

Multivariate Analysis of the Community Composition of Tidal Freshwater Forests on the Altamaha River, Georgia


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Field Sampling
2.3. Forest Community Analysis Overview
2.3.1. Plot-Level Vegetation Composition
2.3.2. Community Analysis
3. Results
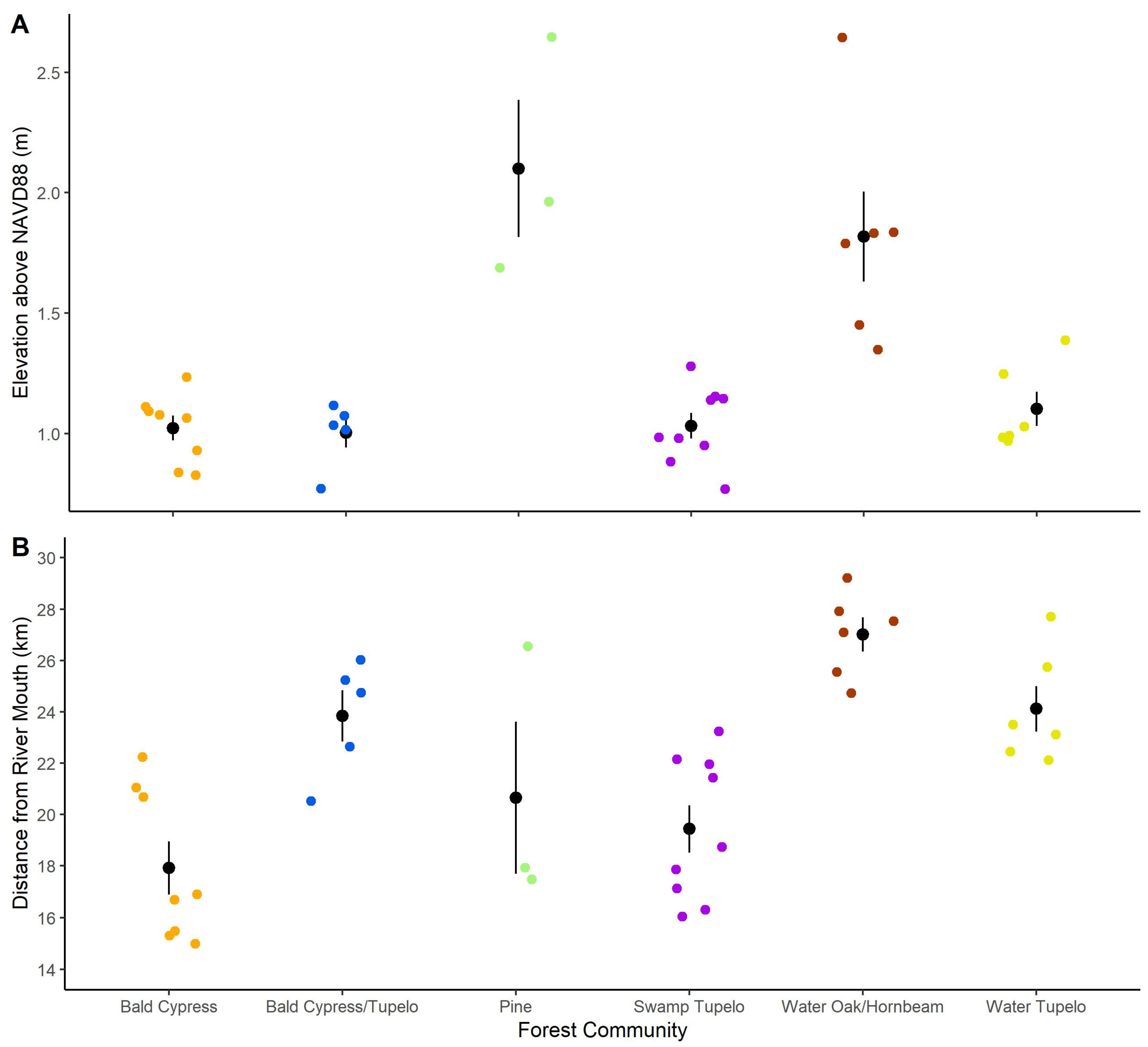
3.1. Plot-Level Species Composition
3.2. Clustering Results
3.3. Community Descriptions
- 1.
- Oak/Hornbeam
- 2.
- Water Tupelo
- 3.
- Bald Cypress/Tupelo
- 4.
- Pine
- 5.
- Swamp Tupelo
- 6.
- Bald Cypress
4. Discussion
4.1. Communities and Ecological Gradients
4.2. Implications and Future Work
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Doyle, T.W.; O'Neil, C.P.; Melder, M.P.V.; From, A.S.; Palta, M.M. Tidal Freshwater Swamps of the Southeastern United States: Effects of Land Use, Hurricanes, Sea-Level Rise, and Climate Change. In Ecology of Tidal Freshwater Forested Wetlands of the Southeastern United States; Conner, W.H., Doyle, T.W., Krauss, K.W., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 1–28. [Google Scholar]
- Grieger, R.; Capon, S.J.; Hadwen, W.L.; Mackey, B. Between a bog and a hard place: A global review of climate change effects on coastal freshwater wetlands. Clim. Change 2020, 163, 161–179. [Google Scholar] [CrossRef]
- Ury, E.A.; Yang, X.; Wright, J.P.; Bernhardt, E.S. Rapid deforestation of a coastal landscape driven by sea-level rise and extreme events. Ecol. Appl. 2021, 31, e02339. [Google Scholar] [CrossRef]
- Chen, Y.; Kirwan, M.L. A phenology-and trend-based approach for accurate mapping of sea-level driven coastal forest retreat. Remote Sens. Environ. 2022, 281, 113229. [Google Scholar] [CrossRef]
- Schieder, N.W.; Walters, D.C.; Kirwan, M.L. Massive upland to wetland conversion compensated for historical marsh loss in Chesapeake Bay, USA. Estuaries Coasts 2018, 41, 940–951. [Google Scholar] [CrossRef]
- Gedan, K.B.; Epanchin-Niell, R.; Qi, M. Rapid land cover change in a submerging coastal county. Wetlands 2020, 40, 1717–1728. [Google Scholar] [CrossRef]
- Anderson, C.J.; Lockaby, B.G.; Click, N. Changes in wetland forest structure, basal growth, and composition across a tidal gradient. Am. Midl. Nat. 2013, 170, 1–13. [Google Scholar] [CrossRef]
- Barendregt, A.; Swarth, C.W. Tidal Freshwater Wetlands: Variation and Changes. Estuaries Coasts 2013, 36, 445–456. [Google Scholar] [CrossRef]
- Duberstein, J.; Kitchens, W. Community composition of select areas of tidal freshwater forest along the Savannah River. In Ecology of Tidal Freshwater Forested Wetlands of the Southeastern United States; Springer: Berlin/Heidelberg, Germany, 2007; pp. 321–348. [Google Scholar]
- Smart, L.S.; Taillie, P.J.; Poulter, B.; Vukomanovic, J.; Singh, K.K.; Swenson, J.J.; Mitasova, H.; Smith, J.W.; Meentemeyer, R.K. Aboveground carbon loss associated with the spread of ghost forests as sea levels rise. Environ. Res. Lett. 2020, 15, 104028. [Google Scholar] [CrossRef]
- Craft, C. Tidal freshwater forest accretion does not keep pace with sea level rise. Glob. Change Biol. 2012, 18, 3615–3623. [Google Scholar] [CrossRef]
- Duberstein, J.A.; Conner, W.H.; Krauss, K.W. Woody vegetation communities of tidal freshwater swamps in South Carolina, Georgia and Florida (US) with comparisons to similar systems in the US and South America. J. Veg. Sci. 2014, 25, 848–862. [Google Scholar] [CrossRef]
- Huylenbroeck, L.; Laslier, M.; Dufour, S.; Georges, B.; Lejeune, P.; Michez, A. Using remote sensing to characterize riparian vegetation: A review of available tools and perspectives for managers. J. Environ. Manag. 2020, 267, 110652. [Google Scholar] [CrossRef]
- Shaffer, G.P.; Wood, W.B.; Hoeppner, S.S.; Perkins, T.E.; Zoller, J.; Kandalepas, D. Degradation of baldcypress–water tupelo swamp to marsh and open water in southeastern Louisiana, USA: An irreversible trajectory? J. Coast. Res. 2009, 54, 152–165. [Google Scholar] [CrossRef]
- Mcleod, E.; Chmura, G.L.; Bouillon, S.; Salm, R.; Björk, M.; Duarte, C.M.; Lovelock, C.E.; Schlesinger, W.H.; Silliman, B.R. A blueprint for blue carbon: Toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Front. Ecol. Environ. 2011, 9, 552–560. [Google Scholar] [CrossRef]
- Stevenson, D.J.; Chandler, H.C. The herpetofauna of conservation lands along the Altamaha River, Georgia. Southeast. Nat. 2017, 16, 261–282. [Google Scholar] [CrossRef]
- Alexander, C.; Hladik, C. High-Resolution Mapping of Vegetation, Elevation, Salinity and Bathymetry to Advance Coastal Habitat Management in Georgia; University of Georgia: Athens, GA, USA, 2015. [Google Scholar]
- Higinbotham, C.B.; Alber, M.; Chalmers, A.G. Analysis of tidal marsh vegetation patterns in two Georgia estuaries using aerial photography and GIS. Estuaries 2004, 27, 670–683. [Google Scholar] [CrossRef]
- White, S.N.; Alber, M. Drought-associated shifts in Spartina alterniflora and S. cynosuroides in the Altamaha River estuary. Wetlands 2009, 29, 215–224. [Google Scholar] [CrossRef]
- Stahl, M.; Widney, S.; Craft, C. Tidal freshwater forests: Sentinels for climate change. Ecol. Eng. 2018, 116, 104–109. [Google Scholar] [CrossRef]
- Carr, J.; Guntenspergen, G.; Kirwan, M. Modeling Marsh-Forest Boundary Transgression in Response to Storms and Sea-Level Rise. Geophys. Res. Lett. 2020, 47, e2020GL088998. [Google Scholar] [CrossRef]
- Craft, C.; Clough, J.; Ehman, J.; Joye, S.; Park, R.; Pennings, S.; Guo, H.; Machmuller, M. Forecasting the effects of accelerated sea-level rise on tidal marsh ecosystem services. Front. Ecol. Environ. 2009, 7, 73–78. [Google Scholar] [CrossRef]
- Zinnert, J.C.; Nippert, J.B.; Rudgers, J.A.; Pennings, S.C.; González, G.; Alber, M.; Baer, S.G.; Blair, J.M.; Burd, A.; Collins, S.L. State changes: Insights from the US Long Term Ecological Research Network. Ecosphere 2021, 12, e03433. [Google Scholar] [CrossRef]
- Jayakaran, A.; Williams, T.; Conner, W.; Hitchcock, D.; Song, B.; Chow, A.; Smith, E. Monitoring water quality changes in a forested freshwater wetland threatened by salinity. In Proceedings of the 2014 South Carolina Water Resources Conference, Columbia, SC, USA, 15–16 October 2014. [Google Scholar]
- Conner, W.; Whitmire, S.; Duberstein, J.; Stalter, R.; Baden, J. Changes within a South Carolina coastal wetland forest in the face of rising sea level. Forests 2022, 13, 414. [Google Scholar] [CrossRef]
- NOAA Tides and Currents. Champney Island, South Altamaha River, GA-Station ID: 8676012. Available online: https://tidesandcurrents.noaa.gov/stationhome.html?id=8676012 (accessed on 2 January 2023).
- Alber, M.; Alexander, C.; Craft, C.; Hladik, C.M.; Medieros, P.M.; Pennings, S.C. RAPID: 2017 Hurricane Irma: How Do the Effects of Pulse Disturbance Caused by Hurricanes Vary with Abiotic Conditions, Disturbance History and Proximity to a Transition Zone; Final Report Submitted to NSF; NSF: Alexandria, VA, USA, 2019; p. 15. [Google Scholar]
- Conner, W.H.; Krauss, K.W.; Doyle, T.W. Ecology of Tidal Freshwater Forests in Coastal Deltaic Louisiana and Northeastern South Carolina. In Ecology of Tidal Freshwater Forested Wetlands of the Southeastern United States; Conner, W.H., Doyle, T.W., Krauss, K.W., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 223–253. [Google Scholar]
- Wharton, C.H.; Kitchens, W.M.; Pendleton, E.C.; Sipe, T.W. The Ecology of Bottomland Hardwood Swamps of the Southeast: A Community Profile; U.S. Forest Service, U.S. Department of Agriculture: Washington, DC, USA, 1982. [Google Scholar]
- Carpenter, J.; Meehan, C. Georgia Coastal Ecosystems LTER Domain; University of Georgia: Athens, GA, USA, 2009. [Google Scholar]
- Anderson, C.J.; Lockaby, B.G. Soils and biogeochemistry of tidal freshwater forested wetlands. In Ecology of Tidal Freshwater Forested Wetlands of the Southeastern United States; Springer: Berlin/Heidelberg, Germany, 2007; pp. 65–88. [Google Scholar]
- NRCS Web Soil Survey. Natural Resources Conservation Service; United States Department of Agriculture: Washington, DC, USA, 2021. [Google Scholar]
- Odum, W.E.; Smith, T.J., III; Hoover, J.K.; McIvor, C.C. The Ecology of Tidal Freshwater Marshes of the United States East Coast: A Community Profile; U.S. Fish and Wildlife Service: Washington, DC, USA, 1984; p. 177. [Google Scholar]
- Tully, K.; Gedan, K.; Epanchin-Niell, R.; Strong, A.; Bernhardt, E.S.; BenDor, T.; Mitchell, M.; Kominoski, J.; Jordan, T.E.; Neubauer, S.C. The invisible flood: The chemistry, ecology, and social implications of coastal saltwater intrusion. BioScience 2019, 69, 368–378. [Google Scholar] [CrossRef]
- Herbert, E.R.; Boon, P.; Burgin, A.J.; Neubauer, S.C.; Franklin, R.B.; Ardón, M.; Hopfensperger, K.N.; Lamers, L.P.; Gell, P. A global perspective on wetland salinization: Ecological consequences of a growing threat to freshwater wetlands. Ecosphere 2015, 6, 1–43. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 2 January 2023).
- Curtis, J.T.; McIntosh, R.P. An Upland Forest Continuum in the Prarie-Forest Border Region of Wisconsin. Ecology 1951, 32, 20. [Google Scholar] [CrossRef]
- Tobe, J.D. Florida Wetland Plants: An Identification Manual; University Press of Florida: Gainesville, FL, USA, 1998. [Google Scholar]
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002; Volume 28. [Google Scholar]
- Legendre, P.; De Cáceres, M. Beta diversity as the variance of community data: Dissimilarity coefficients and partitioning. Ecol. Lett. 2013, 16, 951–963. [Google Scholar] [CrossRef]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 192, 271–280. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O'Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. Version 2.6-4. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 12 July 2022).
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. Version 1.0.7. 2020. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 11 September 2022).
- Milligan, G.W.; Cooper, M.C. An Examination of Procedures for Determining the Number of Clusters in a Data Set. Psychometrika 1985, 50, 159–179. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- De Câceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- Conner, W.H.; Song, B.; Williams, T.M.; Vernon, J.T. Long-term tree productivity of a South Carolina coastal plain forest across a hydrology gradient. J. Plant Ecol. 2011, 4, 67–76. [Google Scholar] [CrossRef]
- Light, H.M.; Darst, M.R.; Lewis, L.J.; Howell, D.A. Hydrology, Vegetation, and Soils of Riverine and Tidal Floodplain Forests of the Lower Suwannee River, Florida, and Potential Impacts of Flow Reductions; US Geological Survey: Reston, VA, USA, 2002; ISSN 2330-7102. [Google Scholar]
- Krauss, K.W.; Duberstein, J.A.; Doyle, T.W.; Conner, W.H.; Day, R.H.; Inabinette, L.W.; Whitbeck, J.L. Site condition, structure, and growth of baldcypress along tidal/non-tidal salinity gradients. Wetlands 2009, 29, 505–519. [Google Scholar] [CrossRef]
- Hladik, C.; Alber, M. Accuracy assessment and correction of a LIDAR-derived salt marsh digital elevation model. Remote Sens. Environ. 2012, 121, 224–235. [Google Scholar] [CrossRef]
- Dunn, O.J. Multiple comparisons using rank sums. Technometrics 1964, 6, 241–252. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Flitcroft, R.; Clinton, P.; Christiansen, K. Adding to the toolbox for tidal-inundation mapping in estuarine areas. J. Coast. Conserv. 2018, 22, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.J.; Lockaby, B.G. Forested wetland communities as indicators of tidal influence along the Apalachicola River, Florida, USA. Wetlands 2011, 31, 895–906. [Google Scholar] [CrossRef]
- Celik, S.; Anderson, C.J.; Kalin, L.; Rezaeianzadeh, M. Long-term salinity, hydrology, and forested wetlands along a tidal freshwater gradient. Estuaries Coasts 2021, 44, 1816–1830. [Google Scholar] [CrossRef]
- Cormier, N.; Krauss, K.W.; Conner, W.H. Periodicity in stem growth and litterfall in tidal freshwater forested wetlands: Influence of salinity and drought on nitrogen recycling. Estuaries Coasts 2013, 36, 533–546. [Google Scholar] [CrossRef]
- Ensign, S.H.; Hupp, C.R.; Noe, G.B.; Krauss, K.W.; Stagg, C.L. Sediment accretion in tidal freshwater forests and oligohaline marshes of the Waccamaw and Savannah Rivers, USA. Estuaries Coasts 2014, 37, 1107–1119. [Google Scholar] [CrossRef]
- Liu, X.; Conner, W.H.; Song, B.; Jayakaran, A.D. Forest composition and growth in a freshwater forested wetland community across a salinity gradient in South Carolina, USA. For. Ecol. Manag. 2017, 389, 211–219. [Google Scholar] [CrossRef]
- Beane, N.R. Baldcypress (Taxodium Distichum) at the Wallisville Lake Project: A Review of Applicable Literature and Management Considerations; US Army Corps of Engineers Engineer Research and Development Center: Vicksburg, MI, USA, 2020. [Google Scholar]
- Day, R.H.; Williams, T.M.; Swarzenski, C.M. Hydrology of tidal freshwater forested wetlands of the southeastern United States. In Ecology of Tidal Freshwater Forested Wetlands of the Southeastern United States; Springer: Berlin/Heidelberg, Germany, 2007; pp. 29–63. [Google Scholar]
- Tiner, R.W. Tidal Wetlands Primer: An Introduction to Their Ecology. Natural History, Status, and Conservation; University of Massachusetts Press: Amherst, MA, USA, 2013. [Google Scholar]
- Sharitz, R.R.; Lee, L.C. Limits on Regeneration Processes in Southeastern Riverine Wetlands. In Proceedings of the Riparian Ecosystems and Their Management: Reconciling Conflicting Uses, Tucson, AZ, USA, 16–18 April 1985. [Google Scholar]
- Williams, K.; Ewel, K.C.; Stumpf, R.P.; Putz, F.E.; Workman, T.W. Sea-level rise and coastal forest retreat on the west coast of Florida, USA. Ecology 1999, 80, 2045–2063. [Google Scholar] [CrossRef]
- Desantis, L.R.; Bhotika, S.; Williams, K.; Putz, F.E. Sea-level rise and drought interactions accelerate forest decline on the Gulf Coast of Florida, USA. Glob. Change Biol. 2007, 13, 2349–2360. [Google Scholar] [CrossRef]
- Conner, W.H.; Doyle, T.W.; Krauss, K.W.; Kroes, D.E.; Hupp, C.R.; Noe, G.B. Sediment, nutrient, and vegetation trends along the tidal, forested Pocomoke River, Maryland. In Ecology of Tidal Freshwater Forested Wetlands of the Southeastern United States; Springer: Berlin/Heidelberg, Germany, 2007; pp. 113–137. [Google Scholar]
- Middleton, B.A. Differences in impacts of Hurricane Sandy on freshwater swamps on the Delmarva Peninsula, Mid-Atlantic Coast, USA. Ecol. Eng. 2016, 87, 62–70. [Google Scholar] [CrossRef]
- Middleton, B.A.; Souter, N.J. Functional integrity of freshwater forested wetlands, hydrologic alteration, and climate change. Ecosyst. Health Sustain. 2016, 2, e01200. [Google Scholar] [CrossRef]
- Noe, G.B.; Bourg, N.A.; Krauss, K.W.; Duberstein, J.A.; Hupp, C.R. Watershed and estuarine controls both influence plant Community and tree growth changes in tidal freshwater forested wetlands along two US Mid-Atlantic Rivers. Forests 2021, 12, 1182. [Google Scholar] [CrossRef]
- Larson, J.S.; Bedinger, M.; Bryan, C.F.; Brown, S.; Huffman, R.T.; Miller, E.L.; Rhodes, D.G.; Touchet, B.A. Transition from wetlands to uplands in southeastern bottomland hardwood forests. In Developments in Agricultural and Managed Forest Ecology; Elsevier: Amsterdam, The Netherlands, 1981; Volume 11, pp. 225–273. [Google Scholar]
- Duberstein, J.A.; Krauss, K.W.; Baldwin, M.J.; Allen, S.T.; Conner, W.H.; Salter Jr, J.S.; Miloshis, M. Small gradients in salinity have large effects on stand water use in freshwater wetland forests. For. Ecol. Manag. 2020, 473, 118308. [Google Scholar] [CrossRef]
- Krauss, K.W.; Chambers, J.L.; Creech, D. Selection for Salt Tolerance in Tidal Freshwater Swamp Species: Advances Using Baldcypress as a Model for Restoration. In Ecology of Tidal Freshwater Forested Wetlands of the Southeastern United States; Conner, W.H., Doyle, T.W., Krauss, K.W., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 385–410. [Google Scholar]
- Craft, C.; Indiana University, Bloomington IN, USA. Personal Communication, 2024.
- Conner, W.H.; Doyle, T.W.; Krauss, K.W.; Hackney, C.T.; Avery, G.B.; Leonard, L.A.; Posey, M.; Alphin, T. Biological, chemical, and physical characteristics of tidal freshwater swamp forests of the Lower Cape Fear River/Estuary, North Carolina. In Ecology of Tidal Freshwater Forested Wetlands of the Southeastern United States; Springer: Berlin/Heidelberg, Germany, 2007; pp. 183–221. [Google Scholar]
- Fagherazzi, S.; Anisfeld, S.C.; Blum, L.K.; Long, E.V.; Feagin, R.A.; Fernandes, A.; Kearney, W.S.; Williams, K. Sea level rise and the dynamics of the marsh-upland boundary. Front. Environ. Sci. 2019, 7, 25. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Gosselink, J.G. Wetlands, 2nd ed.; Van Nostrand Reinhold: New York, NY, USA, 1993. [Google Scholar]
- Raabe, E.A.; Stumpf, R.P. Expansion of tidal marsh in response to sea-level rise: Gulf Coast of Florida, USA. Estuaries Coasts 2015, 39, 145–157. [Google Scholar] [CrossRef]
- Conner, W.H.; Askew, G.R. Impact of saltwater flooding on red maple, redbay, and Chinese tallow seedlings. Castanea 1993, 58, 214–219. [Google Scholar]
- McDowell, N.G.; Ball, M.; Bond-Lamberty, B.; Kirwan, M.L.; Krauss, K.W.; Megonigal, J.P.; Mencuccini, M.; Ward, N.D.; Weintraub, M.N.; Bailey, V. Processes and mechanisms of coastal woody-plant mortality. Glob. Change Biol. 2022, 28, 5881–5900. [Google Scholar] [CrossRef]
- Middleton, B.A.; David, J.L. Trends in vegetation and height of the topographic surface in a tidal freshwater swamp experiencing rooting zone saltwater intrusion. Ecol. Indic. 2022, 145, 109637. [Google Scholar] [CrossRef]
- Di Iorio, D. Continuous Salinity, Temperature and Depth Measurements from Moored Hydrographic Data Loggers Deployed at GCE11_Hydro (Altamaha River Near Lewis Creek, Georgia) from 07-Oct-2017 through 31-Dec-2017; Georgia Coastal Ecosystems LTER Data Portal: Athens, GA, USA, 2018. [Google Scholar] [CrossRef]
- Di Iorio, D. Long-Term Hydrographic Mooring Data from the Georgia Coastal Ecosystems LTER Salinity Monitoring Program-Primary 30 Minute Observational Data; Georgia Coastal Ecosystems LTER Project, Ed.; University of Georgia, Long Term Ecological Research Network: Athens, GA, USA, 2016. [Google Scholar] [CrossRef]
- Marton, J.M.; Herbert, E.R.; Craft, C.B. Effects of salinity on denitrification and greenhouse gas production from laboratory-incubated tidal forest soils. Wetlands 2012, 32, 347–357. [Google Scholar] [CrossRef]
- Neubauer, S.C.; Craft, C.B. Global change and tidal freshwater wetlands: Scenarios and impacts. In Tidal Freshwater Wetlands; Springer: Berlin/Heidelberg, Germany, 2009; pp. 253–266. [Google Scholar]
- Seyfried, G.S.; Chow, A.T.; O'Halloran, T.L. Salinization, inundation and tree mortality interact to affect greenhouse gas emissions from stressed coastal forests. Soil Biol. Biochem. 2023, 184, 109101. [Google Scholar] [CrossRef]
- Pierfelice, K.N.; Graeme Lockaby, B.; Krauss, K.W.; Conner, W.H.; Noe, G.B.; Ricker, M.C. Salinity influences on aboveground and belowground net primary productivity in tidal wetlands. J. Hydrol. Eng. 2017, 22, D5015002. [Google Scholar] [CrossRef]
- Martinez, M.; Ardon, M. Drivers of greenhouse gas emissions from standing dead trees in ghost forests. Biogeochemistry 2021, 154, 471–488. [Google Scholar] [CrossRef]
- Martinez, M.; Ardón, M.; Carmichael, M.J. Identifying Sources and Oxidation of Methane in Standing Dead Trees in Freshwater Forested Wetlands. Front. Environ. Sci. 2022, 9, 701. [Google Scholar] [CrossRef]
- Ensign, S.H.; Noe, G.B. Tidal extension and sea-level rise: Recommendations for a research agenda. Front. Ecol. Environ. 2018, 16, 37–43. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Common Name | Community | |||||
|---|---|---|---|---|---|---|---|
| Oak/Hornbeam | Water Tupelo | Bald Cypress/Tupelo | Pine | Swamp Tupelo | Bald Cypress | ||
| Acer rubrum | Red maple | 0.04 | 0.01 | 0.02 | 0.00 | 0.03 | 0.02 |
| Alnus serrulata | Hazel alder | 0.00 | 0.00 | 0.01 | 0.00 | 0.00 | 0.00 |
| Betula nigra | River birch | 0.01 | 0.00 | 0.01 | 0.00 | 0.00 | 0.00 |
| Carya aquatica | Water hickory | 0.02 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Carpinus caroliniana | American hornbeam | 0.04 | 0.00 | 0.01 | 0.00 | 0.00 | 0.00 |
| Fraxinus spp. | Ash | 0.02 | 0.07 | 0.06 | 0.00 | 0.20 | 0.07 |
| Liquidambar styraciflua | Sweetgum | 0.10 | 0.07 | 0.06 | 0.07 | 0.07 | 0.02 |
| Nyssa aquatica | Water tupelo | 0.01 | 0.56 | 0.23 | 0.00 | 0.03 | 0.00 |
| Nyssa biflora | Swamp tupelo | 0.09 | 0.04 | 0.21 | 0.00 | 0.58 | 0.15 |
| Persea palustris | Swamp bay | 0.00 | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 |
| Pinus spp. | Pine | 0.08 | 0.00 | 0.00 | 0.93 | 0.00 | 0.00 |
| Planera aquatica | Water elm | 0.01 | 0.00 | 0.01 | 0.00 | 0.00 | 0.00 |
| Quercus laurifolia/nigra | Water oak/laurel oak | 0.50 | 0.12 | 0.02 | 0.00 | 0.02 | 0.00 |
| Taxodium distichum | Bald cypress | 0.06 | 0.11 | 0.37 | 0.00 | 0.10 | 0.75 |
| Ulmus americana | American elm | 0.01 | 0.00 | 0.01 | 0.00 | 0.00 | 0.00 |
| Number of Plots | 6 | 6 | 5 | 3 | 9 | 8 | |
| Average Elevation (m above NAVD88) | 1.82 | 1.10 | 1.00 | 2.10 | 1.03 | 1.02 | |
| Basal area (m2·ha−1) | 41.1 | 72.9 | 68.1 | 42.0 | 47.2 | 37.2 | |
| Density (stems·ha−1) | 1140 | 1113 | 1256 | 1033 | 1362 | 668 | |
| Mean Canopy Coverage (%) | 95.9 | 97.1 | 92.9 | 97.5 | 83.0 | 85.5 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costomiris, G.; Hladik, C.M.; Craft, C. Multivariate Analysis of the Community Composition of Tidal Freshwater Forests on the Altamaha River, Georgia. Forests 2024, 15, 200. https://doi.org/10.3390/f15010200
Costomiris G, Hladik CM, Craft C. Multivariate Analysis of the Community Composition of Tidal Freshwater Forests on the Altamaha River, Georgia. Forests. 2024; 15(1):200. https://doi.org/10.3390/f15010200
Chicago/Turabian StyleCostomiris, Galen, Christine M. Hladik, and Christopher Craft. 2024. "Multivariate Analysis of the Community Composition of Tidal Freshwater Forests on the Altamaha River, Georgia" Forests 15, no. 1: 200. https://doi.org/10.3390/f15010200
APA StyleCostomiris, G., Hladik, C. M., & Craft, C. (2024). Multivariate Analysis of the Community Composition of Tidal Freshwater Forests on the Altamaha River, Georgia. Forests, 15(1), 200. https://doi.org/10.3390/f15010200