
Morphological Systematics of Spathoglottis Blume (Orchidaceae: Collabieae) in Peninsular Malaysia and Borneo
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Taxon Sampling
2.2. Species Identification and Enumeration
2.3. Morphological Character and Ecological Characteristic Analyses
2.4. Character Matrix and Phylogenetic Analysis
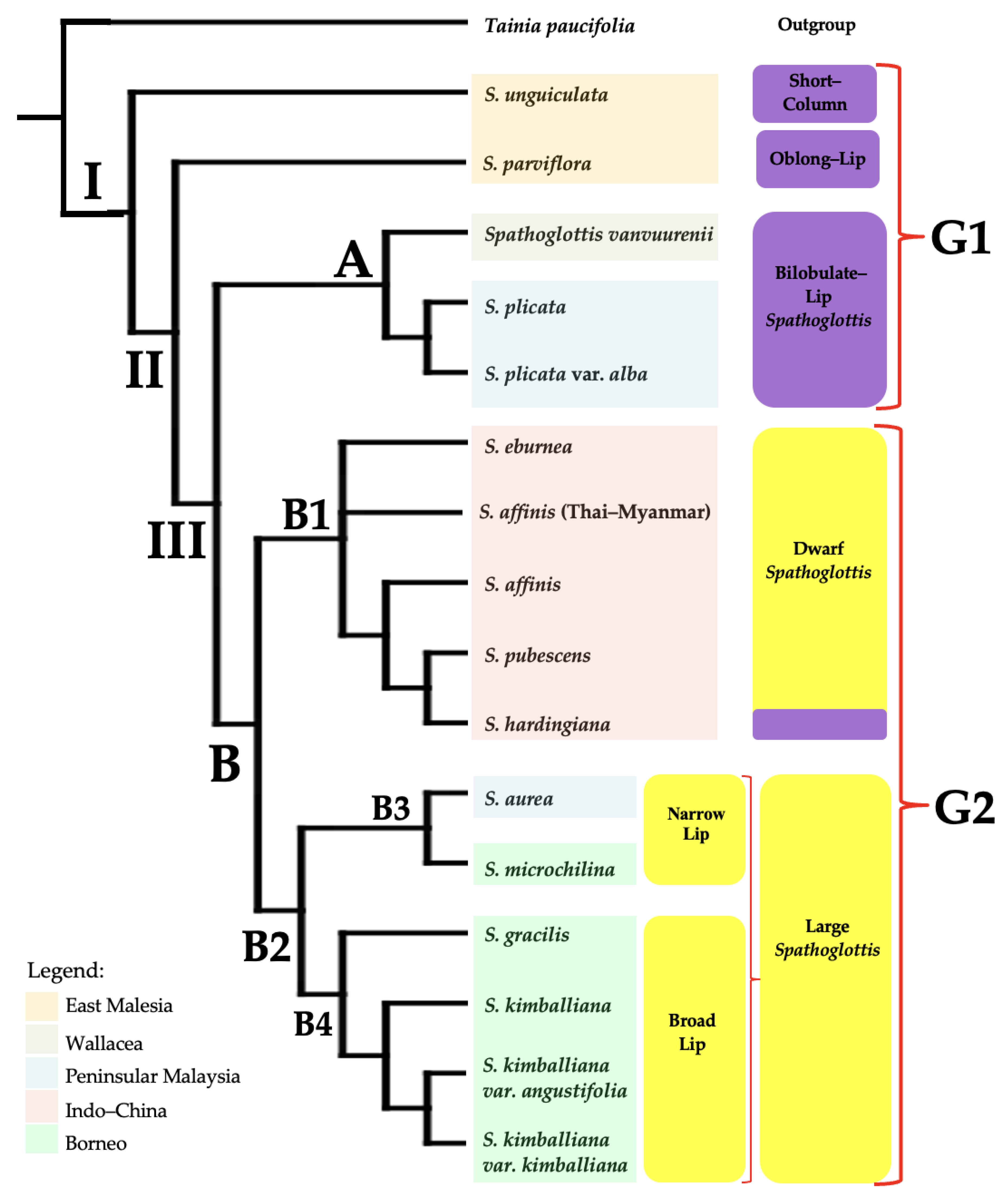
3. Results
3.1. Phylogeny Based on Morphological and Ecological Variation Analyses
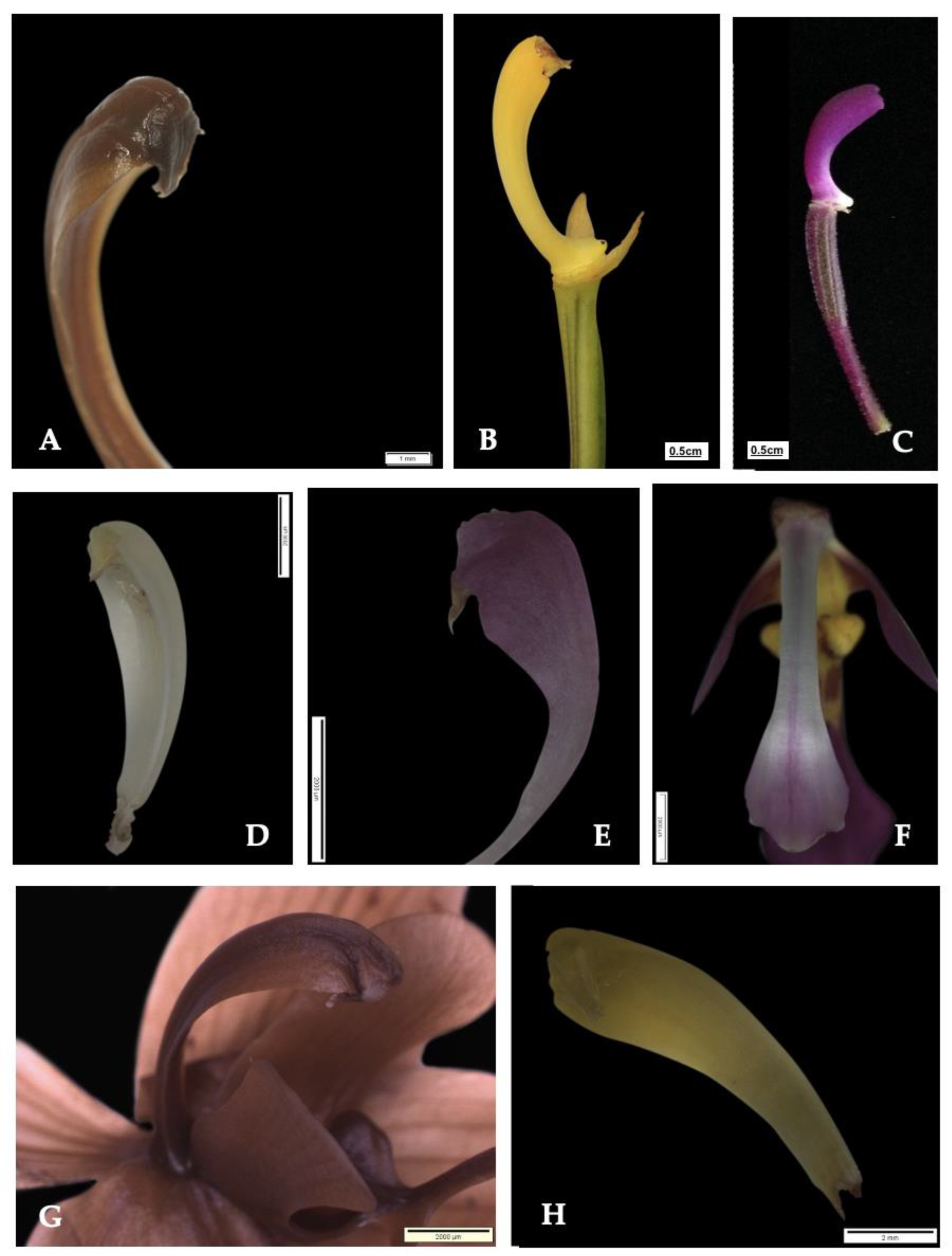
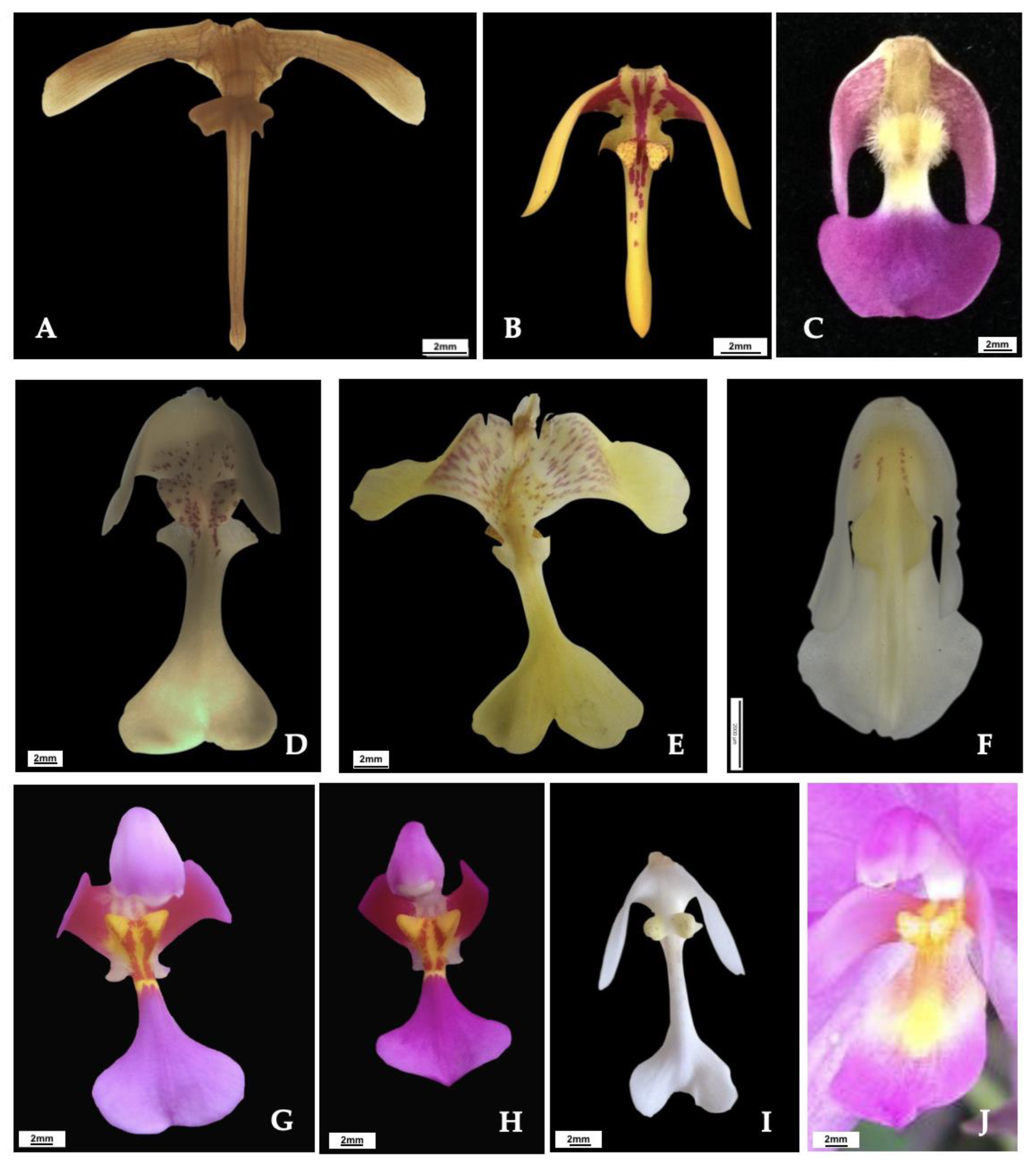
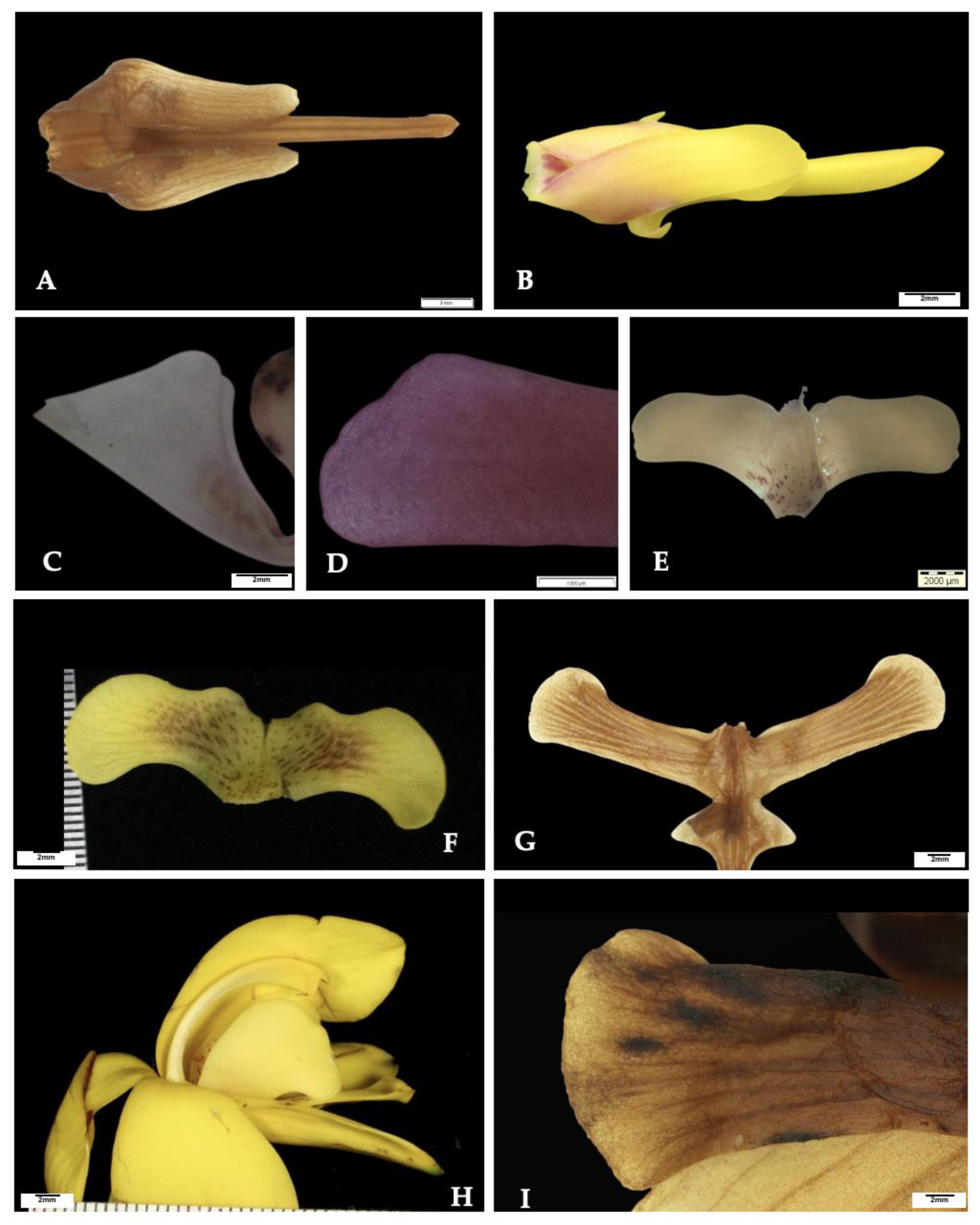
3.2. Delimiting Species Using Discreet Morphological Examination
4. Discussion
4.1. Morphological Variation and Phylogenetic Relationships between Species of Spathoglottis
4.2. Spathoglottis aurea and S. microchilina as Two Distinguished Species
4.3. Phylogenetic Relationships Based on Morphological Data and Molecular Data
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- POWO, Plants of the World Online (Facilitated by the Royal Botanic Gardens, Kew). Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:325895- (accessed on 20 January 2023).
- Ridley, H.N.; Hutchinson, J. The Flora of the Malay Peninsula Vol. 4; A. Asher & Co.: Utrecht, Holland; L. Reeve & Co.: London, UK, 1924. [Google Scholar]
- Holttum, R.E. A Revised Flora of Malaya: Orchids of Malaya Vol. 1, 3rd ed.; Government Printing Office: Singapore, 1964.
- Seidenfaden, G.; Wood, J.J. The Orchids of Peninsular Malaysia and Singapore: A Revision of R.E. Holttum: Orchids of Malaya; Olsen and Olsen in association with The Kew Royal Botanic Gardens, and Singapore Botanic Gardens: Fredensborg, Denmark, 1992. [Google Scholar]
- Chan, C.L.; Lamb, A.; Shim, P.S.; Wood, J.J. Orchids of Borneo Vol. 1 Introduction and a Selection of Species; The Sabah Society in Association with the Bentham—Moxon Trust, England: Kota Kinabalu, Malaysia, 1994. [Google Scholar]
- Wood, J.J. Orchids of Borneo Vol. 3: Dendrobium, Dendrochilum and Others; The Sabah Society in Association with the Bentham Moxon Trust, England: Kota Kinabalu, Malaysia, 1997. [Google Scholar]
- Wood, J.J. Orchids of Borneo Vol. 4; The Sabah Society in Association with the Bentham—Moxon Trust, England: Kota Kinabalu, Malaysia, 2003. [Google Scholar]
- Beaman, T.E.; Wood, J.J.; Beaman, R.S.; Beaman, J.H. Orchids of Sarawak; Natural History Publications in Association with the Royal Botanic Gardens, Kew: Kota Kinabalu, Malaysia, 2001. [Google Scholar]
- Comber, J.B. Orchids of Java; The Royal Botanic Garden’s Kew, Bentham Moxon Trust: London, UK, 1999. [Google Scholar]
- Comber, J.B. Orchids of Sumatra; Natural History Publications in Association with the Royal Botanic Gardens, Kew and Botanic Gardens, Singapore: Kota Kinabalu, Malaysia, 2001. [Google Scholar]
- Seidenfaden, G.; Smitinand, T. The Orchids of Thailand: A Preliminary List; The Siam Society: Bangkok, Thailand, 1959. [Google Scholar]
- Schuiteman, A.; de Vogel, E.F. Orchid Genera of Thailand, Laos, Cambodia and Vietnam; Nationaal Herbarium Nederland: Leiden, The Netherlands, 2000. [Google Scholar]
- Dockrill, A. Australian Indigenous Orchids Vol. 1. In Spathoglottis (Orchidaceae) in Australia and the Pacific Islands; Cribb, P.J., Tang, C.Z., Eds.; Springer: Berlin/Heidelberg, Germany, 1969; Volume 36, pp. 721–729. [Google Scholar]
- Cribb, P.J.; Tang, C.Z. Spathoglottis (Orchidaceae) in Australia and the Pacific Islands. Kew Bull. 1981, 36, 721–729. [Google Scholar] [CrossRef]
- Hallê, N. Flore de la Nouvelle Calêdonie et Dêpendances and Orchidacées. In Spathoglottis (Orchidaceae) in Australia and the Pacific Islands; Cribb, P.J., Tang, C.Z., Eds.; Springer: Berlin/Heidelberg, Germany, 1977; Volume 36, pp. 721–729. [Google Scholar]
- Ginibun, F.C.; Saad, M.R.M.; Hong, T.L.; Othman, R.Y.; Khalid, N.; Bhassu, S. Chloroplast DNA barcoding of Spathoglottis species for genetic conservation. Acta Holtic. 2010, 878, 453–459. [Google Scholar] [CrossRef]
- Nordin, F.A.; Saibeh, K.; Go, R.; Mangsor, K.N.A.; Othman, A.S. Molecular Phylogenetics of the Orchid Genus Spathoglottis (Orchidaceae: Collabieae) in Peninsular Malaysia and Borneo. Forests 2022, 13, 2079. [Google Scholar] [CrossRef]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. Available online: https://sweetgumnybg.org/science/ih (accessed on 14 March 2023).
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.70. Available online: http://mesquiteproject.org (accessed on 10 December 2022).
- Bateman, R.M.; Murphy, A.R.M.; Hollingsworth, P.M.; Hart, M.L.; Denholm, I.; Rudall, P.J. Molecular and Morphological Phylogenetics of the Digitate–Tubered Clade within Subtribe Orchidinae s.s. (Orchidaceae: Orchideae). Kew Bull. 2018, 73, 54. [Google Scholar] [CrossRef]
- Venhuis, C.; Venhuis, P.; Oostermeijer, J.G.B.; van Tienderen, P.H. Morphological Systematics of Serapias L. (Orchidaceae) in Southwest Europe. Plant Syst. Evol. 2007, 265, 165–177. [Google Scholar] [CrossRef]
- Devey, D.S.; Bateman, R.M.; Fay, M.F.; Hawkins, J.A. Friends or Relatives? Phylogenetics and Species Delimitation in the Controversial European Orchid Genus Ophrys. Ann. Bot. 2008, 101, 385–402. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Collection Number/Voucher Number | Collection Locality | Altitude (m) |
|---|---|---|---|
| FAN020/USMP12177 | Ton Nga Chang, Songkhla, Thailand | 100 |
| FAN023/USMP12178 | Fang District, Chiang Mai, Thailand | 1000 |
| FAN025/USMP12179 | Thailand–Myanmar Border | 1000 |
| FAN028/USMP12180 | Padang Tok Sheikh, G. Jerai, Kedah | 1131 |
| FAN006/USMP12181 | G. Ledang, Johor | 1200 |
| FAN008/USMP12182 | G. Lari Tembakau, Pahang | 1800 |
| FAN009/USMP12183 | G. Ulu Kali, Pahang | 1800 |
| FAN018/USMP12184 | G. Jerai, Kedah | 989 |
| FAN019/USMP12185 | Taman Rimba, G. Jerai, Kedah | 986 |
| FAN026/USMP12186 | Taman Rimba, G. Jerai, Kedah | 990 |
| FAN030/USMP12187 | Fraser’s Hill, Pahang | 1300 |
| FAN031/USMP12188 | Fraser’s Hill, Pahang | 1300 |
| FAN032/USMP12189 | Fraser’s Hill, Pahang | 1292 |
| FAN035/USMP12190 | Mile 49, Tanah Rata, Cameron Highlands, Pahang | 1400 |
| FAN036/USMP12191 | Mile 39, Tanah Rata, Cameron Highlands, Pahang | 1410 |
| FAN037/USMP12192 | G. Brinchang, Pahang | 1800 |
| FAN038/USMP12193 | G. Brinchang, Pahang | 1600 |
| FAN039/USMP12194 | G. Ulu Kali, Pahang | 1800 |
| FAN040/USMP12195 | G. Chin Chin, Pahang | 1800 |
| FAN041/USMP12196 | G. Brinchang, Pahang | 1800 |
| FAN044/USMP12197 | Padang Tok Sheikh, G. Jerai, Kedah | 1200 |
| FAN052/USMP12198 | G. Ulu Kali, Pahang | 1800 |
| FAN053/USMP12199 | G. Ulu Kali, Pahang | 1800 |
| FAN054/USMP12200 | Padang Tok Sheikh, G. Jerai, Kedah | 1200 |
| FAN057/USMP12201 | G. Bunga Buah, Selangor | 1400 |
| FAN058/USMP12202 | G. Bunga Buah, Selangor | 1400 |
| FAN059/USMP12203 | G. Bunga Buah, Selangor | 1400 |
| FAN060/USMP12204 | G. Bunga Buah, Selangor | 1400 |
| FAN069/USMP12205 | Tanah Rata, Cameron Highlands, Pahang | 1419 |
| FAN070/USMP12206 | Tanah Rata, Cameron Highlands, Pahang | 1419 |
| FAN071/USMP12207 | Huta Tinggi, Samarindo, Samosir, North Sumatra | 1400 |
| FAN098/USMP12208 | Tanah Rata, Cameron Highlands, Pahang | 1419 |
| FAN099/USMP12209 | Tanah Rata, Cameron Highlands, Pahang | 1419 |
| FAN022/USMP12210 | Fang District, Chiang Mai, Thailand | 1000 |
| FAN094/USMP12211 KIP1266f | Kg. Liposu, Ranau, Sabah | 850 |
| FAN016/USMP12212 | G. Baling, Kedah | 450 |
| FAN056/USMP12213 | G. Pong, Kenering, Perak | 420 |
| FAN105/USMP12214 K20160013 | Tg. Asan, Pulau Timun, Langkawi, Kedah | 200 |
| FAN085/USMP12215 | Ranau, Sabah | 1300 |
| FAN076/USMP12216 THH13 6 99 | Bidu Bidu FR, Telupid, Sabah | 71 |
| FAN077/USMP12217 | Bidu Bidu FR, Telupid, Sabah | 71 |
| FAN104/USMP12218 | Sungai Tongod, Telupid, Sabah | 60 |
| FAN067/USMP12219 | Mt. Kinabalu, Ranau, Sabah | 1300 |
| FAN092/USMP12220 | Kota Belud, Sabah | 800 |
| FAN093/USMP12221 | Pekan Nabalu, Ranau, Sabah | 1400 |
| FAN082/USMP12222 | Bundu Tuhan View Trail, Kinabalu Park, Sabah | 1601 |
| FAN083/USMP12223 | Kinabalu Park Research Centre, Sabah | 1599 |
| FAN084/USMP12224 | Kinabalu Park, Sabah | 850 |
| FAN086/USMP12225 | Mamut Copper Mine, Sabah | 1324 |
| FAN087/USMP12226 | Mamut Copper Mine, Sabah | 1485 |
| FAN088/USMP12227 | Mamut Copper Mine, Sabah | 1372 |
| FAN089/USMP12228 | Mamut Copper Mine, Sabah | 1405 |
| FAN091/USMP12229 | Sg. Lohan, Ranau, Sabah | 1375 |
| FAN095/USMP12230 | Tambunan, Sabah | 1000 |
| FAN096/USMP12231 | Ranau, Sabah | 850 |
| FAN061/USMP12232 | Wamena, Irian Jaya | 1679 |
| FAN001/USMP12233 | Peak of G. Ledang, Johor | 1000 |
| FAN002/USMP12234 | Peak of G. Ledang, Johor | 1100 |
| FAN003/USMP12235 | Penang Hill, Penang | 686 |
| FAN004/USMP12236 | Penang Hill, Penang | 735 |
| FAN005/USMP12237 | Sungai Rui, Kedah | 700 |
| FAN007/USMP12238 | K. Kubu Bharu, Selangor | 1392 |
| FAN010/USMP12239 | G. Ulu Kali, Pahang | 1800 |
| FAN011/USMP12240 | G. Jerai, Kedah | 980 |
| FAN012/USMP12241 | G. Jerai, Kedah | 1200 |
| FAN015/USMP12242 | Gerik, Perak | 1100 |
| FAN017/USMP12243 | Baling, Kedah | 650 |
| FAN021/USMP12244 | Peak of G. Jerai, Kedah | 1210 |
| FAN027/USMP12245 | G. Jerai, Kedah | 1000 |
| FAN029/USMP12246 | G. Jerai, Kedah | 1100 |
| FAN033/USMP12247 | Raub, Pahang | 1000 |
| FAN042/USMP12248 | Taiping Hill, Perak | 1200 |
| FAN043/USMP12249 | G. Jerai, Kedah | 926 |
| FAN046/USMP12250 | Taman Negara Endau Rompin, Johor | 78 |
| FAN047/USMP12251 | Taman Negara Endau–Rompin, Johor | 80 |
| FAN048/USMP12252 | Kota Tinggi Waterfall, Johor | 100 |
| FAN049/USMP12253 | Ladang Lok Heng, Kota Tinggi, Johor | 40 |
| FAN050/USMP12254 | Jalan Mersing, Kahang, Johor | 76 |
| FAN051/USMP12255 | Peak of G. Ledang, Johor | 1200 |
| FAN055/USMP12256 | G. Jerai, Kedah | 1200 |
| FAN063/USMP12257 | Lata Chemerong, Terengganu | 200 |
| FAN064/USMP12258 | Lata Chemerong, Terengganu | 70 |
| FAN065/USMP12259 | Lata Chemerong, Terengganu | 110 |
| FAN066/USMP12260 | Lata Chemerong, Terengganu | 70 |
| FAN072/USMP12261 | Tasik Kenyir, Terengganu | 30 |
| FAN073/USMP12262 | Mile 49, Tamparuli to Ranau Road, Sabah | 839 |
| FAN074/USMP12263 | Jalan Kaung to Ranau, Kota Belud, Sabah | 880 |
| FAN075/USMP12264 | Kg. Labong–Labong, Kota Belud, Sabah | 800 |
| FAN078/USMP12265 | Sg. Keripir, 45 miles to Tambunan, Sabah | 750 |
| FAN079/USMP12266 | Tambunan Road, Sabah | 757 |
| FAN080/USMP12267 | Tambunan Road, Sabah | 757 |
| FAN081/USMP12268 | Tambunan Road, Sabah | 468 |
| FAN090/USMP12269 | Mamut Copper Mine, Sabah | 1405 |
| FAN097/USMP12270 | Hulu Telom, Pahang | 708 |
| FAN100/USMP12271 | Semengoh Nature Reserve, Sarawak | 46 |
| FAN101/USMP12272 | Semengoh Nature Reserve, Sarawak | 50 |
| FAN102/USMP12273 | Bario, Sarawak | 1065 |
| FAN103/USMP12274 | Long Baleh, Sarawak | 944 |
| FAN034/USMP12275 | Lata Tembakah, Terengganu | 36 |
| FAN045/USMP12276 | Lata Tembakah, Terengganu | 36 |
| FAN068/USMP12277 | Fang District, Chiang Mai, Thailand | 800 |
| FAN024/USMP12278 | Isle of Pines, New Caledonia | 200 |
| FAN062/USMP12279 | Seram, Maluku Island | 834 |
| FAN597/USMP12280 | Taman Rimba Kenong, Pahang | 20 m |
| Character | Character States |
|---|---|
| Vegetative Characters | |
| 0 = dwarf (≤ 30 cm tall), 1 = large (≥ 30 cm up to 2 m tall) |
| 0 = thick, 1 = filiforme |
| 0 = ovoid, 1 = conical, 2 = flattened |
| 0 = green, 1 = purple, 2 = white |
| 0 = presence, 1 = absence |
| 0 = distinct, 1 = indistinct |
| 0 = presence, 1 = absence |
| 0 = greenish, 1 = tinged with purple |
| 0 = 1, 1 = 2–3, 2 = 4, 3 = > 4 |
| 0 = sheath clasping the stem, 1 = petiolated |
| 0 = plicate with strong midrib, 1 = non-plicate |
| 0 = thin, 1 = coriaceous (tough), 2 = velvety |
| 0 = glabrous, 1 = pubescent |
| 0 = entire, 1 = crisp |
| 0 = linear (grass-like), 1 = linear–lanceolate, 2 = lanceolate, 3 = broadly elliptic |
| 0 = acute, 1 = acuminate |
| 0 = greenish throughout, 1 = greenish-grey, 2 = green tinged with purple |
| 0 = grass-like (≤ 1.0 cm wide), 1 = narrow (c. 1.1–2.0 cm wide), 2 = medium (2.1–5.0 cm wide), 3 = very broad (>5.1 cm wide) |
| 0 = short (≤15.0 cm long), 1 = medium (15.1–50.0 cm long), 2 = long (50.1–100 cm long), 3 = very long (>100.1 cm long) |
| Reproductive Characters | |
| 0 = shorter than plant, 1 = longer than plant |
| 0 = green, 1 = purple |
| 0 = glabrous, 1 = covered with fine hairs |
| 0 = elliptic, 1 = obovate |
| 0 = deciduous, 1 = persistent |
| 0 = obovate, 1 = elliptic, 2 = triangular with wavy margin |
| 0 = free, 1 = clasping the peduncle |
| 0 = green, 1 = purple |
| 0 = hard, 1 = soft |
| 0 = glabrous, 1 = covered in fine hairs |
| 0 = upon flower senescence, 1 = flower still in bud stage |
| 0 = 2–3 (<5 flowers), 1 = many up to 20 flowers |
| 0 = determinate (terminal flower mature first), 1 = indeterminate (flower at the bottom mature first), 2 = all bloom at once |
| 0 = white, 1 = yellow, 2 = pink, 3 = mauve to deep purple |
| 0 = small (c. 3.0 cm in dimension), 1 = medium (c. 3.1–5.0 cm in dimension), 2 = large (>5.1 cm in dimension) |
| 0 = creamy yellow, 1 = pale yellow, 2 = golden yellow, 3 = other shades (not yellow) |
| 0 = presence (resupinate), 1 = absence (non resupinate) |
| 0 = insect pollinated, 1 = dimorphic cleistogamy, 2 = geitonogamy |
| 0 = 50% set into fruit, 1 = almost 100% set into fruit |
| 0 = spreading, 1 = drooping, 2 = turned backwards, 3 = remained closed |
| 0 = different in size, 1 = almost equal in width |
| 0 = ovate, 1 = elliptic, 2 = narrow–elliptic |
| 0 = thin, 1 = tough |
| 0 = flush red or brown on the outer part, 1 = no colouration, 2 = streaks with red in the inner part |
| 0 = hairy at base, 1 = hairy on the entire outer part, 2 = glabrous |
| 0 = obtuse, 1 = tapering towards end, 2 = thickened, 3 = mucronate |
| 0 = free, 1 = concave, 2 = curved backwards |
| 0 = presence, 1 = absence |
| 0 = obtuse, 1 = elliptic, 2 = oblong–elliptic |
| 0 = narrow, 1 = spathulate, 2 = obcordate, 3 = oblong–obovate, 4 = fiddle-shaped, 5 = square |
| 0 = length half of the sepals, 1 = length as long as sepals |
| 0 = incurved, 1 = entire, 2 = wavy |
| 0 = thread-like, 1 = almost rounded, 2 = square, 3 = bilobulate |
| 0 = mesochile narrow towards the end (≤4.0 mm wide), 1 = mesochile broad (c. 4.1–15 mm wide) |
| 0 = dilate, 1 = pointed, 2 = truncate |
| 0 = keels or raised ridges presence, 1 = keels or raised ridges absence |
| 0 = spotted red or purple, 1 = splashed with yellow/white at the base |
| 0 = triangular, 1 = square, 2 = oblong, 3 = falcate, 4 = auriculate |
| 0 = flushed with purple or crimson, 1 = splashed in yellow at base, 2 = spotted red or crimson or purple |
| 0 = sparsely hairy at base, 1 = glabrous |
| 0 = half as the width, 1 = almost double as wide |
| 0 = half as the length, 1 = almost similar to the length |
| 0 = presence, 1 = absence |
| 0 = on one plane, 1 = bending at an acute angle to the left |
| 0 = distinct, 1 = indistinct, 2 = absence |
| 0 = small appendages, 1 = triangular |
| 0 = oblong, 1 = cuneate, 2 = obcuneate, 3 = falcate, 4 = rounded |
| 0 = sparsely hairy at base, 1 = hairy entirely, 2 = hairy on top surface |
| 0 = spotted red or purple, 1 = yellow |
| 0 = united at base, 1 = risen individually |
| 0 = arched, 1 = slender, 2 = curved |
| 0 = shorter than the lip, 1 = as long as the lip |
| 0 = almost tubular, 1 = broadly winged |
| Ecological Characteristics | |
| 0 = terrestrial, 1 = lithophytic |
| 0 = montane forest above 900 m, 1 = lowland to hill forest (0–700 m asl), 2 = riverside |
| 0 = multisubstrates, 1 = limestone/quartzite, 2 = granite, 3 = ultramafic |
| Species | Characters and Character States | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | |
| S. affinis | 0 | 1 | 2 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| S. aurea | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 1 | 2 | 0 | 2 | 3 | 3 | 1 | 1 |
| S. eburnea | 0 | 1 | 2 | 2 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 2 | 1 | 1 | 1 |
| S. gracilis | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 2 | 0 | 0 | 3 | 1 | 0 | 0 |
| S. hardingiana | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 2 | 1 | 0 | 1 | 1 | 1 | 2 | 2 | 0 | 1 |
| S. kimballiana | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 1 | 2 | 0 | 0 | 3 | 3 | 1 | 0 |
| S. kimballiana var. angustifolia | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 0 |
| S. kimballiana var. kimballiana | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 0 |
| S. microchilina | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 2 | 0 | 0 | 3 | 3 | 1 | 0 |
| S. parviflora | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 3 | 0 | 0 | 1 | 0 | 1 | 2 | 0 | 0 | 3 | 3 | 1 | 0 |
| S. plicata | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 3 | 0 | 0 | 1 | 0 | 1 | 2 | 0 | 0 | 3 | 3 | 1 | 0 |
| S. plicata var. alba | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 3 | 0 | 0 | 1 | 0 | 1 | 2 | 0 | 0 | 3 | 3 | 1 | 0 |
| S. pubescens | 0 | 1 | 2 | 2 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| S. unguiculata | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 1 | 2 | 1 | 0 | 3 | 3 | 0 | 0 |
| S. vanvuurenii | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 3 | 0 | 0 | 1 | 0 | 1 | 2 | 0 | 0 | 3 | 3 | 1 | 0 |
| Tainia paucifolia | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 3 | 2 | 1 | 1 |
| 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | 40 | 41 | 42 | |
| S. affinis | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| S. aurea | 0 | 1 | 1 | 2 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 2 | 0 | 1 | 1 | 3 | 1 | 0 | 1 |
| S. eburnea | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| S. gracilis | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 2 | 2 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| S. hardingiana | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 3 | 0 | 3 | 1 | 0 | 0 | 2 | 0 | 1 | 0 |
| S. kimballiana | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 2 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | 40 | 41 | 42 | |
| S. kimballiana var. angustifolia | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 2 | 2 | 0 | 0 | 0 | 1 | 0 | 0 | 1 |
| S. kimballiana var. kimballiana | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| S. microchilina | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 3 | 0 | 0 | 1 |
| S. parviflora | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 3 | 2 | 3 | 0 | 2 | 0 | 0 | 0 | 0 | 0 |
| S. plicata | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 3 | 1 | 3 | 0 | 2 | 1 | 0 | 0 | 0 | 0 |
| S. plicata var. alba | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 1 | 1 | 0 |
| S. pubescens | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 3 | 0 | 0 | 0 | 0 | 1 | 1 | 0 |
| S. unguiculata | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 3 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| S. vanvuurenii | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 2 | 1 | 3 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Tainia paucifolia | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 2 | 3 | 1 | 3 | 0 | 0 | ? | 0 | 0 | 2 | 1 |
| 43 | 44 | 45 | 46 | 47 | 48 | 49 | 50 | 51 | 52 | 53 | 54 | 55 | 56 | 57 | 58 | 59 | 60 | 61 | 62 | 63 | |
| S. affinis | 2 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 3 | 1 | 0 | 1 | 0 | 2 | 2 | 0 | 0 | 0 | 0 | 0 |
| S. aurea | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 1 | 0 |
| S. eburnea | 0 | 1 | 0 | 0 | 1 | 0 | 5 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 2 | 0 | 0 | 0 | 1 | 0 |
| S. gracilis | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 3 | 1 | 0 | 0 | 0 | 1 | 2 | 0 | 1 | 0 | 0 | 1 |
| S. hardingiana | 0 | 2 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 2 | 1 | 0 | 0 | 1 | 0 |
| S. kimballiana | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 3 | 1 | 0 | 0 | 0 | 4 | 2 | 0 | 1 | 1 | 0 | 0 |
| S. kimballiana var. angustifolia | 1 | 0 | 0 | 0 | 0 | 0 | 4 | 1 | 1 | 1 | 1 | 2 | 0 | 0 | 4 | 2 | 0 | 1 | 1 | 1 | 0 |
| S. kimballiana var. kimballiana | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 3 | 1 | 0 | 0 | 0 | 2 | 2 | 0 | 0 | 0 | 0 | 0 |
| S. microchilina | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 1 | 0 |
| S. parviflora | 1 | 0 | 0 | 0 | 1 | 0 | 3 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 3 | 1 | 0 | 0 | 0 | 1 | 0 |
| S. plicata | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 3 | 1 | 0 | 0 | 0 | 1 | 2 | 0 | 0 | 0 | 0 | 0 |
| S. plicata var. alba | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 3 | 1 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| S. pubescens | 1 | 1 | 1 | 1 | 1 | 0 | 2 | 0 | 1 | 2 | 1 | 0 | 0 | 0 | 2 | 2 | 0 | 0 | 0 | 1 | 0 |
| S. unguiculata | 1 | 2 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 2 | 1 | 0 | 0 | 0 | 1 | 0 |
| S. vanvuurenii | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 3 | 1 | 0 | 0 | 0 | 2 | 2 | 0 | 0 | 0 | 1 | 0 |
| Tainia paucifolia | 1 | 2 | 3 | 2 | 0 | 2 | 2 | 1 | 2 | 1 | 1 | 2 | 0 | 0 | 2 | 0 | 1 | 0 | 0 | 1 | 0 |
| 64 | 65 | 66 | 67 | 68 | 69 | 70 | 71 | 72 | 73 | 74 | 75 | ||||||||||
| S. affinis | 0 | 1 | 0 | 2 | 0 | 1 | 2 | 1 | 1 | 1 | 0 | 1 | |||||||||
| S. aurea | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 2 | |||||||||
| S. eburnea | 0 | 1 | 3 | 0 | 1 | 1 | 2 | 1 | 1 | 1 | 0 | 1 | |||||||||
| S. gracilis | 0 | 1 | 0 | 0 | 0 | 1 | 2 | 1 | 1 | 0 | 0 | 3 | |||||||||
| S. hardingiana | 1 | 0 | 4 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | |||||||||
| S. kimballiana | 0 | 1 | 0 | 0 | 0 | 1 | 2 | 1 | 1 | 0 | 0 | 3 | |||||||||
| S. kimballiana var. angustifolia | 0 | 1 | 0 | 0 | 0 | 0 | 2 | 1 | 1 | 0 | 2 | 3 | |||||||||
| S. kimballiana var. kimballiana | 0 | 1 | 0 | 0 | 0 | 0 | 2 | 1 | 1 | 0 | 0 | 3 | |||||||||
| S. microchilina | 0 | 1 | 3 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 3 | |||||||||
| S. parviflora | 1 | 0 | 4 | 0 | 1 | 1 | 2 | 1 | 1 | 0 | 0 | 0 | |||||||||
| S. plicata | 0 | 1 | 2 | 2 | 0 | 0 | 2 | 1 | 1 | 0 | 1 | 0 | |||||||||
| S. plicata var. alba | 0 | 1 | 2 | 2 | 1 | 0 | 2 | 1 | 1 | 0 | 2 | 0 | |||||||||
| S. pubescens | 1 | 0 | 2 | 0 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | |||||||||
| S. unguiculata | 1 | 0 | 4 | 1 | 1 | 1 | 2 | 0 | 1 | 0 | 1 | 0 | |||||||||
| S. vanvuurenii | 0 | 1 | 0 | 0 | 0 | 1 | 2 | 1 | 1 | 0 | 0 | 0 | |||||||||
| Tainia paucifolia | 2 | ? | ? | ? | ? | ? | 0 | 1 | 1 | 0 | 2 | 0 | |||||||||
| Relationships from Morphological Data | Relationships from Molecular Data | |
|---|---|---|
| 1. | Species were first separated according to the colour of the flowers. | Groupings of Spathoglottis were first made according to plant size. |
| 2. | The Dwarf Spathoglottis is nested within the Yellow-Flowered Spathoglottis group. | The Dwarf Spathoglottis formed a separate group from the Large Spathoglottis group. |
| 3. | The Large Yellow-Flowered Spathoglottis group is well resolved, and species are grouped according to the shapes of labellum. | Species are arranged based on their geographical distribution, ecological niches and flower size. |
| 4. | Spathoglottis kimballiana is well placed within the Large Yellow-Flowered Spathoglottis group with other Spathoglottis species from Borneo. | Spathoglottis kimballiana is nested within the Large Purple Spathoglottis group. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nordin, F.A.; Raffi, A.; Go, R.; Seok Yien Yong, C.; Saibeh, K.; Othman, A.S. Morphological Systematics of Spathoglottis Blume (Orchidaceae: Collabieae) in Peninsular Malaysia and Borneo. Forests 2023, 14, 940. https://doi.org/10.3390/f14050940
Nordin FA, Raffi A, Go R, Seok Yien Yong C, Saibeh K, Othman AS. Morphological Systematics of Spathoglottis Blume (Orchidaceae: Collabieae) in Peninsular Malaysia and Borneo. Forests. 2023; 14(5):940. https://doi.org/10.3390/f14050940
Chicago/Turabian StyleNordin, Farah Alia, Akmal Raffi, Rusea Go, Christina Seok Yien Yong, Kartini Saibeh, and Ahmad Sofiman Othman. 2023. "Morphological Systematics of Spathoglottis Blume (Orchidaceae: Collabieae) in Peninsular Malaysia and Borneo" Forests 14, no. 5: 940. https://doi.org/10.3390/f14050940
APA StyleNordin, F. A., Raffi, A., Go, R., Seok Yien Yong, C., Saibeh, K., & Othman, A. S. (2023). Morphological Systematics of Spathoglottis Blume (Orchidaceae: Collabieae) in Peninsular Malaysia and Borneo. Forests, 14(5), 940. https://doi.org/10.3390/f14050940