



Hydrological Coupling and Decoupling of Hydric Hemiboreal Forest Sites Inferred from Soil Water Models and Tree-Ring Chronology



Abstract
:1. Introduction
2. Materials and Methods
2.1. Climate
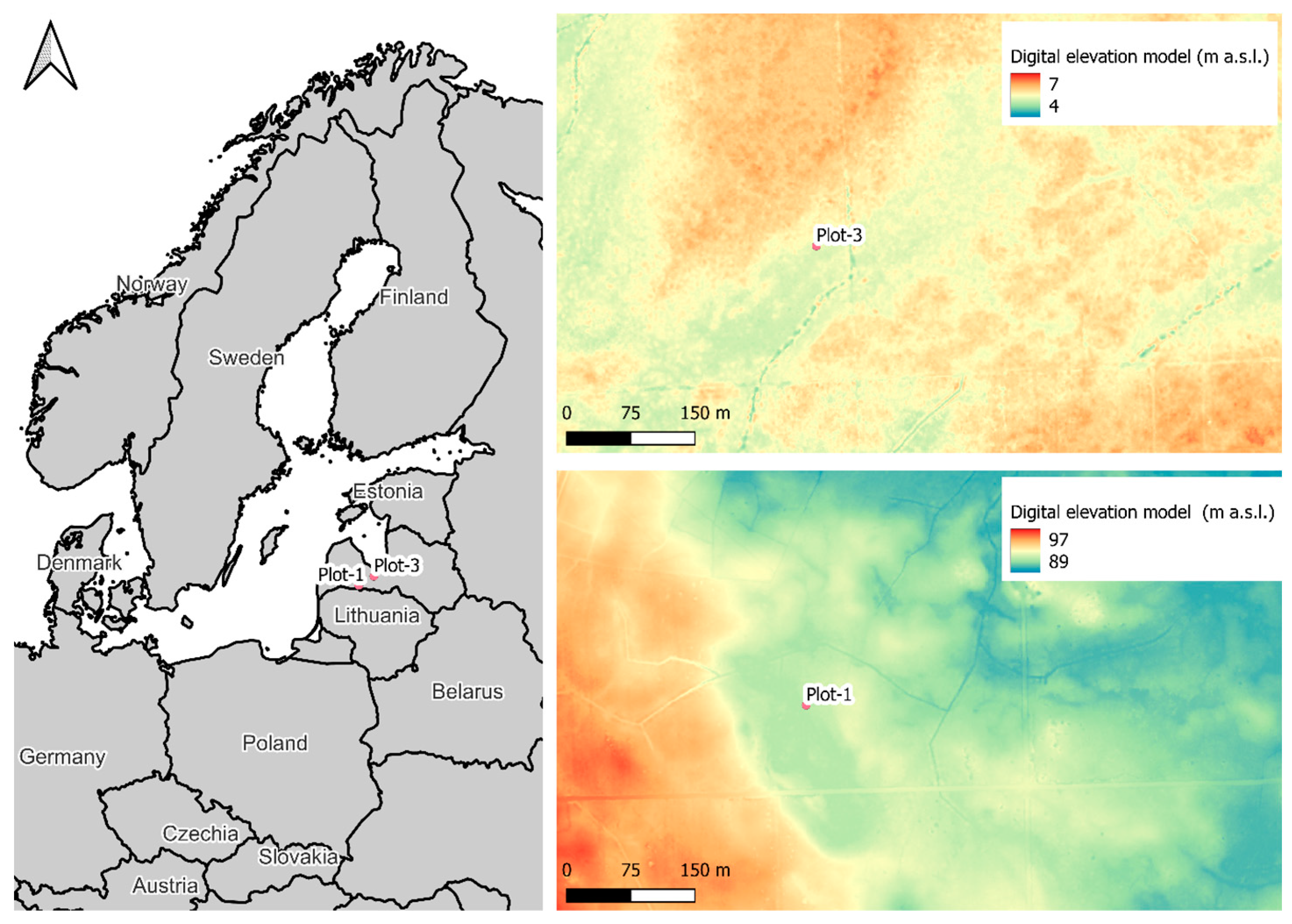
2.2. Study Site Description
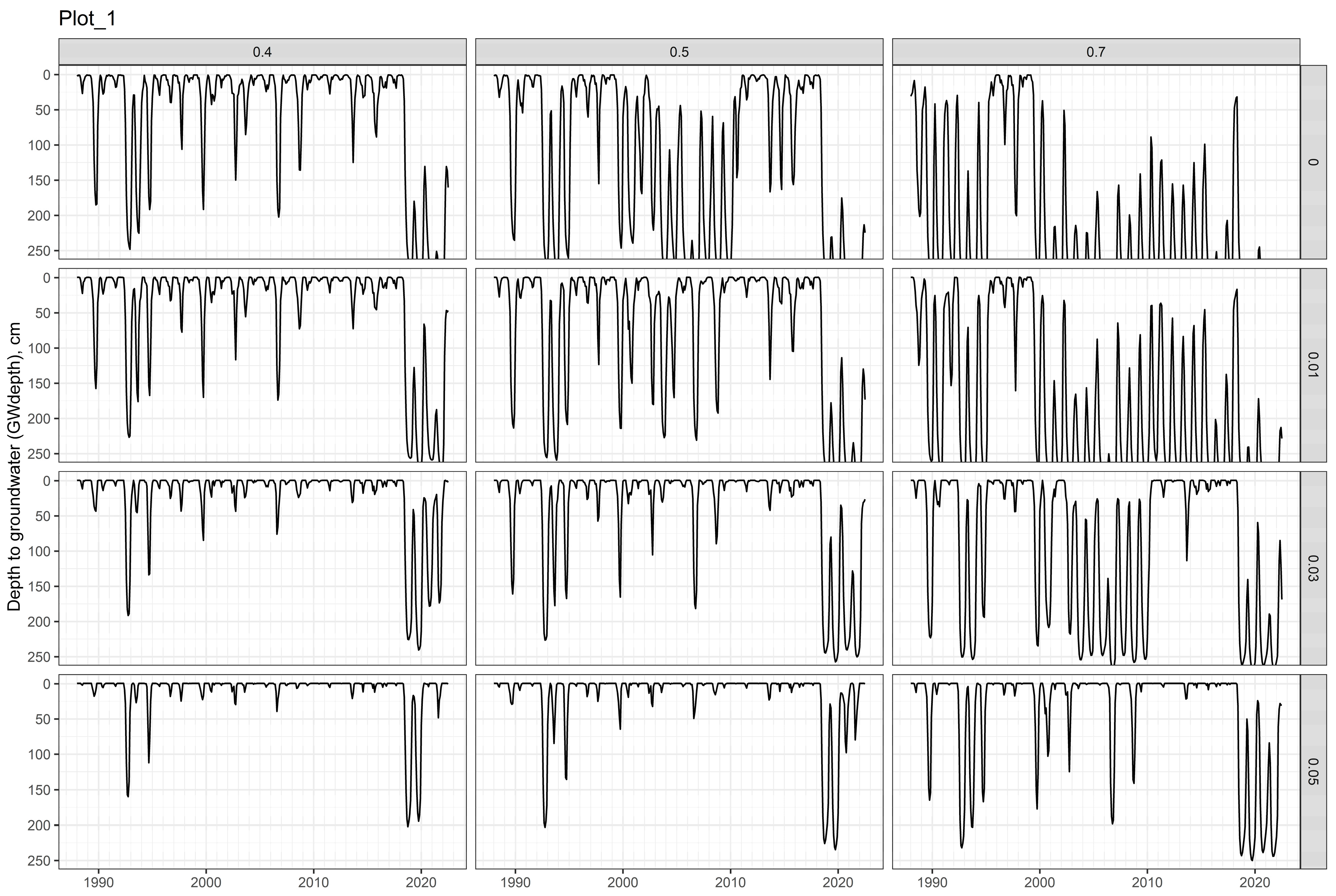
2.2.1. Plot-1
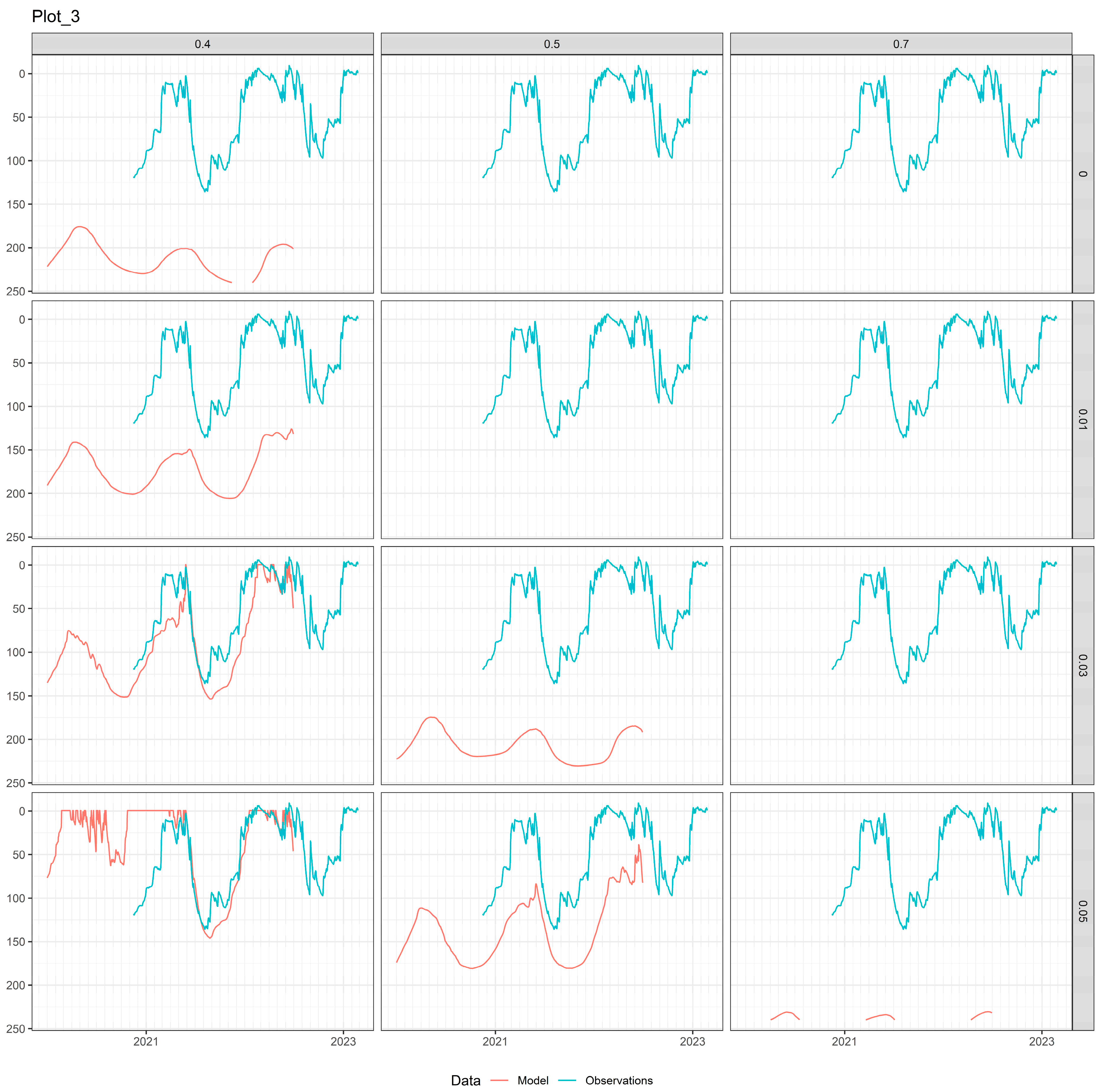
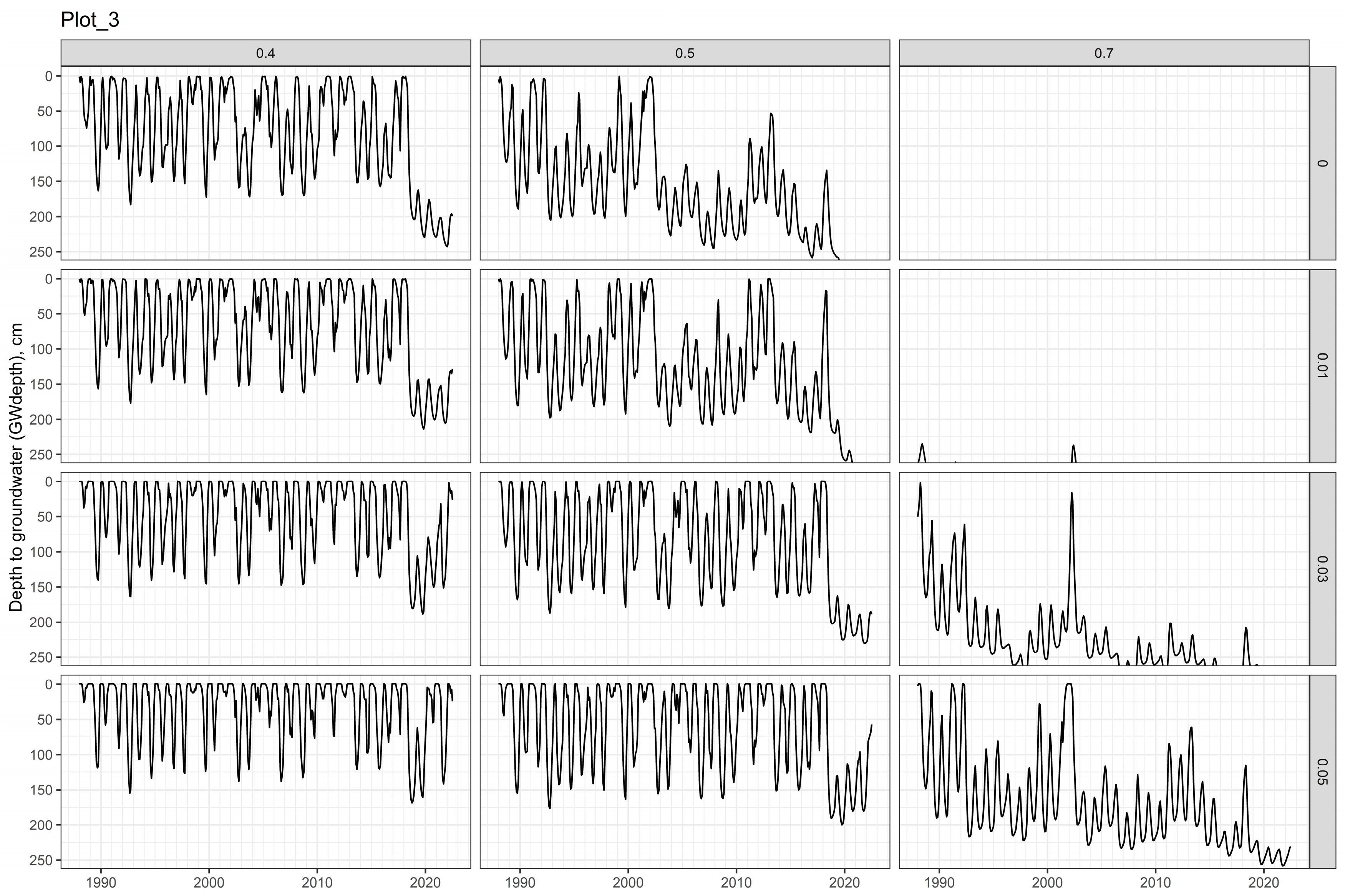
2.2.2. Plot-3
2.3. Soil Water Regime Observations
2.4. Model Setup
2.4.1. Modeling Scenarios: Model Instances
2.4.2. Model Calibration and Uncertainty
2.4.3. Soil Hydrological Properties
2.5. Meteorological Data: E-OBS
2.6. Root Depth Distribution
2.7. Root Water Uptake Parameterization
2.8. Leaf Area Index (LAI) Seasonal Trajectory Model
2.8.1. LAI Model
- As there were large gaps in the available LAI observations during winter, the winter background LAI value () was calculated as the average of a few available LAI observations between days of the year from 320 to 100. This value was interpreted as green parts of conifers and overwintering bryophytes.
- Continuous daily LAI time series were obtained by linearly interpolating the available LAI observations.
- The start of the vegetation season () was assumed to be the day of the year when the LAI value in spring exceeded that of the winter LAI by 1.5 ().
- Furthermore, all the available observed seasonal LAI trajectories were aligned to match the start of the vegetation season (); the master seasonal trajectory () was calculated as the daily median of the aligned LAI value for each day of the year except for winter (days of the year from 320 to 100).
- The start of the vegetation season () for each year was calculated using a simple degree day phenological model [64,72]. 1 January was set as the start of the degree day accumulation () and the phase onset was assumed to match the date when the active temperature sum () exceeded the predefined value for daily average temperature () and base temperature () (4):
- The aligned master trajectory () was compressed or stretched to match the fixed end date (31 December) via proportionally thinning (removing an appropriate number of evenly spread daily LAI data points) or upscaling (inserting an appropriate number of new daily data points interpolating the LAI value), respectively, for late or early springs.
- The most appropriate base temperature () and critical degree day () parameter set was selected by generating a range of base temperature () and critical degree day () values and selecting values that produced LAI trajectories with the least RMSE (root mean squared error) when compared to the observations.
- Finally, the LAI seasonal trajectory for the model period was calculated with daily average temperature from the E-OBS dataset as the input.
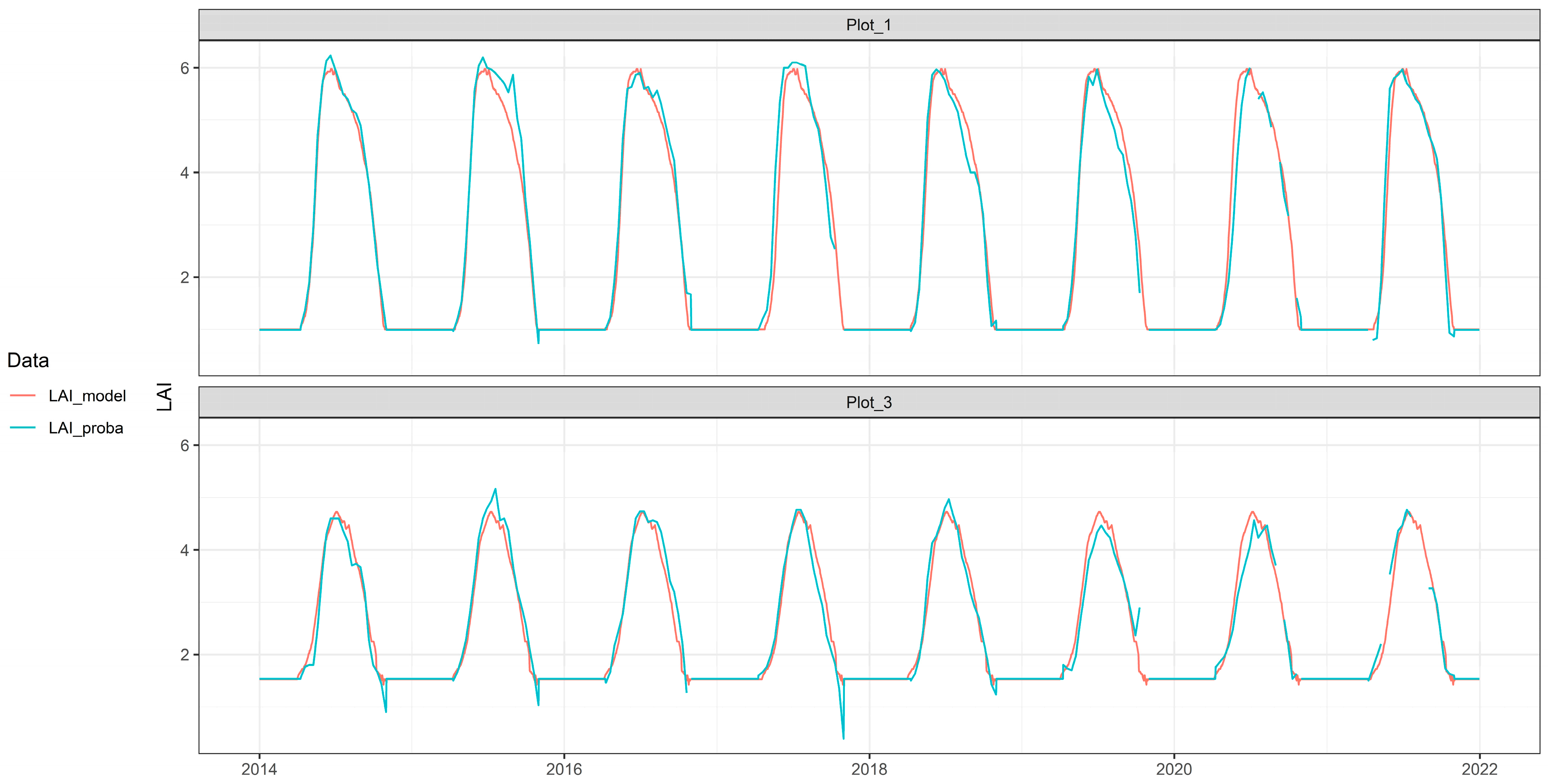
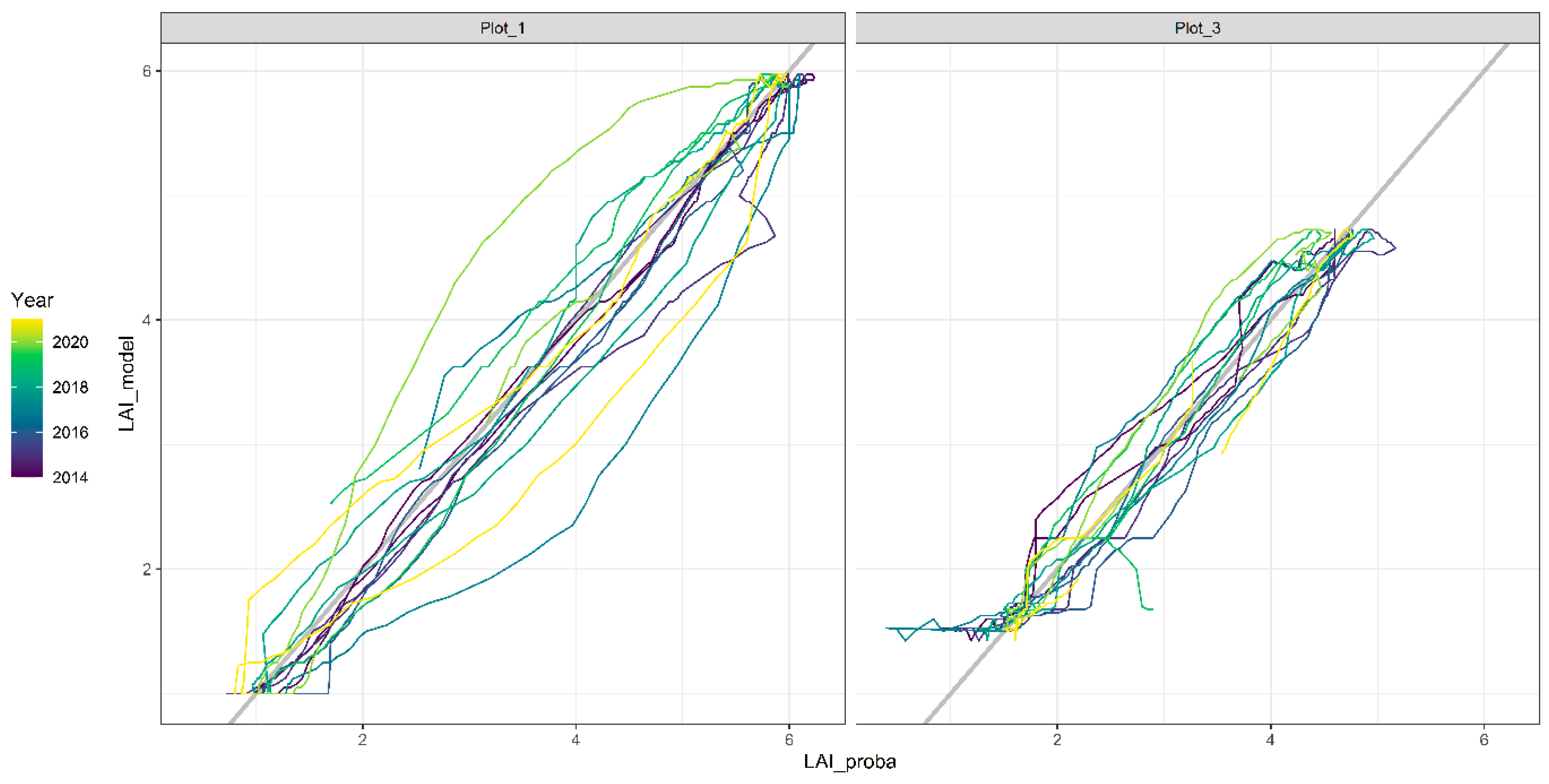
2.8.2. LAI Seasonal Model Results
2.9. Albedo
2.10. Interception
2.11. Tree-Ring Chronology
2.12. Analysis and Interpretation
3. Results
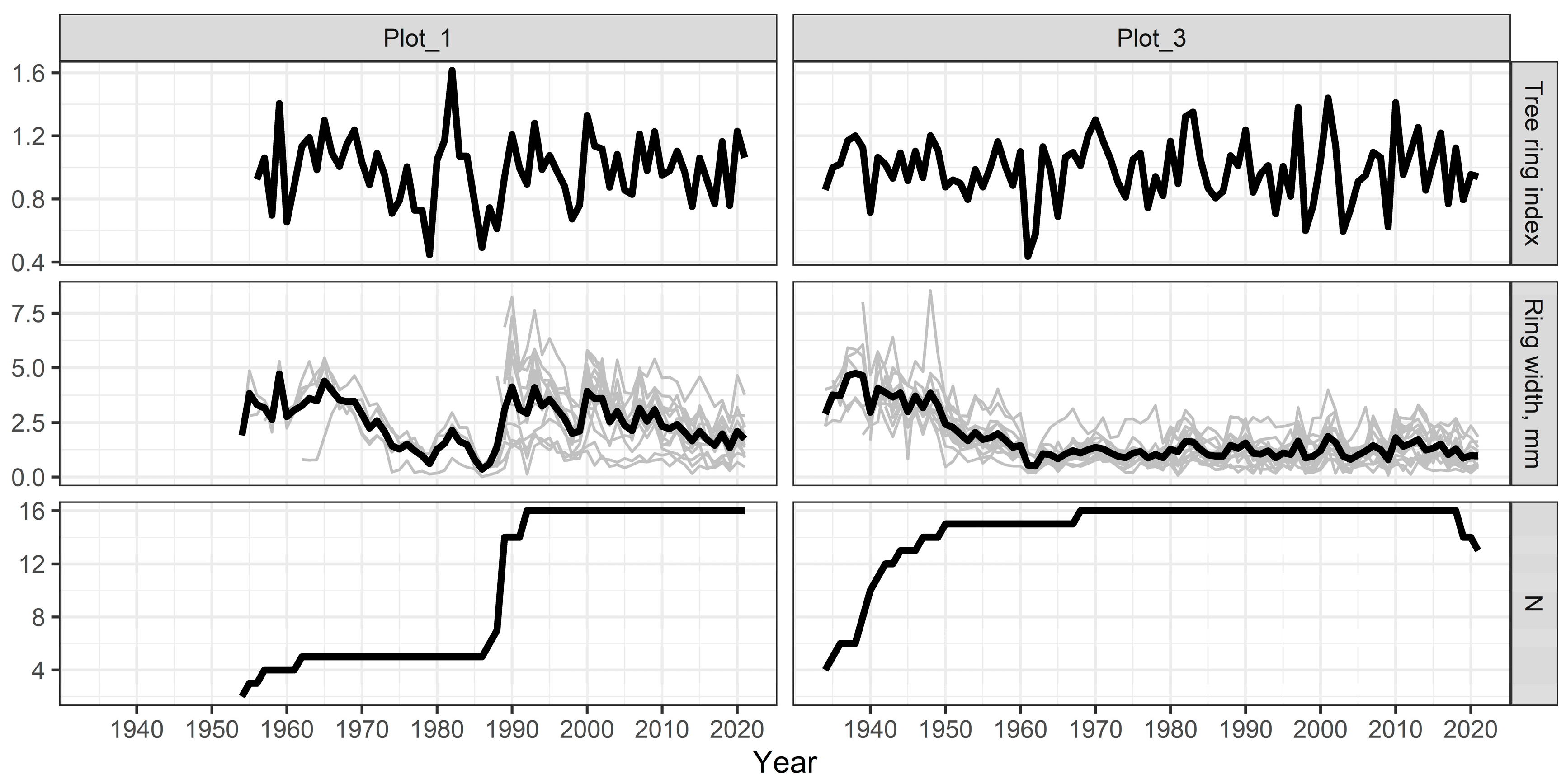
3.1. Tree-Ring Chronology
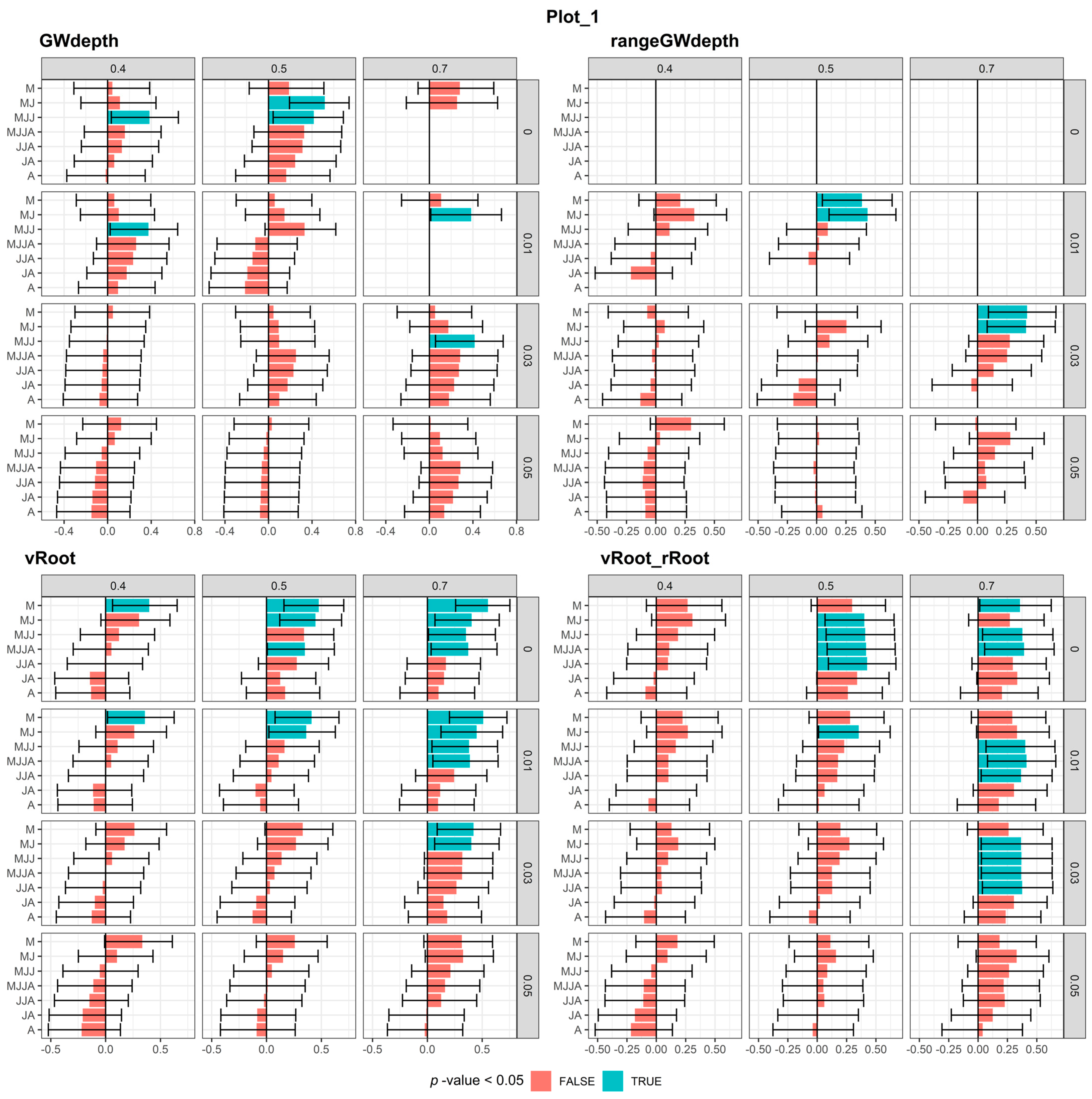
3.1.1. Plot-1
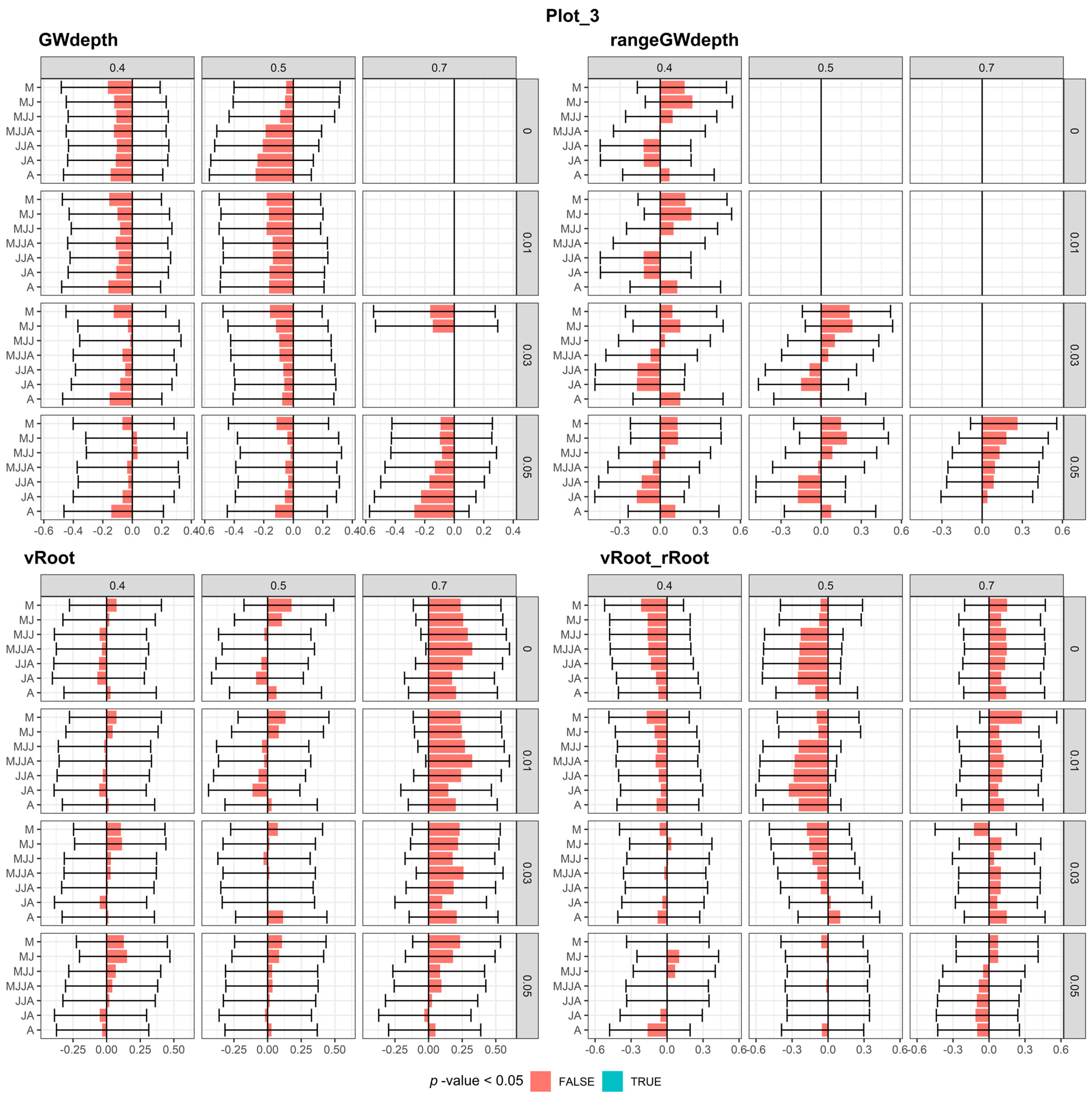
3.1.2. Plot-3
3.2. Soil Water Models
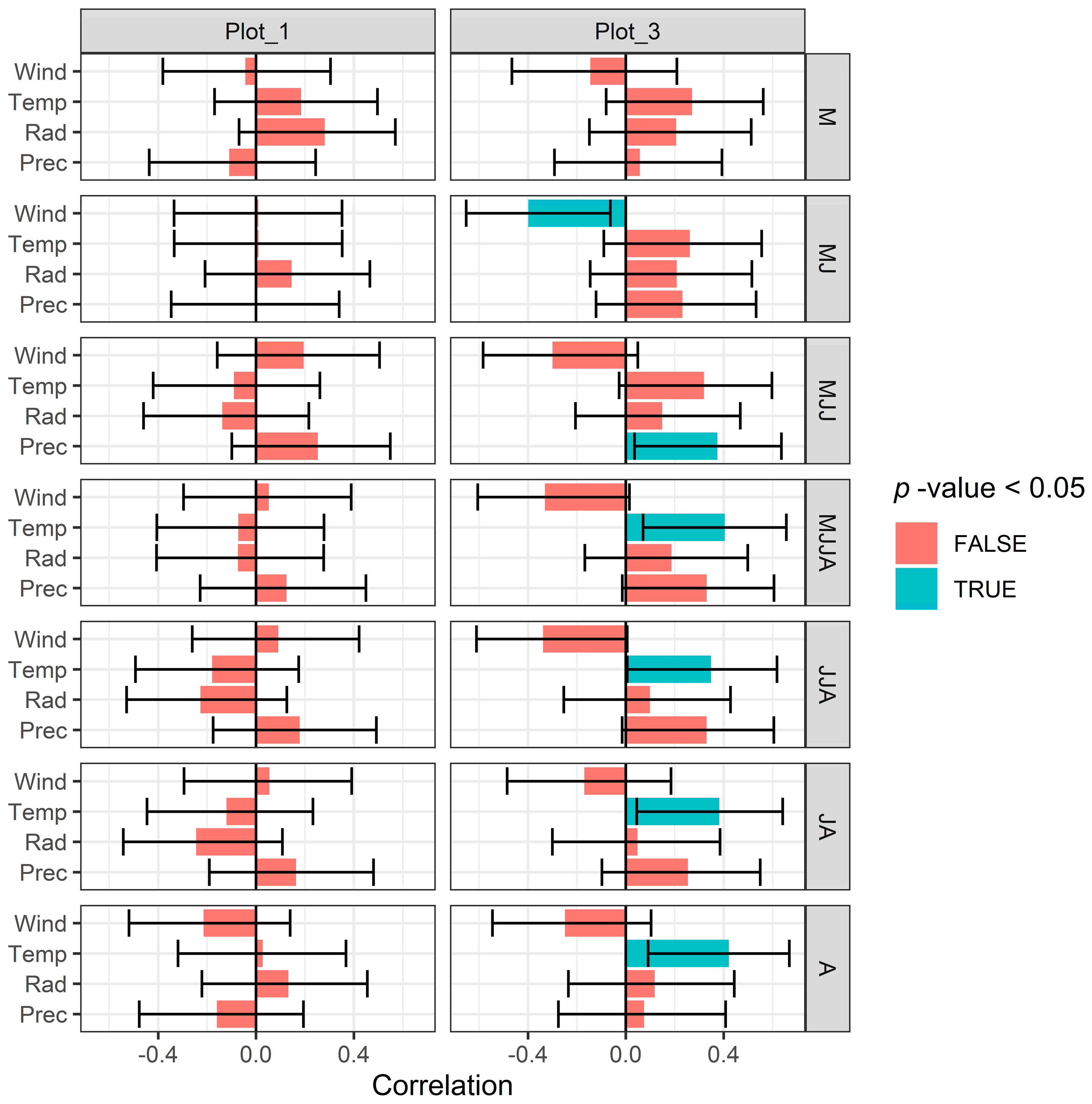
3.3. Correlation between Meteorological and Soil Water Conditions and Tree-Ring Chronology
3.3.1. Meteorological Conditions
3.3.2. Modeled Soil Water Conditions
4. Discussion
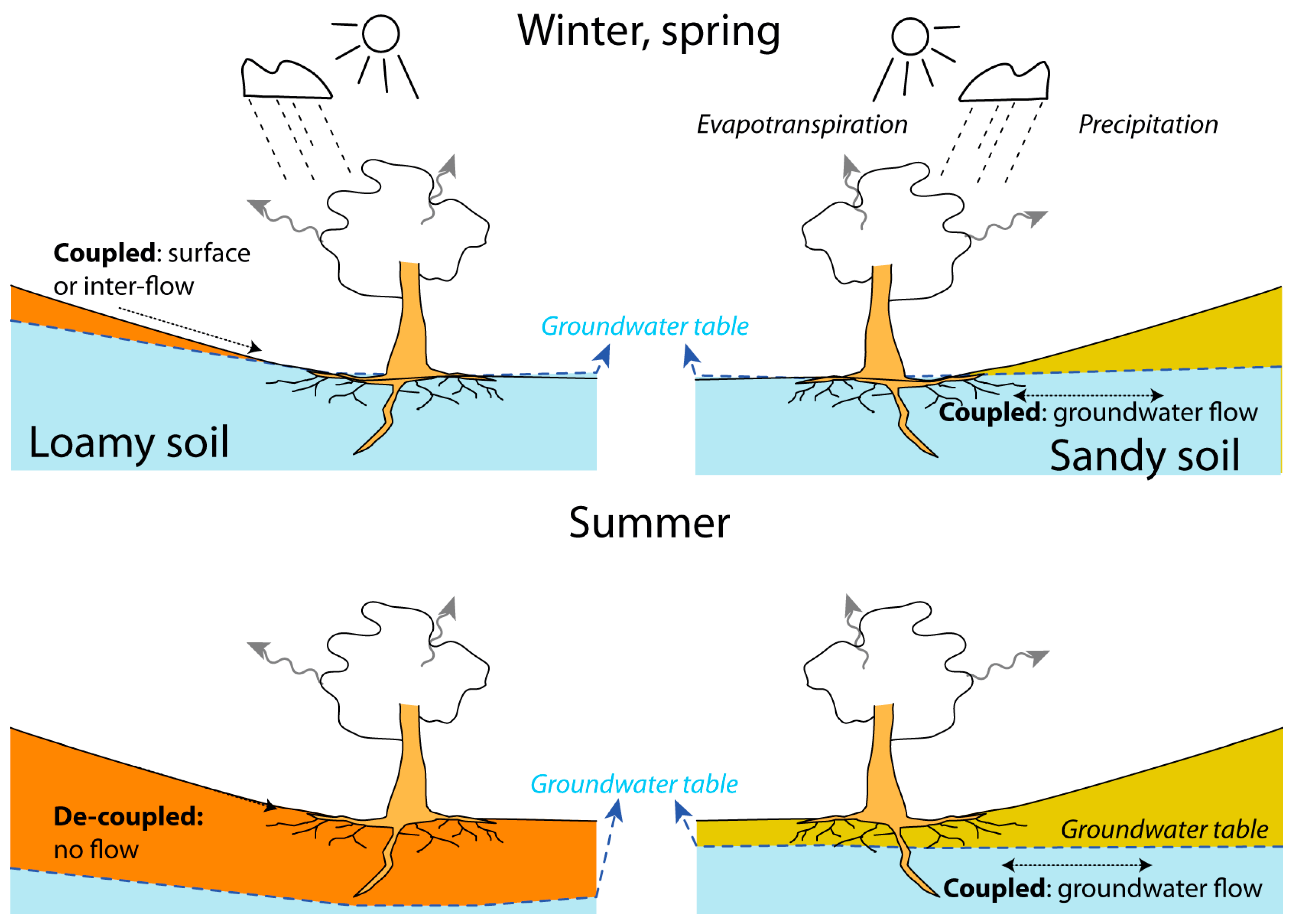
4.1. Hydrological Coupling and Decoupling
4.2. Implications of the Hydrological Coupling/Decoupling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Forzieri, G.; Dakos, V.; McDowell, N.G.; Ramdane, A.; Cescatti, A. Emerging Signals of Declining Forest Resilience under Climate Change. Nature 2022, 608, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Bermudez, R.; Montgomery, R.A.; Rich, R.L.; Rice, K.E.; Hobbie, S.E.; Stefanski, A. Even Modest Climate Change May Lead to Major Transitions in Boreal Forests. Nature 2022, 608, 540–545. [Google Scholar] [CrossRef] [PubMed]
- van der Velde, Y.; Temme, A.J.A.M.; Nijp, J.J.; Braakhekke, M.C.; van Voorn, G.A.K.; Dekker, S.C.; Dolman, A.J.; Wallinga, J.; Devito, K.J.; Kettridge, N.; et al. Emerging Forest–Peatland Bistability and Resilience of European Peatland Carbon Stores. Proc. Natl. Acad. Sci. USA 2021, 118, e2101742118. [Google Scholar] [CrossRef]
- Nave, L.E.; Gough, C.M.; Perry, C.H.; Hofmeister, K.L.; Le Moine, J.M.; Domke, G.M.; Swanston, C.W.; Nadelhoffer, K.J. Physiographic Factors Underlie Rates of Biomass Production during Succession in Great Lakes Forest Landscapes. For. Ecol. Manag. 2017, 397, 157–173. [Google Scholar] [CrossRef]
- Mustroph, A.; Albrecht, G. Tolerance of Crop Plants to Oxygen Deficiency Stress: Fermentative Activity and Photosynthetic Capacity of Entire Seedlings under Hypoxia and Anoxia. Physiol. Plant. 2003, 117, 508–520. [Google Scholar] [CrossRef]
- Mander, Ü.; Krasnova, A.; Schindler, T.; Megonigal, J.P.; Escuer-Gatius, J.; Espenberg, M.; Machacova, K.; Maddison, M.; Pärn, J.; Ranniku, R.; et al. Long-Term Dynamics of Soil, Tree Stem and Ecosystem Methane Fluxes in a Riparian Forest. Sci. Total Environ. 2022, 809, 151723. [Google Scholar] [CrossRef]
- Evans, C.D.; Peacock, M.; Baird, A.J.; Artz, R.R.E.; Burden, A.; Callaghan, N.; Chapman, P.J.; Cooper, H.M.; Coyle, M.; Craig, E.; et al. Overriding Water Table Control on Managed Peatland Greenhouse Gas Emissions. Nature 2021, 593, 548–552. [Google Scholar] [CrossRef]
- Zalitis, P.; Indriksons, A. The Hydrological Properties of Waterlogged and Drained Forests in Latvia. J. Water Land Dev. 2009, 13, 69–86. [Google Scholar] [CrossRef]
- Nieminen, M.; Hökkä, H.; Laiho, R.; Juutinen, A.; Ahtikoski, A.; Pearson, M.; Kojola, S.; Sarkkola, S.; Launiainen, S.; Valkonen, S.; et al. Could Continuous Cover Forestry Be an Economically and Environmentally Feasible Management Option on Drained Boreal Peatlands? For. Ecol. Manag. 2018, 424, 78–84. [Google Scholar] [CrossRef]
- Laudon, H.; Hasselquist, E.M. Applying Continuous-Cover Forestry on Drained Boreal Peatlands; Water Regulation, Biodiversity, Climate Benefits and Remaining Uncertainties. Trees For. People 2023, 11, 100363. [Google Scholar] [CrossRef]
- Devito, K.J.; Hokanson, K.J.; Moore, P.A.; Kettridge, N.; Anderson, A.E.; Chasmer, L.; Hopkinson, C.; Lukenbach, M.C.; Mendoza, C.A.; Morissette, J.; et al. Landscape Controls on Long-Term Runoff in Subhumid Heterogeneous Boreal Plains Catchments. Hydrol. Process. 2017, 31, 2737–2751. [Google Scholar] [CrossRef]
- Gribovszki, Z.; Kalicz, P.; Palocz-Andresen, M.; Szalay, D.; Varga, T. Hydrological Role of Central European Forests in Changing Climate –Review. Idojaras 2019, 123, 535–550. [Google Scholar] [CrossRef]
- Tóth, J. A Theoretical Analysis of Groundwater Flow in Small Drainage Basins. J. Geophys. Res. 1963, 68, 4795–4812. [Google Scholar] [CrossRef]
- Hokanson, K.J.; Peterson, E.S.; Devito, K.J.; Mendoza, C.A. Forestland-Peatland Hydrologic Connectivity in Water-Limited Environments: Hydraulic Gradients Often Oppose Topography. Environ. Res. Lett. 2020, 15, 034021. [Google Scholar] [CrossRef]
- Komatsu, H.; Kume, T. Modeling of Evapotranspiration Changes with Forest Management Practices: A Genealogical Review. J. Hydrol. 2020, 585, 124835. [Google Scholar] [CrossRef]
- Tor-ngern, P.; Oren, R.; Palmroth, S.; Novick, K.; Oishi, A.; Linder, S.; Ottosson-Löfvenius, M.; Näsholm, T. Water Balance of Pine Forests: Synthesis of New and Published Results. Agric. For. Meteorol. 2018, 259, 107–117. [Google Scholar] [CrossRef]
- Finzi, A.C.; Giasson, M.A.; Barker Plotkin, A.A.; Aber, J.D.; Boose, E.R.; Davidson, E.A.; Dietze, M.C.; Ellison, A.M.; Frey, S.D.; Goldman, E.; et al. Carbon Budget of the Harvard Forest Long-Term Ecological Research Site: Pattern, Process, and Response to Global Change. Ecol. Monogr. 2020, 90, e01423. [Google Scholar] [CrossRef]
- Dow, C.; Kim, A.Y.; D’Orangeville, L.; Gonzalez-Akre, E.B.; Helcoski, R.; Herrmann, V.; Harley, G.L.; Maxwell, J.T.; McGregor, I.R.; McShea, W.J.; et al. Warm Springs Alter Timing but Not Total Growth of Temperate Deciduous Trees. Nature 2022, 608, 552–557. [Google Scholar] [CrossRef]
- Šimůnek, J.; Šejna, M.; Saitoh, T.M.; Sakai, M.; van Genuchten, M.T. The HYDRUS-1D Software Package for Simulating the One-Dimensional Movement of Water, Heat, and Multiple Solutes in Variably-Saturated Media. In University of California-Riverside Research Reports; University of California: Los Angeles, CA, USA, 2013; p. 343. [Google Scholar]
- Kalvāns, A.; Dauškane, I. Data Set Supporting Publication “Seasonal Hydrological Coupling-Decoupling as a Key Element in Soil Water Regime of Hydric Forests” Submitted for the Journal of Hydrology by Kalvāns A. and Dauškāne I. [Data Set]. Zenodo 2023. [Google Scholar] [CrossRef]
- LĢIA Digital Height Model Basic Data. Available online: https://www.lgia.gov.lv/en/Digitālaisvirsmasmodelis (accessed on 14 October 2021).
- Jaagus, J.; Briede, A.; Rimkus, E.; Remm, K. Variability and Trends in Daily Minimum and Maximum Temperatures and in the Diurnal Temperature Range in Lithuania, Latvia and Estonia in 1951–2010. Theor. Appl. Climatol. 2014, 118, 57–68. [Google Scholar] [CrossRef]
- Kalvāns, A.; Kalvāne, G.; Zandersons, V.; Gaile, D.; Briede, A. Recent Seasonally Contrasting and Persistent Recent Warming Trends in Latvia. Theor. Appl. Climatol. 2023. [Google Scholar] [CrossRef]
- Jaagus, J.; Briede, A.; Rimkus, E.; Sepp, M. Changes in Precipitation Regime in the Baltic Countries in 1966–2015. Theor. Appl. Climatol. 2018, 131, 433–443. [Google Scholar] [CrossRef]
- Jaagus, J.; Aasa, A.; Aniskevich, S.; Boincean, B.; Bojariu, R.; Briede, A.; Danilovich, I.; Castro, F.D.; Dumitrescu, A.; Labuda, M.; et al. Long-term Changes in Drought Indices in Eastern and Central Europe. Int. J. Climatol. 2022, 42, 225–249. [Google Scholar] [CrossRef]
- Keith, D.A.; Ferrer-Paris, J.R.; Nicholson, E.; Bishop, M.J.; Polidoro, B.A.; Ramirez-Llodra, E.; Tozer, M.G.; Nel, J.L.; Mac Nally, R.; Gregr, E.J.; et al. A Function-Based Typology for Earth’s Ecosystems. Nature 2022, 610, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Keith, D.A.; Ferrer-paris, J.R.; Nicholson, E.; Kingsford, R.T. IUCN Global Ecosystem Typology 2.0: Descriptive Profiles for Biomes and Ecosystem Functional Groups; IUCN, International Union for Conservation of Nature: Gland, Switzerland, 2020; ISBN 9782831720777. [Google Scholar]
- Burton, T.M. Swamps—Wooded Wetlands. Encycl. Inl. Waters 2009, 549–557. [Google Scholar] [CrossRef]
- Misans, J.; Murnieks, A.; Strautnieks, I. Latvijas Geologiska Karte, Merogs 1:200000. 32.Lapa-Jelgava (Paskaidrojuma Teksts Un Kartes) (in Latvian) Latvian State Geological Fund No. 12444; 2001; Available online: https://www.meteo.lv/en/pakalpojumi/geologiska-un-hidrogeologiska-informacija/kartes/latvijas-geologiska-karte-kvartara-nogulumu-karte-meroga-1-200000?id=3339&cid=1037 (accessed on 24 August 2023).
- Takčidi, E. Datu Bāzes “Urbumi” Dokumentācija [Documentation of the Database “Boreholes”]; State Geological Survey: Riga, Latvia, 1999. [Google Scholar]
- Spalviņš, A.; Krauklis, K.; Aleksāns, O.; Lāce, I. Latvijas Hidroģeoloģiskā Modeļa LAMO Izveidošana, Izmantošana Un Pilnveidošana (Development, Application and Upgrading of the Hydrogeological Model of Latvia LAMO; in Latvian). Bound. Field Probl. Comput. Simul. 2018, 57, 5–14. [Google Scholar] [CrossRef]
- Valtera, M.; Schaetzl, R.J. Pit-Mound Microrelief in Forest Soils: Review of Implications for Water Retention and Hydrologic Modelling. For. Ecol. Manag. 2017, 393, 40–51. [Google Scholar] [CrossRef]
- Megonigal, J.P.; Patrick, W.H.; Faulkner, S.P. Wetland Identification in Seasonally Flooded Forest Soils: Soil Morphology and Redox Dynamics. Soil Sci. Soc. Am. J. 1993, 57, 140–149. [Google Scholar] [CrossRef]
- Virbulis, J.; Bethers, U.; Saks, T.; Sennikovs, J.; Timuhins, A. Hydrogeological Model of the Baltic Artesian Basin. Hydrogeol. J. 2013, 21, 845–862. [Google Scholar] [CrossRef]
- Monteith, J.L. Evaporation and Surface Temperature. Q. J. R. Meteorol. Soc. 1981, 107, 1–27. [Google Scholar] [CrossRef]
- McMahon, T.A.; Peel, M.C.; Lowe, L.; Srikanthan, R.; McVicar, T.R. Estimating Actual, Potential, Reference Crop and Pan Evaporation Using Standard Meteorological Data: A Pragmatic Synthesis. Hydrol. Earth Syst. Sci. 2013, 17, 1331–1363. [Google Scholar] [CrossRef]
- Zālītis, P. Mežs Un Ūdens; Laiviņš, M., Ed.; Latviajs Valsts mežzinātnes institūts “Silava”: Salaspils, Latvia, 2012; Volume 356, ISBN 9789934801662. [Google Scholar]
- Kalvans, A. Run-on Contribution to the Soil Water Balance to the Temperate Forests. In Proceedings of the XXXI Nordic Hydrological Conference, Tallinn, Estonia, 15–18 August 2022. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration-Guidelines for Computing Crop Water Requirements-FAO Irrigation and Drainage Paper 56; FAO: Rome, Italy, 1998; ISBN 92-5-104219-5. [Google Scholar]
- Wolfs, D.; Verger, A.; Van der Goten, R.; Sánchez-Zapero, J. Product User Manual Leaf Area Index (LAI), Fraction of Absorbed Photosynthetically Active Radiation (FAPAR) Fraction of Green Vegetation Cover (FCOVER) Collection 300m, version 1.1; VITO: Mol, Belgium, 2021. [Google Scholar]
- Sutton, O.F.; Price, J.S. Soil Moisture Dynamics Modelling of a Reclaimed Upland in the Early Post-Construction Period. Sci. Total Environ. 2020, 718, 134628. [Google Scholar] [CrossRef]
- Eijkelkamp. 09.02 Laboratory-Permeameters, Operating Instructions, Brand Data Laboratory Permeameter; Eijkelkamp: Giesbeek, The Netherlands, 2013; Available online: https://en.eijkelkamp.com/products/laboratory-equipment/soil-water-permeameters.html (accessed on 10 September 2022).
- Schindler, U.; Durner, W.; von Unold, G.; Müller, L. Evaporation Method for Measuring Unsaturated Hydraulic Properties of Soils: Extending the Measurement Range. Soil Sci. Soc. Am. J. 2010, 74, 1071–1083. [Google Scholar] [CrossRef]
- Schindler, U.; Durner, W.; von Unold, G.; Mueller, L.; Wieland, R. The Evaporation Method: Extending the Measurement Range of Soil Hydraulic Properties Using the Air-Entry Pressure of the Ceramic Cup. J. Plant Nutr. Soil Sci. 2010, 173, 563–572. [Google Scholar] [CrossRef]
- van Genuchten, M. A Closed Form Equation for Predicting the Hydraulic Conductivity of Unsaturated Soils. Soil Sci. Soc. Am. J. 1980, 44, 892–898. [Google Scholar] [CrossRef]
- Pertassek, T.; Peters, A.; Durner, W. HYPROP-FIT User’s Manual, version 3.1; METER Group AG: München, Germany, 2017; p. 66. [Google Scholar]
- Cornes, R.C.; van der Schrier, G.; van den Besselaar, E.J.M.; Jones, P.D. An Ensemble Version of the E-OBS Temperature and Precipitation Data Sets. J. Geophys. Res. Atmos. 2018, 123, 9391–9409. [Google Scholar] [CrossRef]
- Fan, Y.; Miguez-Macho, G.; Jobbágy, E.G.; Jackson, R.B.; Otero-Casal, C. Hydrologic Regulation of Plant Rooting Depth. Proc. Natl. Acad. Sci. USA 2017, 114, 10572–10577. [Google Scholar] [CrossRef]
- Gale, M.R.; Grigal, D.F. Vertical Root Distributions of Northern Tree Species in Relation to Successional Status. Can. J. For. Res. 1987, 17, 829–834. [Google Scholar] [CrossRef]
- Jackson, R.B.; Mooney, H.A.; Schulze, E.D. A Global Budget for Fine Root Biomass, Surface Area, and Nutrient Contents. Proc. Natl. Acad. Sci. USA 1997, 94, 7362–7366. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, M.A.; De Jong Van Lier, Q.; Van Dam, J.C.; Bezerra, A.H.F. Benchmarking Test of Empirical Root Water Uptake Models. Hydrol. Earth Syst. Sci. 2017, 21, 473–493. [Google Scholar] [CrossRef]
- Carminati, A.; Javaux, M. Soil Rather Than Xylem Vulnerability Controls Stomatal Response to Drought. Trends Plant Sci. 2020, 25, 868–880. [Google Scholar] [CrossRef] [PubMed]
- Feddes, R.A.; Kowalik, P.; Kolinska-Malinka, K.; Zaradny, H. Simulation of Field Water Uptake by Plants Using a Soil Water Dependent Root Extraction Function. J. Hydrol. 1976, 31, 13–26. [Google Scholar] [CrossRef]
- Jarvis, N.J. A Simple Empirical Model of Root Water Uptake. J. Hydrol. 1989, 107, 57–72. [Google Scholar] [CrossRef]
- Jarvis, N.J. Simple Physics-Based Models of Compensatory Plant Water Uptake: Concepts and Eco-Hydrological Consequences. Hydrol. Earth Syst. Sci. 2011, 15, 3431–3446. [Google Scholar] [CrossRef]
- Sutton, O.F.; Price, J.S. Modelling the Hydrologic Effects of Vegetation Growth on the Long-Term Trajectory of a Reclamation Watershed. Sci. Total Environ. 2020, 734, 139323. [Google Scholar] [CrossRef]
- van Genuchten, M.T. A Numerical Model for Water and Solute Movement in and below the Root Zone; United States Department of Agriculture Agricultural Research Service US Salinity Laboratory: Riverside, CA, USA, 1987. [Google Scholar]
- de Jong van Lier, Q.; van Dam, J.C.; Durigon, A.; dos Santos, M.A.; Metselaar, K. Modeling Water Potentials and Flows in the Soil-Plant System Comparing Hydraulic Resistances and Transpiration Reduction Functions. Vadose Zone J. 2013, 12, vzj2013.02.0039. [Google Scholar] [CrossRef]
- Gong, D.; Kang, S.; Zhang, L.; Du, T.; Yao, L. A Two-Dimensional Model of Root Water Uptake for Single Apple Trees and Its Verification with Sap Flow and Soil Water Content Measurements. Agric. Water Manag. 2006, 83, 119–129. [Google Scholar] [CrossRef]
- Green, S.R.; Vogeler, I.; Clothier, B.E.; Mills, T.M.; Dijssel, C. van den Modelling Water Uptake by a Mature Apple Tree. Soil Res. 2003, 41, 365. [Google Scholar] [CrossRef]
- McVean, D.N. Ecology of Alnus glutinosa (L.) Gaertn: IV. Root System. J. Ecol. 1956, 44, 219. [Google Scholar] [CrossRef]
- Eschenbach, C.; Kappen, L. Leaf Water Relations of Black Alder [Alnus glutinosa (L.) Gaertn.] Growing at Neighbouring Sites with Different Water Regimes. Trees 1999, 14, 28–38. [Google Scholar] [CrossRef]
- Copernicus Global Land Service Leaf Area Index. Available online: https://land.copernicus.eu/global/products/lai (accessed on 14 February 2022).
- Kalvāns, A.; Bitāne, M.; Kalvāne, G. Forecasting Plant Phenology: Evaluating the Phenological Models for Betula Pendula and Padus Racemosa Spring Phases, Latvia. Int. J. Biometeorol. 2015, 59, 165–179. [Google Scholar] [CrossRef] [PubMed]
- Menzel, A.; Yuan, Y.; Matiu, M.; Sparks, T.; Scheifinger, H.; Gehrig, R.; Estrella, N. Climate Change Fingerprints in Recent European Plant Phenology. Glob. Change Biol. 2020, 26, 2599–2612. [Google Scholar] [CrossRef] [PubMed]
- Kalvāne, G.; Kalvāns, A. Phenological Trends of Multi-Taxonomic Groups in Latvia, 1970–2018. Int. J. Biometeorol. 2021, 65, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Laube, J.; Sparks, T.H.; Estrella, N.; Menzel, A. Does Humidity Trigger Tree Phenology? Proposal for an Air Humidity Based Framework for Bud Development in Spring. New Phytol. 2014, 202, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Delpierre, N.; Dufrêne, E.; Soudani, K.; Ulrich, E.; Cecchini, S.; Boé, J.; François, C. Modelling Interannual and Spatial Variability of Leaf Senescence for Three Deciduous Tree Species in France. Agric. For. Meteorol. 2009, 149, 938–948. [Google Scholar] [CrossRef]
- Fracheboud, Y.; Luquez, V.; Bjorken, L.; Sjodin, A.; Tuominen, H.; Jansson, S. The Control of Autumn Senescence in European Aspen. Plant Physiol. 2009, 149, 1982–1991. [Google Scholar] [CrossRef]
- Zani, D.; Crowther, T.W.; Mo, L.; Renner, S.S.; Zohner, C.M. Increased Growing-Season Productivity Drives Earlier Autumn Leaf Senescence in Temperate Trees. Science 2020, 370, 1066–1071. [Google Scholar] [CrossRef]
- Kumar, R.; Bishop, E.; Bridges, W.C.; Tharayil, N.; Sekhon, R.S. Sugar Partitioning and Source–Sink Interaction Are Key Determinants of Leaf Senescence in Maize. Plant Cell Environ. 2019, 42, 2597–2611. [Google Scholar] [CrossRef]
- Herms, D. Using Degree-Days and Plant Phenology to Predict Pest Activity. In IPM (Integrated Pest Management) of Midwest Landscapes; Krischik, V., Davidson, J., Eds.; Minnesota Agricultural Experiment Station Publication: Saint Paul, MN, USA, 2004; pp. 49–59. [Google Scholar]
- Smets, B.; Sánchez-Zapero, J. Vegetation and Energy, Product User Manual, Surface Albedo, Collection 1km, Version 1; VITO: Mol, Belgium, 2018; Volume 93. [Google Scholar]
- Copernicus Global Land Service. Surface Albedo. 2020. Available online: https://land.copernicus.eu/global/products/sa (accessed on 9 September 2022).
- Van Dijk, A.I.J.M.; Gash, J.H.; Van Gorsel, E.; Blanken, P.D.; Cescatti, A.; Emmel, C.; Gielen, B.; Harman, I.N.; Kiely, G.; Merbold, L.; et al. Rainfall Interception and the Coupled Surface Water and Energy Balance. Agric. For. Meteorol. 2015, 214, 402–415. [Google Scholar] [CrossRef]
- van Dam, J.C.; Huygen, J.; Wesseling, J.G.; Feddes, R.A.; Kabat, P.; van Walsum, P.E.V.; Groenendijk, P.; van Diepen, C.A. Theory of SWAP version 2.0 Simulation of water flow, solute transport and plant growth in the Soil-Water-Atmosphere-Plant environment. Report 71; Department Water Resources, Wageningen Agricultural University: Wageningen, The Netherlands, 1997. [Google Scholar]
- Šípek, V.; Hnilica, J.; Vlček, L.; Hnilicová, S.; Tesař, M. Influence of Vegetation Type and Soil Properties on Soil Water Dynamics in the Šumava Mountains (Southern Bohemia). J. Hydrol. 2020, 582, 124285. [Google Scholar] [CrossRef]
- Dohnal, M.; Černý, T.; Votrubová, J.; Tesař, M. Rainfall Interception and Spatial Variability of Throughfall in Spruce Stand. J. Hydrol. Hydromech. 2014, 62, 277–284. [Google Scholar] [CrossRef]
- Grissino-Mayer, H. A Manual and Tutorial for the Proper Use of an Increment Borer. Tree-Ring Res. 2003, 59, 63–79. [Google Scholar]
- Rinn, F. TSAP-Win: Time Series Analysis and Presentation for Dendrochronology and Related Applications, version 0.55; User Reference: Heidelberg, Germany, 2003. [Google Scholar]
- Holmes, R.L. Computer-Assisted Quality Control in Tree-Ring Dating and Measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Bunn, A.G. A Dendrochronology Program Library in R (DplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011; Volume 1, p. 409. [Google Scholar]
- Holmes, R.L. Dendrochronology Program Library (DPL) Users Manual; The University of Arizona: Tuscon, Arizona, 1999. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Laganis, J.; Pečkov, A.; Debeljak, M. Modeling Radial Growth Increment of Black Alder (Alnus glutionsa (L.) Gaertn.) Tree. Ecol. Model. 2008, 215, 180–189. [Google Scholar] [CrossRef]
- Babre, A.; Kalvāns, A.; Avotniece, Z.; Retiķe, I.; Bikše, J.; Jemeljanova, M.; Popovs, K.; Zelenkevičs, A.; Dēliņa, A. The Use of Predefined Drought Indices for the Assessment of Groundwater Drought Episodes in the Baltic States over the Period 1989–2018. J. Hydrol. Reg. Stud. 2022, 40, 101049. [Google Scholar] [CrossRef]
- Schnabel, F.; Purrucker, S.; Schmitt, L.; Engelmann, R.A.; Kahl, A.; Richter, R.; Seele, C.; Skiadaresis, G.; Wirth, C. Cumulative Growth and Stress Responses to the 2018–2019 Drought in a European Floodplain Forest. Glob. Change Biol. 2022, 28, 1870–1883. [Google Scholar] [CrossRef]
- Skaggs, R.W.; Amatya, D.M.; Chescheir, G.M. Effects of Drainage for Silviculture on Wetland Hydrology. Wetlands 2020, 40, 47–64. [Google Scholar] [CrossRef]
- Miguez-Macho, G.; Fan, Y. Spatiotemporal Origin of Soil Water Taken up by Vegetation. Nature 2021, 598, 624–628. [Google Scholar] [CrossRef]
- Vainu, M.; Terasmaa, J. Changes in Climate, Catchment Vegetation and Hydrogeology as the Causes of Dramatic Lake-Level Fluctuations in the Kurtna Lake District, NE Estonia. Est. J. Earth Sci. 2014, 63, 45–61. [Google Scholar] [CrossRef]
- Kalvāns, A.; Popovs, K.; Priede, A.; Koit, O.; Retiķe, I.; Bikše, J.; Dēliņa, A.; Babre, A. Nitrate Vulnerability of Karst Aquifers and Associated Groundwater-Dependent Ecosystems in the Baltic Region. Environ. Earth Sci. 2021, 80, 628. [Google Scholar] [CrossRef]
- Koit, O.; Tarros, S.; Pärn, J.; Küttim, M.; Abreldaal, P.; Sisask, K.; Vainu, M.; Terasmaa, J.; Retike, I.; Polikarpus, M. Contribution of Local Factors to the Status of a Groundwater Dependent Terrestrial Ecosystem in the Transboundary Gauja-Koiva River Basin, North-Eastern Europe. J. Hydrol. 2021, 600, 126656. [Google Scholar] [CrossRef]
- Van Stempvoort, D.R.; MacKay, D.R.; Koehler, G.; Collins, P.; Brown, S.J. Subsurface Hydrology of Tile-drained Headwater Catchments: Compatibility of Concepts and Hydrochemistry. Hydrol. Process. 2021, 35, e14342. [Google Scholar] [CrossRef]
- Liu, Y.; Du, J.; Xu, X.; Kardol, P.; Hu, D. Microtopography-Induced Ecohydrological Effects Alter Plant Community Structure. Geoderma 2020, 362, 114119. [Google Scholar] [CrossRef]
- Kalvāns, A.; Kalvāne, G. Soil Waterlogging Stress Compensated by Root System Adaptation in a Pot Experiment with Sweet Corn Zea Mays Var. Saccharate. In Proceedings of the 22nd International Multidisciplinary Scientific GeoConference SGEM 2022, Issue, 3.1. Albena, Bulgaria, 4–10 June 2022; Trofymchuk, O., Rivza, B., Eds.; STEF92 Technology: Vienna, Austria, 2022; pp. 167–178. [Google Scholar]
- Solly, E.F.; Brunner, I.; Helmisaari, H.S.; Herzog, C.; Leppälammi-Kujansuu, J.; Schöning, I.; Schrumpf, M.; Schweingruber, F.H.; Trumbore, S.E.; Hagedorn, F. Unravelling the Age of Fine Roots of Temperate and Boreal Forests. Nat. Commun. 2018, 9, 3006. [Google Scholar] [CrossRef]
- Haaf, E.; Giese, M.; Heudorfer, B.; Stahl, K.; Barthel, R. Physiographic and Climatic Controls on Regional Groundwater Dynamics. Water Resour. Res. 2020, 56, e2019WR026545. [Google Scholar] [CrossRef]
- Barthel, R.; Haaf, E.; Giese, M.; Nygren, M.; Heudorfer, B.; Stahl, K. Similarity-Based Approaches in Hydrogeology: Proposal of a New Concept for Data-Scarce Groundwater Resource Characterization and Prediction. Hydrogeol. J. 2021, 29, 1693–1709. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Plot-1 | Plot-3 |
|---|---|---|
| Lat. | 56.4640 | 56.7146 |
| Lon. | 23.0078 | 23.7426 |
| Yearly mean temperature * | 7.2 °C | 7.1 °C |
| Warmest month, mean temperature * | July, 17.4 °C | July, 17.0 °C |
| Coldest month, mean temperature * | February, −2.7 °C | February, −2.7 °C |
| Yearly mean precipitation * | 580.5 mm/year | 651.1 mm/year |
| Wettest month, mean precipitation * | July, 77.1 mm/moth | July, 82.1 mm/month |
| Driest month, mean precipitation * | March, 29.6 mm/moth | March, 33.8 mm/month |
| Elevation | 91.75 m a.s.l. | 5.22 m a.s.l. |
| Dominant tree species | Alnus glutinosa | Alnus glutinosa |
| Tree height ** | 20 m | 20 m |
| Soil Composition (% Dry Weight) | Van Genuchten–Mualem Parameters | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Site | Depth (cm) | Model Layer No. | Org. Mater | Sand | Silt | Clay | Qr (cm3 cm−3) | Qs (cm3 cm−3) | Alpha (cm−1) | n | Ks (cm day−1) | l |
| Plot-1 | 0.02–0.07 | 1 | 73% | 0.244 | 0.899 | 0.224 | 1.405 | 1898.8 | −0.797 | |||
| Plot-1 | 0.25–0.30 | 2 | 8.6% | 15.6 | 60.2 | 24.2 | 0.250 | 0.507 | 0.3377 | 1.149 | 424.7 | −1.474 |
| Plot-1 | 0.63–0.68 | 3 | 3.1% | 27.9 | 46.6 | 25.5 | 0.290 | 0.41 | 0.0200 | 2.000 | 1.08 | −0.797 |
| Plot-3 | 0.02–0.07 | 1 | 18.5% | 68.8 | 31.2 | 0.0 | 0.026 | 0.817 | 0.0251 | 1.380 | 731.1 | 5.317 |
| Plot-3 | 0.12–0.17 | 2 | 18.5% | 68.8 | 31.2 | 0.0 | 0.075 | 0.656 | 0.0328 | 1.305 | 172.1 | 1.718 |
| Plot-3 | 0.61–0.65 | 3 | 0.9% | 94.5 | 5.5 | 0.0 | 0.029 | 0.375 | 0.0200 | 1.835 | 6.08 | 0.391 |
| Depth (cm) | Boreal | Temperate Deciduous | Temperate Coniferous |
|---|---|---|---|
| 0 | 1.00 | 1.00 | 1.00 |
| 25 | 0.23 | 0.43 | 0.60 |
| 50 | 0.053 | 0.19 | 0.36 |
| 100 | 0.0028 | 0.035 | 0.13 |
| 200 | 0 | 0.0012 | 0.018 |
| Site | RMSE * | (DoY) | |||
|---|---|---|---|---|---|
| Plot-1 | 0.36 | 2.5 | 260 | 0.99 | 129 |
| Plot-3 | 0.21 | 1 | 565 | 1.53 | 147 |
| Site | SeepIn (cm day−1) | Par | kLAI | ||
|---|---|---|---|---|---|
| 0.4 | 0.5 | 0.7 | |||
| Plot-1 | 0 | h_240 cm | 250 | 280 | 320 |
| Plot-1 | 0.01 | h_240 cm | 200 | 240 | 270 |
| Plot-1 | 0.03 | h_240 cm | 74 | 170 | 210 |
| Plot-1 | 0.05 | h_240 cm | 29 | 18 | 160 |
| Plot-3 | 0 | h_240 cm | 170 | 280 | NA |
| Plot-3 | 0.01 | h_240 cm | 120 | 230 | 300 |
| Plot-3 | 0.03 | h_240 cm | 31 | 160 | 230 |
| Plot-3 | 0.05 | h_240 cm | 42 | 79 | 200 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalvāns, A.; Dauškane, I. Hydrological Coupling and Decoupling of Hydric Hemiboreal Forest Sites Inferred from Soil Water Models and Tree-Ring Chronology. Forests 2023, 14, 1734. https://doi.org/10.3390/f14091734
Kalvāns A, Dauškane I. Hydrological Coupling and Decoupling of Hydric Hemiboreal Forest Sites Inferred from Soil Water Models and Tree-Ring Chronology. Forests. 2023; 14(9):1734. https://doi.org/10.3390/f14091734
Chicago/Turabian StyleKalvāns, Andis, and Iluta Dauškane. 2023. "Hydrological Coupling and Decoupling of Hydric Hemiboreal Forest Sites Inferred from Soil Water Models and Tree-Ring Chronology" Forests 14, no. 9: 1734. https://doi.org/10.3390/f14091734
APA StyleKalvāns, A., & Dauškane, I. (2023). Hydrological Coupling and Decoupling of Hydric Hemiboreal Forest Sites Inferred from Soil Water Models and Tree-Ring Chronology. Forests, 14(9), 1734. https://doi.org/10.3390/f14091734