; Graham, C.H.; Pearman, P.B.; Iverson, L.R.; Skov, F. Geography, topography, and history affect realized-to-potential tree species richness patterns in Europe. Ecography 2010, 33, 1070–1080. [Google Scholar] [CrossRef]
- Paul, J.R.; Morton, C.; Taylor, C.M.; Tonsor, S.J. Evolutionary time for dispersal Limits the Extent but Not the Occupancy of Species’ Potential Ranges in the Tropical Plant Genus Psychotria (Rubiaceae). Am. Nat. 2009, 173, 188–199. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- Franklin, J. Moving beyond static species distribution models in support of conservation biogeography. Divers. Distrib. 2010, 16, 321–330. [Google Scholar] [CrossRef]
- Li, G.Q.; Liu, C.C.; Liu, Y.G.; Yang, J.; Zhang, X.S.; Guo, K. Advances in theoretical issues of species distribution models. Acta Ecol. Sin. 2013, 33, 4827–4835. [Google Scholar] [CrossRef]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Model. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J.; Hijmans, M.R. Package ‘dismo’. Circles 2017, 9, 1–68. [Google Scholar]
- R Core Team, R. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Braconnot, P.; Otto-Bliesner, B.; Harrison, S.; Joussaume, S.; Peterchmitt, J.Y.; Abe-Ouchi, A.; Crucifix, M.; Driesschaert, E.; Fichefet, T.; Hewitt, C.D. Results of PMIP2 coupled simulations of the Mid-Holocene and Last Glacial Maximum—Part 1: Experiments and large-scale features. Clim. Past 2007, 3, 261–277. [Google Scholar] [CrossRef]
- Rana, S.K.; Rana, H.K.; Luo, D.; Sun, H. Estimating climate-induced ‘Nowhere to go’ range shifts of the Himalayan incarvillea Juss. using multi-model median ensemble species distribution models. Ecol. Indic. 2021, 121, 107127. [Google Scholar] [CrossRef]
- Tebaldi, C.; Debeire, K.; Eyring, V.; Fischer, E.; Fyfe, J.; Friedlingstein, P.; Knutti, R.; Lowe, J.; O’Neill, B.; Sanderson, B. Climate model projections from the scenario model intercomparison project (ScenarioMIP) of CMIP6. Earth Syst. Dyn. 2021, 12, 253–293. [Google Scholar] [CrossRef]
- Fotheringham, A.S.; Oshan, T.M. Geographically weighted regression and multicollinearity: Dispelling the myth. J. Geogr. Syst. 2016, 18, 303–329. [Google Scholar] [CrossRef]
- Kuhn, M. Building Predictive Models in R Using the caret Package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef]
- Cao, Y.T.; Lu, Z.P.; Gao, X.Y.; Liu, M.L.; Sa, W.; Liang, J.; Wang, L.; Yin, W.; Shang, Q.H.; Li, Z.H. Maximum entropy modeling the distribution area of Morchella dill. Ex Pers. species in China under changing climate. Biology 2022, 11, 1027. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Naimi, B.; Araújo, M.B. sdm: A reproducible and extensible R platform for species distribution modelling. Ecography 2016, 39, 368–375. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Komac, B.; Esteban, P.; Trapero, L.; Caritg, R. Modelization of the current and future habitat suitability of Rhododendron ferrugineum using potential snow accumulation. PLoS ONE 2016, 11, e0147324. [Google Scholar] [CrossRef]
- Qin, M.X.; Jiao, L.N.; Yang, Z.Y.; Wang, J.X.; Ji, W. Potential distribution prediction and suitability evaluation Forsythia suspensa on MaxEnt and GIS in Shanxi Province. Chin. J. Agric. Resour. Reg. Plan. 2021, 42, 109–117. [Google Scholar]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef]
- Fang, J.; Lechowicz, M.J. Climatic limits for the present distribution of beech (Fagus L.) species in the world. J. Biogeogr. 2006, 33, 1804–1819. [Google Scholar] [CrossRef]
- Xu, J.; Deng, M.; Jiang, X.L.; Westwood, M.; Song, Y.G.; Turkington, R. Phylogeography of Quercus glauca (Fagaceae), a dominant tree of East Asian subtropical evergreen forests, based on three chloroplast DNA interspace sequences. Tree Genet. Genom. 2014, 11, 805. [Google Scholar] [CrossRef]
- Guan, X.Y.; Shi, W.; Cao, K.F. Effect of climate change in future on geographical distribution of widespread Quercus acutissima and analysis of dominant climatic factors. J. Trop. Subtrop. Bot. 2018, 26, 661–668. [Google Scholar]
- Cavender-Bares, J.; Holbrook, N.M. Hydraulic properties and freezing-induced cavitation in sympatric evergreen and deciduous oaks with contrasting habitats. Plant Cell Environ. 2001, 24, 1243–1256. [Google Scholar] [CrossRef]
- Gan, S.L. Development of Economic Animal and Plant Resources in Sichuan; Sichuan Social Sciences Press: Chengdu, China, 1988; pp. 99–180. [Google Scholar]
- Jump, A.S.; Mátyás, C.; Peñuelas, J. The altitude-for-latitude disparity in the range retractions of woody species. Trends Ecol. Evol. 2009, 24, 694–701. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Fang, Y.M. Prediction of potential suitable distribution areas of Quercus fabri in China based on an optimized Maxent model. Sci. Silvae Sin. 2018, 54, 153–164. [Google Scholar]
- Gao, W.Q.; Wang, X.F.; Jiang, Z.P.; Liu, J.F. Impact of climate change on the potential geographical distribution pattern and dominant climatic factors of Quercus variabilis. Acta Ecol. Sin. 2016, 36, 4475–4484. [Google Scholar]
- Jiang, X.L.; Xu, G.B.; Deng, M. Spatial genetic patterns and distribution dynamics of the rare oak Quercus chungii: Implications for biodiversity conservation in Southeast China. Forests 2019, 10, 821. [Google Scholar] [CrossRef]
- Tian, M.H.; Tang, A.J. Seed desiccation sensitivity of Quercus fabri and Castanopsis fissa (Fagaceae). Seed Sci. Technol. 2010, 38, 225–230. [Google Scholar] [CrossRef]
- Cheng, Y.P.; Hwang, S.Y.; Lin, T.P. Potential refugia in Taiwan revealed by the phylogeographical study of Castanopsis carlesii Hayata (Fagaceae). Mol. Ecol. 2005, 14, 2075–2085. [Google Scholar] [CrossRef]
- Xie, C.P.; Tian, E.; Jim, C.Y.; Liu, D.W.; Hu, Z.K. Effects of climate-change scenarios on the distribution patterns of Castanea henryi. Ecol. Evol. 2022, 12, e9597. [Google Scholar] [CrossRef]
- Liu, R.B.; Jiang, X.L.; Dai, X.L.; Deng, M. Potential distribution area of Quercus cocciferoides and its response to climate change. J. Plant Resour. Environ. 2019, 28, 85–95. [Google Scholar]
- Zhang, Q.C.; Gao, S.H.; Duan, Y.H.; Kong, L.K. China’s tropical and subtropical agroclimatic resources and their rational utilization. Resour. Sci. 1983, 01, 67–74. [Google Scholar]
- Chou, C.; Chiang, J.C.H.; Lan, C.W.; Chung, C.H.; Liao, Y.C.; Lee, C.J. Increase in the range between wet and dry season precipitation. Nat. Geosci. 2013, 6, 263–267. [Google Scholar] [CrossRef]
- Rajeevan, M.; Gadgil, S.; Bhate, J. Active and break spells of the Indian summer monsoon. J. Earth Syst. Sci. 2010, 119, 229–247. [Google Scholar] [CrossRef]
- Hughes, P.D.; Gibbard, P.L.; Ehlers, J. Timing of glaciation during the last glacial cycle: Evaluating the concept of a global ‘Last Glacial Maximum’ (LGM). Earth Sci. Rev. 2013, 125, 171–198. [Google Scholar] [CrossRef]
- Wang, S.Y.; Xu, X.T.; Shrestha, N.; Zimmermann, N.E.; Tang, Z.Y.; Wang, Z.H. Response of spatial vegetation distribution in China to climate changes since the Last Glacial Maximum (LGM). PLoS ONE 2017, 12, e0175742. [Google Scholar] [CrossRef]
- Scheiter, S.; Kumar, D.; Corlett, R.T.; Gaillard, C.; Langan, L.; Lapuz, R.S.; Martens, C.; Pfeiffer, M.; Tomlinson, K.W. Climate change promotes transitions to tall evergreen vegetation in tropical Asia. Glob. Chang. Biol. 2020, 26, 5106–5124. [Google Scholar] [CrossRef]
- Kim, D.; Ha, K.J.; Yeo, J.H. New drought projections over East Asia using evapotranspiration deficits from the CMIP6 warming scenarios. Earth’s Future 2021, 9, e2020EF001697. [Google Scholar] [CrossRef]
- Dai, A.; Zhao, T.; Chen, J. Climate change and drought: A precipitation and evaporation perspective. Curr. Clim. Change Rep. 2018, 4, 301–312. [Google Scholar] [CrossRef]
- Shukla, P.R.; Skeg, J.; Buendia, E.C.; Masson-Delmotte, V.; Pörtner, H.O.; Roberts, D.C.; Zhai, P.; Slade, R.; Connors, S.; Van Diemen, S. Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2019; pp. 99–100. [Google Scholar]
- Teskey, R.; Wertin, T.; Bauweraerts, I.; Ameye, M.; McGuire, M.A.; Steppe, K. Responses of tree species to heat waves and extreme heat events. Plant Cell Environ. 2015, 38, 1699–1712. [Google Scholar] [CrossRef]
- Lobell, D.B.; Hammer, G.L.; Chenu, K.; Zheng, B.; McLean, G.; Chapman, S.C. The shifting influence of drought and heat stress for crops in northeast Australia. Glob. Chang. Biol. 2015, 21, 4115–4127. [Google Scholar] [CrossRef]
- Lin, L.; Jiang, X.L.; Guo, K.Q.; Byrne, A.; Deng, M. Climate change impacts the distribution of Quercus section Cyclobalanopsis (Fagaceae), a keystone lineage in East Asian evergreen broadleaved forests. Plant Divers. 2023, in press. [Google Scholar] [CrossRef]
- Liu, G.H.; Fu, B.J. Effects of global climate change on forest ecosystems. J. Nat. Resour. 2001, 16, 71–78. [Google Scholar]
- Rathore, P.; Roy, A.; Karnatak, H. Modelling the vulnerability of Taxus wallichiana to climate change scenarios in South East Asia. Ecol. Indic. 2019, 102, 199–207. [Google Scholar] [CrossRef]
- Still, C.J.; Foster, P.N.; Schneider, S.H. Simulating the effects of climate change on tropical montane cloud forests. Nature 1999, 398, 608–610. [Google Scholar] [CrossRef]
- Foster, P. The potential negative impacts of global climate change on tropical montane cloud forests. Earth Sci. Rev. 2001, 55, 73–106. [Google Scholar] [CrossRef]
- Chen, I.C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Song, Y.G.; Petitpierre, B.; Deng, M.; Wu, J.P.; Kozlowski, G. Predicting climate change impacts on the threatened Quercus arbutifolia in montane cloud forests in southern China and Vietnam: Conservation implications. For. Ecol. Manag. 2019, 444, 269–279. [Google Scholar] [CrossRef]
- Hu, W.; Zhang, Z.Y.; Chen, L.D.; Peng, Y.S.; Wang, X. Changes in potential geographical distribution of Tsoongiodendron odorum since the Last Glacial Maximum. Chin. J. Plant Ecol. 2020, 44, 44. [Google Scholar] [CrossRef]
- Zhao, Y.C.; Zhao, M.Y.; Zhang, L.; Wang, C.Y.; Xu, Y.L. Predicting possible distribution of tea (Camellia sinensis L.) under climate change scenarios Using MaxEnt model in China. Agriculture 2021, 11, 1122. [Google Scholar] [CrossRef]
- Hong, T.D.; Ellis, R.H. A Protocol to Determine Seed Storage Behaviour; International Plant Genetic Resources Institute: Rome, Italy, 1996; p. 46. [Google Scholar]
- Li, J.J.; Jaganathan, G.K.; Han, K.; Liu, B.L. Embryo position of Castanopsis sclerophylla (Fagaceae) seeds with recalcitrant storage behavior differs from Quercus genus but response to desiccation shows no difference. Botany 2021, 100, 401–407. [Google Scholar] [CrossRef]
- Xia, K.; Hill, L.M.; Li, D.Z.; Walters, C. Factors affecting stress tolerance in recalcitrant embryonic axes from seeds of four Quercus (Fagaceae) species native to the USA or China. Ann. Bot. 2014, 114, 1747–1759. [Google Scholar] [CrossRef]
- Johnson, W.C.; Webb, T. The role of blue jays (Cyanocitta cristata L.) in the postglacial dispersal of Fagaceous trees in Eastern North America. J. Biogeogr. 1989, 16, 561–571. [Google Scholar] [CrossRef]
- Zheng, S.S.; Jiang, X.L.; Huang, Q.J.; Deng, M. Historical dynamics of semi-humid evergreen forests in the Southeast Himalaya biodiversity hotspot: A case study of the Quercus franchetii complex (Fagaceae). Front. Plant Sci. 2021, 12, 774232. [Google Scholar] [CrossRef]
- Jiang, X.L.; Deng, M.; Li, Y. Evolutionary history of subtropical evergreen broad-leaved forest in Yunnan Plateau and adjacent areas: An insight from Quercus schottkyana (Fagaceae). Tree Genet. Genom. 2016, 12, 104. [Google Scholar] [CrossRef]
- Jiang, X.L.; Su, Z.H.; Xu, G.B.; Deng, M. Genomic signals reveal past evolutionary dynamics of Quercus schottkyana and its response to future climate change. J. Syst. Evol. 2021, 59, 985–997. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Description | Relative Variable Importance (%) |
|---|---|---|
| PrecWQ | Precipitation of Warmest Quarter/mm | 11.2 |
| PrecDM | Precipitation of Driest Month/mm | 10.9 |
| MDR | Mean Diurnal Range (Mean of monthly (max temp—min temp))/°C | 8.1 |
| TS | Temperature Seasonality (Standard Deviation × 100)/% | 7.6 |
| MTWM | Max Temperature of Warmest Month/°C | 3.5 |
| PS | Precipitation Seasonality (Coefficient of Variation) | 3.4 |
| AMT | Annual Mean Temperature/°C | 3.0 |
| PrecWM | Precipitation of Wettest Month/mm | 2.5 |
| Climate Scenario | Unsuitability | Low Suitability | Medium Suitability | High Suitability | Suitable Area | |
|---|---|---|---|---|---|---|
| LGM | 860.39 | 395.91 | 31.84 | / | 427.76 | |
| Current | 817.03 | 108.27 | 98.02 | 109.6 | 315.89 | |
| 2041–2060 | SSP1-2.6 | 797.84 | 195.33 | 140.5 | / | 335.83 |
| SSP2-4.5 | 800.05 | 190.58 | 143.05 | / | 333.63 | |
| SSP3-7.0 | 796.17 | 209.82 | 127.68 | / | 337.51 | |
| SSP5-8.5 | 790.26 | 210.01 | 133.4 | / | 343.41 | |
| 2061–2080 | SSP1-2.6 | 795.95 | 194.74 | 142.98 | / | 337.72 |
| SSP2-4.5 | 787.44 | 206.53 | 139.7 | / | 346.23 | |
| SSP3-7.0 | 792.11 | 224.64 | 116.92 | / | 341.57 | |
| SSP5-8.5 | 776.65 | 230.36 | 126.66 | / | 357.02 | |
| 2081–2100 | SSP1-2.6 | 783.83 | 206.86 | 142.98 | / | 349.84 |
| SSP2-4.5 | 777.65 | 203.63 | 152.39 | / | 356.02 | |
| SSP3-7.0 | 763.11 | 234.07 | 136.49 | / | 370.56 | |
| SSP5-8.5 | 758.94 | 261.71 | 113.02 | / | 374.73 | |
| Climate Scenario | Range Expansion | No Occupancy | No Change | Range Contraction |
|---|---|---|---|---|
| LGM to current | 43.66 | 716.23 | 304.46 | 71.52 |
| Current to 2041–2060 SSP1-2.6 | 60.24 | 707.61 | 303.88 | 39.47 |
| Current to 2041–2060 SSP2-4.5 | 59.7 | 707.48 | 301.66 | 41.69 |
| Current to 2041–2060 SSP3-7.0 | 66.5 | 699.71 | 298.76 | 44.55 |
| Current to 2041–2060 SSP5-8.5 | 69.24 | 697.42 | 302.17 | 41.18 |
| 2041–2060 to 2061–2080 SSP1-2.6 | 10.67 | 805.41 | 357.88 | 8.94 |
| 2041–2060 to 2061–2080 SSP2-4.5 | 17.13 | 791.34 | 360.34 | 3.75 |
| 2041–2060 to 2061–2080 SSP3-7.0 | 12.05 | 792.84 | 360.25 | 7.74 |
| 2041–2060 to 2061–2080 SSP5-8.5 | 22.72 | 778.16 | 365.9 | 8.35 |
| 2061–2080 to 2081–2100 SSP1-2.6 | 15.39 | 799.33 | 366.5 | 2.05 |
| 2061–2080 to 2081–2100 SSP2-4.5 | 27.23 | 785.5 | 360.64 | 16.83 |
| 2061–2080 to 2081–2100 SSP3-7.0 | 42.04 | 777.82 | 361.04 | 11.26 |
| 2061–2080 to 2081–2100 SSP5-8.5 | 24.99 | 766.83 | 382.07 | 6.55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Lin, L.; Tan, Y.; Deng, M. How Climate Change Impacts the Distribution of Lithocarpus hancei (Fagaceae), a Dominant Tree in East Asian Montane Cloud Forests. Forests 2023, 14, 1049. https://doi.org/10.3390/f14051049
Yang Y, Lin L, Tan Y, Deng M. How Climate Change Impacts the Distribution of Lithocarpus hancei (Fagaceae), a Dominant Tree in East Asian Montane Cloud Forests. Forests. 2023; 14(5):1049. https://doi.org/10.3390/f14051049
Chicago/Turabian StyleYang, Yongjingwen, Lin Lin, Yunhong Tan, and Min Deng. 2023. "How Climate Change Impacts the Distribution of Lithocarpus hancei (Fagaceae), a Dominant Tree in East Asian Montane Cloud Forests" Forests 14, no. 5: 1049. https://doi.org/10.3390/f14051049
APA StyleYang, Y., Lin, L., Tan, Y., & Deng, M. (2023). How Climate Change Impacts the Distribution of Lithocarpus hancei (Fagaceae), a Dominant Tree in East Asian Montane Cloud Forests. Forests, 14(5), 1049. https://doi.org/10.3390/f14051049