
Proficient Lignocellulolytic Novel Bacterial Isolates from Diversified Galiyat Forests of Lower Himalaya

 ,
,  , , ,
, , ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Chemical Reagents and Culture Media
2.3. Physicochemical Characteristics of Samples
2.4. Bacterial Isolation, Screening, and Preservation
2.5. Characterization of Isolated Bacterial Strains
2.6. Phylogenetic Analysis of the Lignocellulose-Degrading Bacterial Isolates
2.7. Statistical Analyses
3. Results
3.1. Physical Characteristics
3.2. Enrichment of Lignocelluzlolytic Lower Himalayan Bacterial Strains
3.3. Enzymatic Characterization of Lower Himalayan Lignocellulolytic Bacterial Strains
3.4. Validation of Lignocellulolytic Enzyme Production by Lower Himalayan Bacterial Strains
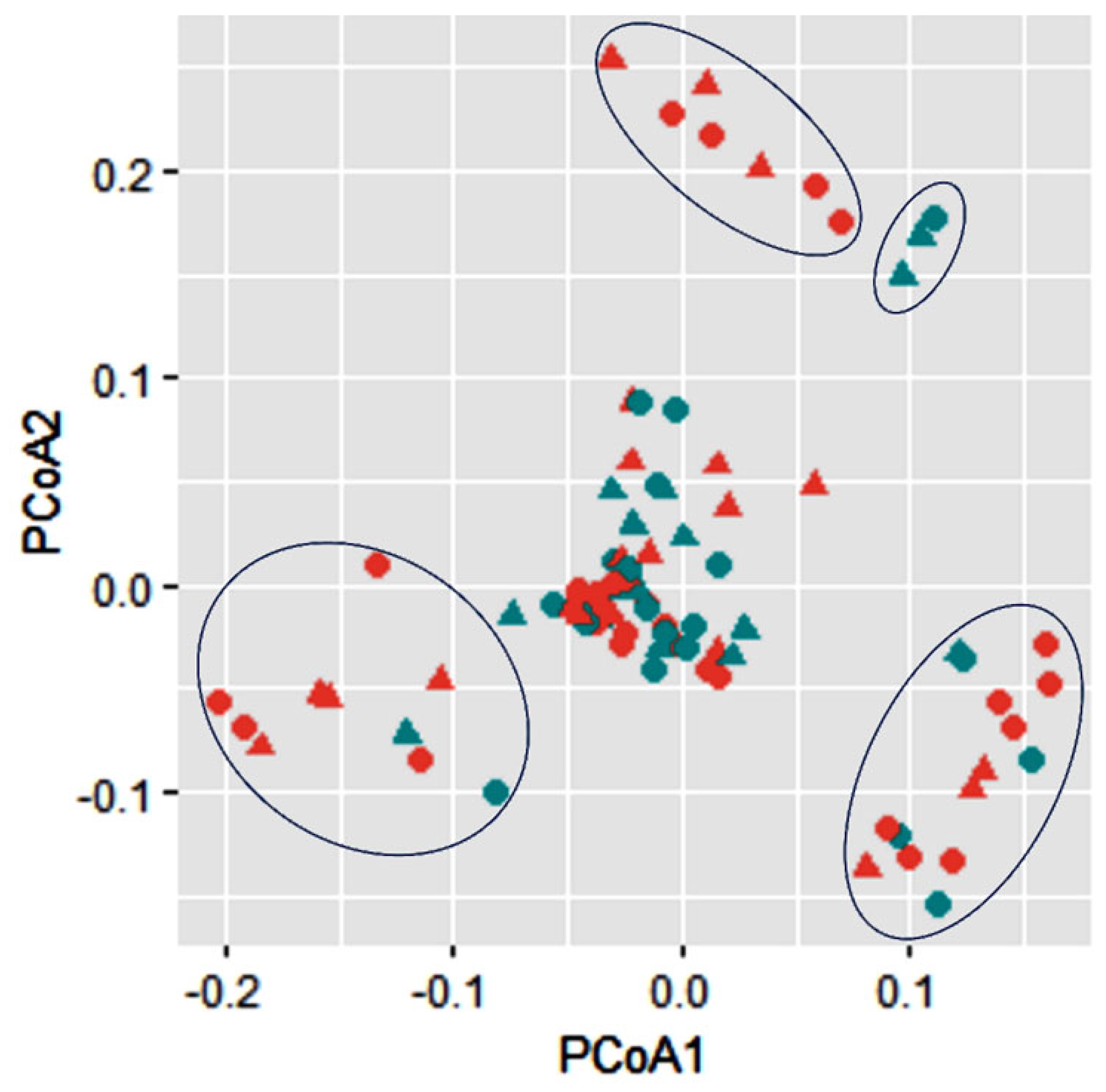
3.5. Phylogenetic Analysis of Lignocellulolytic Bacterial Strains
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bilgen, S. Structure and environmental impact of global energy consumption. In Renewable and Sustainable Energy Reviews; Elsevier Ltd.: Amsterdam, The Netherlands, 2014; Volume 38, pp. 890–902. [Google Scholar] [CrossRef]
- Ramos, M.D.N.; Milessi, T.S.; Candido, R.G.; Mendes, A.A.; André Aguiar, A. Enzymatic catalysis as a tool in biofuels production in Brazil: Current status and perspectives. Energy Sustain. Dev. 2022, 68, 103–119. [Google Scholar] [CrossRef]
- Lynd, L.R.; Weimer, P.J.; Van Zyl, W.H.; Pretorius, I.S. Microbial Cellulose Utilization: Fundamentals and Biotechnology. Bioresour. Technol. 2002, 66, 506–577. [Google Scholar]
- Oke, M.A.; Annuar, M.S.M.; Simarani, K. Mixed Lignocellulosic Biomass Degradation and Utilization for Bacterial Cellulase Production. Waste Biomass Valorization 2017, 8, 893–903. [Google Scholar] [CrossRef]
- Sindhu, R.; Binod, P.; Pandey, A. Biological pretreatment of lignocellulosic biomass—An overview. Bioresour. Technol. 2016, 199, 76–82. [Google Scholar] [CrossRef]
- Huntley, N.F.; Patience, J.F. Xylose: Absorption, fermentation, and post-absorptive metabolism in the pig. J. Anim. Sci. Biotechnol. 2018, 9, 4. [Google Scholar] [CrossRef] [Green Version]
- Jeong, H.J.; Cha, J.-Y.; Choi, J.H.; Jang, K.-S.; Lim, J.; Kim, W.-Y.; Seo, D.-C.; Jeon, J.-R. One-Pot Transformation of Technical Lignins into Humic-Like Plant Stimulants through Fenton-Based Advanced Oxidation: Accelerating Natural Fungus-Driven Humification. ACS Omega 2018, 3, 7441–7453. [Google Scholar] [CrossRef] [Green Version]
- Yu, O.; Kim, K.H. Lignin to materials: A focused review on recent novel lignin applications. Appl. Sci. 2020, 10, 4626. [Google Scholar] [CrossRef]
- Baruah, J.; Nath, B.K.; Sharma, R.; Kumar, S.; Deka, R.C.; Baruah, D.C.; Kalita, E. Recent trends in the pretreatment of lignocellulosic biomass for value-added products. Front. Energy Res. 2018, 6, 141. [Google Scholar] [CrossRef]
- Shen, P.; Murphy, D.V.; George, S.J.; Lapis-Gaza, H.; Xu, M.; Gleeson, D.B. Increasing the size of the microbial biomass altered bacterial community structure which enhances plant phosphorus uptake. PLoS ONE 2016, 11, e0166062. [Google Scholar] [CrossRef] [Green Version]
- Cragg, S.M.; Beckham, G.T.; Bruce, N.C.; Bugg, T.D.H.; Distel, D.L.; Dupree, P.; Etxabe, A.G.; Goodell, B.S.; Jellison, J.; McGeehan, J.E.; et al. Lignocellulose degradation mechanisms across the Tree of Life. Curr. Opin. Chem. Biol. 2015, 29, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Patel, R.J.; Bhaskaran, L. Screening of novel ascomycetes for the production of laccase enzyme using different lignin model compounds. Int. J. Pharma Bio Sci. 2016, 7, 452–458. [Google Scholar] [CrossRef]
- Wongfaed, N.; O-Thong, S.; Sittijunda, S.; Reungsang, A. Taxonomic and enzymatic basis of the cellulolytic microbial consor tium KKU- MC1 and its application in enhancing biomethane production. Sci. Rep. 2023, 13, 2968. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Mathur, A.S.; Gupta, R.P.; Barrow, C.J.; Tuli, D.K.; Puri, M. Enzyme systems of thermophilic anaerobic bacteria for lignocellulosic biomass conversion. Int. J. Biol. Macromol. 2021, 168, 572–590. [Google Scholar] [CrossRef] [PubMed]
- Bredon, M.; Dittmer, J.; Noël, C.; Moumen, B.; Bouchon, D. Lignocellulose degradation at the holobiont level: Teamwork in a keystone soil invertebrate 06 Biological Sciences 0605 Microbiology. Microbiome 2018, 6, 162. [Google Scholar] [CrossRef] [Green Version]
- Chukwuma, O.B.; Rafatullah, M.; Tajarudine, H.A.; Ismail, N. A review on bacterial contribution to lignocellulose breakdown into useful bio-products. Int. J. Environ. Res. Public Health 2021, 18, 6001. [Google Scholar] [CrossRef]
- Sethi, S.; Datta, A.; Gupta, B.L.; Gupta, S. Optimization of Cellulase Production from Bacteria Isolated from Soil. In International Scholarly Research Notices; Hindawi Publishing Corporation: London, UK, 2013; Volume 2013. [Google Scholar]
- Behera, B.C.; Singdevsachan, S.K.; Mishra, R.; Dutta, S.; Thatoi, H.N. Diversity, mechanism and biotechnology of phosphate solubilising microorganism in mangrove—A review. Biocatal. Agric. Biotechnol. 2014, 3, 97–110. [Google Scholar] [CrossRef]
- Curran, L.L.K.; Pham, L.T.M.; Sale, K.L.; Simmons, B.A. Review of advances in the development of laccases for the valorization of lignin to enable the production of lignocellulosic biofuels and bioproducts. Biotechnol. Adv. 2022, 54, 107809. [Google Scholar] [CrossRef]
- Arora, R.; Behera, S.; Sharma, N.K.; Kumar, S. Bioprospecting thermostable cellulosomes for efficient biofuel production from lignocellulosic biomass. Bioresour. Bioprocess. 2015, 2, 38. [Google Scholar] [CrossRef] [Green Version]
- Breisha, G.Z. Production of 16% ethanol from 35% sucrose. Biomass Bioenergy 2010, 34, 1243–1249. [Google Scholar] [CrossRef]
- Jaggi, V.; Brindhaa, N.T.; Sahgal, M. Microbial diversity in north western Himalayan agroecosystems: Functions and applications. Microbiol. Adv. High. Alt. Agro-Ecosyst. Sustain. 2020, 2020, 135–161. [Google Scholar]
- Thakur, V.; Kumar, V.; Kumar, S.; Singh, D. Diverse culturable bacterial communities with cellulolytic potential revealed from pristine habitat in Indian trans-Himalaya. Can. J. Microbiol. 2018, 64, 798–808. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, S.; Gowdaman, V.; Prabagaran, S.R. Culturable and Culture-Independent Bacterial Diversity and the Prevalence of Cold-Adapted Enzymes from the Himalayan Mountain Ranges of India and Nepal. Microb. Ecol. 2014, 69, 472–491. [Google Scholar] [CrossRef] [PubMed]
- Aziz, K.; Saleem, A.; Malik, A.M. Litter Fall Production and Decomposition in Deodar Forest Ecosystem. Pak. J. Agric. Res. 2019, 32, 441. [Google Scholar] [CrossRef]
- Hayat, W.; Aman, H.; Irshad, U.; Azeem, M.; Iqbal, A.; Nazir, R. Analysis of ecological attributes of bacterial phosphorus solubilizers, native to pine forests of Lower Himalaya. Appl. Soil Ecol. 2017, 112, 51–59. [Google Scholar] [CrossRef]
- Rahim, I.; Ali, S.M.; Aslam, M. GIS Based Landslide Susceptibility Mapping with Application of Analytical Hierarchy Process in District Ghizer, Gilgit Baltistan Pakistan. J. Geosci. Environ. Prot. 2018, 06, 34–49. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.K.; Park, T.S.; Kwon, I.H.; Piao, M.Y.; Lee, C.H.; Ha, J.K. Characterization of cellulolytic and xylanolytic enzymes of Bacillus licheniformis JK7 isolated from the rumen of a native Korean goat. Asian-Australas. J. Anim. Sci. 2013, 26, 50–58. [Google Scholar] [CrossRef]
- Malini, B.; Revathi, M.; Yadav, A.; Sakthivel, N. Purification and Characterization of a Thermophilic Cellulase from a Novel Cellulolytic Strain, Paenibacillus barcinonensis. J. Microbiol. Biotechnol. 2012, 22, 1501–1509. [Google Scholar]
- Khatiwada, P.; Ahmed, J.; Sohag, M.H.; Islam, K.; Azad, A.K. Isolation, Screening and Characterization of Cellulase Producing Bacterial Isolates from Municipal Solid Wastes and Rice Straw Wastes. J. Bioprocess. Biotech. 2016, 6, 4–8. [Google Scholar]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Lee, S.; Garcia, S.; Qian, X. Modeling and Simulation of the Mixed Mode Ventilation Strategies with Heat Recovery and Energy Recovery Wheels for Energy Conservation and IAQ Improvement in the Commercial Buildings. In Recent Researches in Urban Sustainability. Architecture and Structures; Morgan State University: Baltimore, MD, USA, 2013; pp. 209–215. [Google Scholar]
- Kamble, R.D.; Jadhav, A.R. Isolation, purification, and characterization of xylanase produced by a new species of bacillus in solid state fermentation. Int. J. Microbiol. 2012, 2012, 683193. [Google Scholar] [CrossRef] [Green Version]
- Bagewadi, Z.K.; Mulla, S.I.; Ninnekar, H.Z. Optimization of laccase production and its application in delignification of biomass. Int. J. Recycl. Org. Waste Agric. 2017, 6, 351–365. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Kong, F. Isolation and identification of potassium-solubilizing bacteria from tobacco rhizospheric soil and their effect on tobacco plants. Appl. Soil Ecol. 2014, 82, 18–25. [Google Scholar] [CrossRef]
- Woo, H.L.; Hazen, T.C.; Simmons, B.A.; DeAngelis, K.M. Enzyme activities of aerobic lignocellulolytic bacteria isolated from wet tropical forest soils. Syst. Appl. Microbiol. 2014, 37, 60–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hess, J.; Balasundaram, S.V.; Bakkemo, R.I.; Drula, E.; Henrissat, B.; Högberg, N.; Skrede, I. Niche differentiation and evolution of the wood decay machinery in the invasive fungus Serpula lacrymans. ISME J. 2021, 15, 592–604. [Google Scholar] [CrossRef] [PubMed]
- Nuccio, E.E.; Starr, E.; Karaoz, U.; Brodie, E.L.; Zhou, J.; Tringe, S.G.; Malmstrom, R.R.; Woyke, T.; Banfield, J.F.; Firestone, M.K.; et al. Niche differentiation is spatially and temporally regulated in the rhizosphere. ISME J. 2020, 14, 999–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez, D.J.; Dini-Andreote, F.; DeAngelis, K.M.; Singer, S.W.; Falcão Salles, J.; van Elsas, J.D. Ecological Insights into the Dynamics of Plant Biomass Degrading Microbial Consortia. Trends Microbiol. 2017, 25, 788–796. [Google Scholar] [CrossRef]
- Menzel, T.; Neubauer, P.; Junne, S. Role of microbial hydrolysis in anaerobic digestion. Energies 2020, 13, 5555. [Google Scholar] [CrossRef]
- Serrano-Gamboa, J.G.; Rojas-Herrera, R.A.; González-Burgos, A.; Folch-Mallol, J.L.; Jiménez, D.J.; Sánchez-González, M.N. Degradation profile of nixtamalized maize pericarp by the action of the microbial consortium PM-06. AMB Express 2019, 9, 85. [Google Scholar] [CrossRef]
- Berini, F.; Casciello, C.; Marcone, G.L.; Marinelli, F. Metagenomics: Novel enzyme from non- culturable microbes. FEMS Microbiol. Lett 2017, 364, 21. [Google Scholar] [CrossRef]
- Zhang, X.; Borjigin, Q.; Gao, J.L.; Yu, X.F.; Hu, S.P.; Zhang, B.Z.; Sheng-Cai Han, S.C. Community succession and functional prediction of microbial consortium with straw degradation during subculture at low temperature. Sci. Rep. 2022, 12, 20163. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, L.; Xian, H.; Zhang, X. Newly isolated cellulose-degrading bacterium achromobacter xylosoxidans L2 has deinking potential. BioResources 2019, 14, 2256–2268. [Google Scholar] [CrossRef]
- Hu, X.; Yu, J.; Wang, C.; Chen, H. Cellulolytic Bacteria Associated with the Gut of Dendroctonus armandi Larvae (Coleoptera: Curculionidae: Scolytinae). Forests 2014, 5, 455–465. [Google Scholar] [CrossRef]
- Álvarez, C.; Reyes-Sosa, F.M.; Díez, B. Enzymatic hydrolysis of biomass from wood. Microb. Biotechnol. 2016, 9, 149–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghasemi, Y.; Sadeghi, H.; Gholami, A.; Mohkam, M.; Kargar, M. Isolation and Identification of Highly Xylanase Producing Bacterium Sphingobacterium sp. SaH-05 from Soil. Int. J. Sci. Eng. Res. 2014, 5, 205–209. [Google Scholar]
- Anand, A.A.P.; Vennison, S.J.; Sankar, S.G.; Gilwax, D.I.; Vasan, P.T.; Raghuraman, T.; Geoffrey, C.J.; Vendan, S.E. Isolation and characterization of bacteria from the gut of Bombyx mori that degrade cellulose, xylan, pectin and starch and their impact on digestion. J. Insect Sci. 2010, 10, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berini, F.; Verce, M.; Ausec, L.; Rosini, E.; Tonin, F.; Pollegioni, L.; Mandić-Mulec, I. Isolation and characterization of a heterologously expressed bacterial laccase from the anaerobe Geobacter metallireducens. Appl. Microbiol. Biotechnol. 2018, 102, 2425–2439. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues Reis, C.E.; Hu, B. Vinasse from sugarcane ethanol production: Better treatment or better utilization? Front. Energy Res. 2017, 5, 7. [Google Scholar] [CrossRef] [Green Version]
- Bomble, Y.J.; Lin, C.Y.; Amore, A.; Wei, H.; Holwerda, E.K.; Ciesielski, P.N.; Donohoe, B.S.; Decker, S.R.; Lynd, L.R.; Himmel, M.E. Lignocellulose deconstruction in the biosphere. Curr. Opin. Chem. Biol. 2017, 41, 61–70. [Google Scholar] [CrossRef]
- Tolalpa, L.; Salles, J.F.; van Elsas, J.D. Bacterial Synergism in Lignocellulose Biomass Degradation—Complementary Roles of Degraders As Influenced by Complexity of the Carbon Source. Front. Microbiol. 2017, 8, 1628. [Google Scholar] [CrossRef]
- Puentes-Téllez, P.E.; Falcao Salles, J. Construction of Effective Minimal Active Microbial Consortia for Lignocellulose Degradation. Microb. Ecol. 2018, 76, 419–429. [Google Scholar] [CrossRef] [Green Version]
- Azeem, M.; Borg-Karlson, A.K.; Rajarao, G.K. Sustainable bio-production of styrene from forest waste. Bioresour. Technol. 2013, 144, 684–688. [Google Scholar] [CrossRef] [PubMed]
- Ferdes, M.; Dincă, M.N.; Moiceanu, G.; Zăbavă, B.S.; Paraschiv, G. Microorganisms and Enzymes Used in the Biological Pretreatment of the Substrate to Enhance Biogas Production: A Review. Sustainability 2020, 12, 7205. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Origin | Site | Geographical Coordinates | Elevation (Meters) | Type of Sample | Code | Soil Type | pH | Organic Matter (%) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Thandiani | Site 1 | 34° 13 51.43 N 73° 21 08.42 E | 2664 | Soil | TS1 | Clayey | 6.7 | 2.23 ± 0.17 |
| 2 | Wood | TW1 | N/A | N/A | N/A | ||||
| 3 | Site2 | 34° 13 51.47 N 73° 21 08.42 E | 2665 | Soil | TS2 | Clayey Loam | 6.9 | 2.17 ± 0.24 | |
| 4 | Wood | TW2 | N/A | N/A | N/A | ||||
| 5 | Nathiagali | Site 1 | 34° 03 51.22 N 73° 24 22.44 E | 2540 | Soil | NS1 | Clayey | 7.0 | 2.91 ± 0.2 |
| 6 | Wood | NW1 | N/A | N/A | N/A | ||||
| 7 | Site2 | 34° 03 51.22 N 73° 24 22.44 E | 2551 | Soil | NS2 | Clayey | 6.8 | 2.79 ± 0.13 | |
| 8 | Wood | NW2 | N/A | N/A | N/A |
| No. | Strain Code | Enzymatic Potential | Closest Relative | % Identity | Accession ID |
|---|---|---|---|---|---|
| 1 | TS1B3 | Cellulase | Ochrobactrum sp. strain DB30 | 100 | MH588247.1 |
| 2 | TW1B4 | Cellulase | Pseudomonas mendocina strain fsznc-01 | 99.36 | MK106366.1 |
| 3 | TS1B9 | Cellulase | Pseudomonas sp. strain Gamma-14 | 99.54 | MH703445.1 |
| 4 | NW1B7 | Cellulase | Pseudomonas sp. strain 2016NX1 | 98.94 | MH368491.1 |
| 5 | NW2B2 | Cellulase | Pseudomonas sp. strain JLT15231 | 94.95 | KX989279.1 |
| 6 | NS2B10 | Cellulase | Pseudomonas aeruginosa strain PA06 | 99.38 | MN326719.1 |
| 7 | NS1B8 | Cellulase | Ochrobactrum sp. strain YE2-10 | 97.01 | KT992334.1 |
| 8 | NS1B10 | Cellulase | Pseudomonas aeruginosa isolate H8 | 99.81 | MG706125.1 |
| 9 | NW1B8 | Cellulase | Pseudomonas sp. LMB-2 | 99.64 | KR048282.1 |
| 10 | NW1B11 | Cellulase | Pseudomonas stutzeri strain | 98.05 | KT380573.1 |
| 11 | NS2B9 | Cellulase | Pseudomonas aeruginosa M4 | 99.9 | MT180543.1 |
| 12 | TS1B5 | Cellulase | Verticiella sediminum strain XJ-4 | 95.28 | KU525641.1 |
| 13 | NW2B7 | Cellulase | Brevundimonas sp. strain HBUMF35 | 98.33 | MF285791.1 |
| 14 | NS1B7 | Cellulase | Pseudomonas aeruginosa YB01 | 98.7 | CP028132.1 |
| 15 | NS1B4 | Cellulase | Pseudomonas aeruginosa strain RHH1 | 99.73 | HQ143612.1 |
| 16 | NS2B1 | Cellulase | Pseudomonas aeruginosa strain QK-4 | 98.71 | MH746107.1 |
| 17 | TW1B9 | Laccase | Brevundimonas vesicularis SCSIO_43705 | 100 | MH283789.1 |
| 18 | NS1B6 | Laccase | Achromobacter sp. P3′(2012) | 99.21 | JX083295.1 |
| 19 | TS2B12 | Laccase | Flavitalea sp. HBUM179779 | 89.87 | KR906086.1 |
| 20 | TS1B11 | Laccase | Hydrogenophaga sp. PBC | 95.84 | CP017311.1 |
| 21 | NS1B12 | Laccase | Stenotrophomonas maltophilia strain C1A | 99.2 | KJ466103.1 |
| 22 | TS2B7 | Laccase | Rhizobiales bacterium ROi51 | 98.21 | EF219048.1 |
| 23 | NS2B8 | Laccase | Stenotrophomonas acidaminiphila PRSB5 | 99.63 | MG450366.1 |
| 24 | TS1B12 | Laccase | Serinibacter salmoneus strain Kis4-28 | 96.63 | NR_041692.1 |
| 25 | NS2B7 | Laccase | Paracoccus sp. PW-P2 | 99.36 | JF772176.1 |
| 26 | NW2B5 | Laccase | Pseudomonas sp. strain BNC2906 | 99.9 | MN136109.1 |
| 27 | NS1B1 | Xylanase | Xenophilus sp. C12b | 91.83 | GU394956.1 |
| 28 | NS1B2 | Xylanase | Salmonella enterica CVM N17S1509 | 99.3 | CP052817.1 |
| 29 | TS2B15 | Xylanase | Bacillus sp. EnB-alf12 | 97.62 | KP792637.1 |
| 30 | NS2B3 | Xylanase | Bacillus cereus strain RTR | 99.46 | MK014289.1 |
| 31 | NW2B4 | Xylanase | Sphingopyxis panaciterrulae strain DCY34 | 99.54 | NR_116164.1 |
| 32 | TS2B1 | Xylanase | Stenotrophomonas acidaminiphila strain PRSB5 | 99.63 | MG450366.1 |
| 33 | TS1B10 | Xylanase | Pseudoxanthomonas sp. 45-47b | 99.03 | LC481375.1 |
| 34 | NS2B5 | Xylanase | Pseudoxanthomonas sp. ROi44 | 99.34 | EF219047.1 |
| 35 | TW2B4 | Xylanase | Klebsiella pneumoniae strain FNK1 (nv) | 93.07 | AF492000.1 |
| 36 | TW2B7 | Xylanase | Stenotrophomonas acidaminiphila strain JR34 | 99.44 | MG450368 |
| No. | Strains | Activity | Substrate | Enzymatic Activity (UI/ml) |
|---|---|---|---|---|
| 1 | Branhamella sp. | Cellulase | CMC | 2.56 |
| 2 | P. terrae ME27-1 | Cellulase | CMC | 2.08 |
| 3 | Bacillus licheniformis JK7 | Cellulase | CMC | 1.46 |
| 4 | Ganoderma applanatum LPB MR-56 | Cellulase | CMC | 3.29 |
| 5 | NS1B7 | Cellulase | CMC | 4.99 |
| 6 | Ganoderma applanatum LPB MR-56 | Xylanase | 2.09 | |
| 7 | Paenibacillus sp. XJ18 | Xylanase | Corn cob xylan | 2.4578 |
| 8 | Pseudomonas stutzeri | Xylanase | Molasses | 1.3 |
| 9 | Cellulosimicrobium cellulans | Xylanase | Baggasse | 0.7 |
| 10 | TS2B1 | Xylanase | Corn cob xylan | 6.6 |
| 11 | P. sanguineus | Laccase | ABTS | 2 |
| 12 | Lysinibacillus sphaericus JD1103 | Laccase | ABTS | 1.13 |
| 13 | L. sphaericus BR2308 | Laccase | ABTS | 1.93 |
| 14 | Flavitalea sp. TS2B12 | Laccase | ABTS | 4.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Awan, M.O.U.; Iqbal, A.; Rashid, M.I.; Irshad, U.; Hafeez, F.; Ullah, F.; Irshad, M.; Ondrasek, G.; Mustac, I.; Nazir, R. Proficient Lignocellulolytic Novel Bacterial Isolates from Diversified Galiyat Forests of Lower Himalaya. Forests 2023, 14, 1180. https://doi.org/10.3390/f14061180
Awan MOU, Iqbal A, Rashid MI, Irshad U, Hafeez F, Ullah F, Irshad M, Ondrasek G, Mustac I, Nazir R. Proficient Lignocellulolytic Novel Bacterial Isolates from Diversified Galiyat Forests of Lower Himalaya. Forests. 2023; 14(6):1180. https://doi.org/10.3390/f14061180
Chicago/Turabian StyleAwan, Malik Owais Ullah, Akhtar Iqbal, Muhammad Imtiaz Rashid, Usman Irshad, Farhan Hafeez, Farid Ullah, Muhammad Irshad, Gabrijel Ondrasek, Ivan Mustac, and Rashid Nazir. 2023. "Proficient Lignocellulolytic Novel Bacterial Isolates from Diversified Galiyat Forests of Lower Himalaya" Forests 14, no. 6: 1180. https://doi.org/10.3390/f14061180
APA StyleAwan, M. O. U., Iqbal, A., Rashid, M. I., Irshad, U., Hafeez, F., Ullah, F., Irshad, M., Ondrasek, G., Mustac, I., & Nazir, R. (2023). Proficient Lignocellulolytic Novel Bacterial Isolates from Diversified Galiyat Forests of Lower Himalaya. Forests, 14(6), 1180. https://doi.org/10.3390/f14061180