
Do Mature Quercus aliena Primarily Use Deep Soil Water?


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
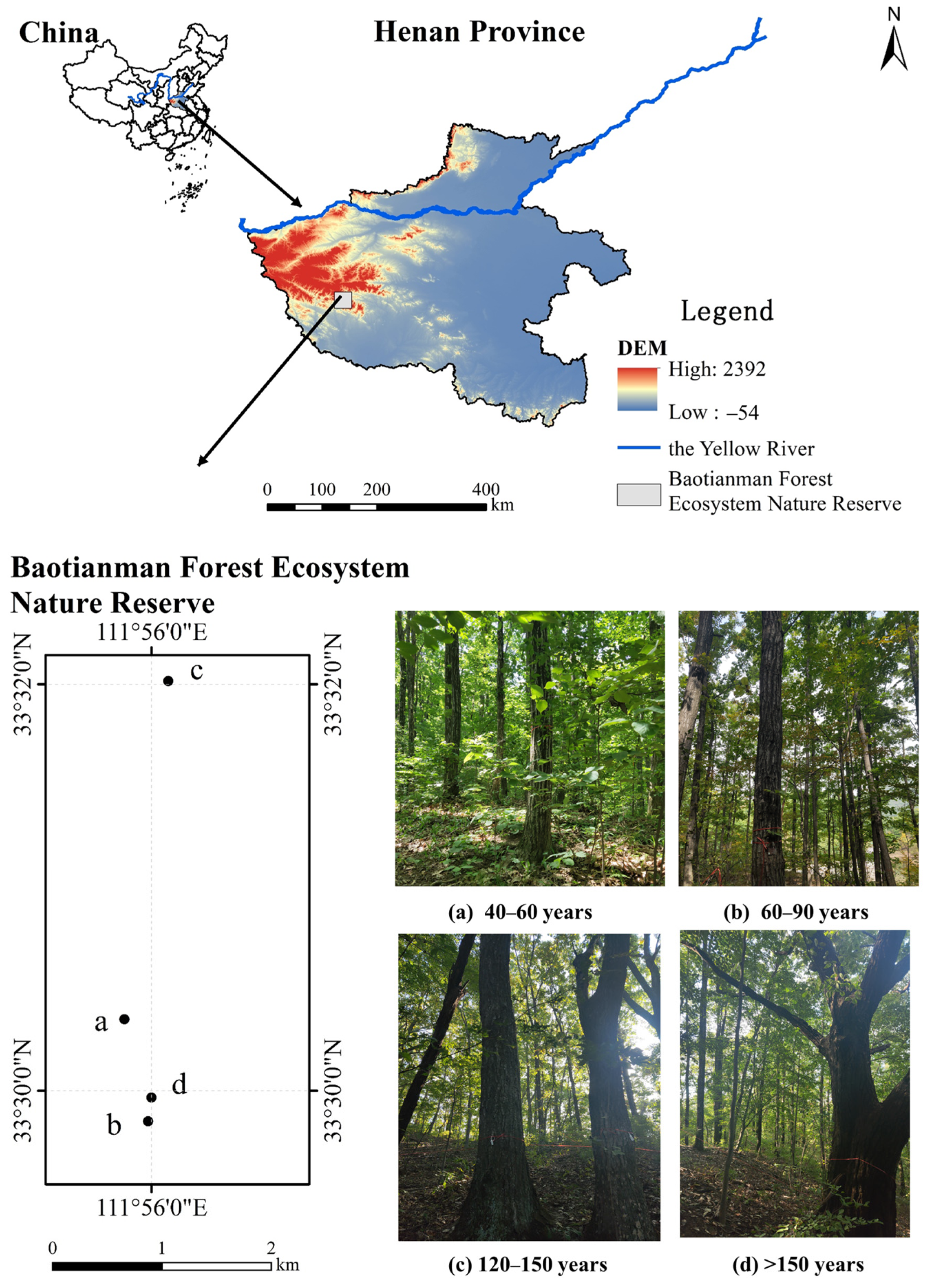
2.1. Study Area
2.2. Samples Collection
2.3. Sample Pre-Treatment and Isotope Analysis
2.4. Determination of Water Uptake Pattern
2.5. Determination of Vegetation Biomass and Soil Physical Properties
2.6. Statistical Analysis
3. Results
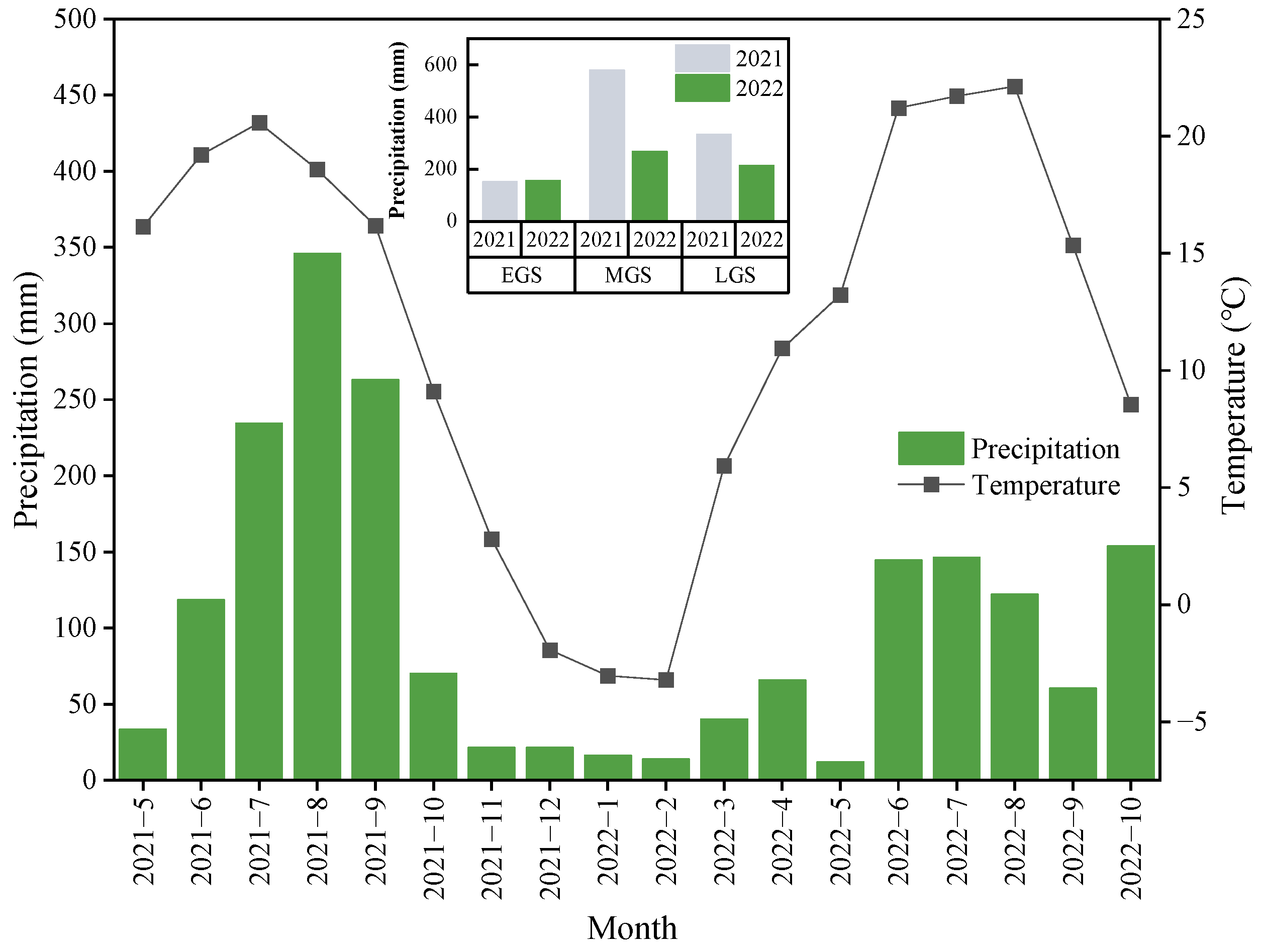
3.1. Environmental Conditions
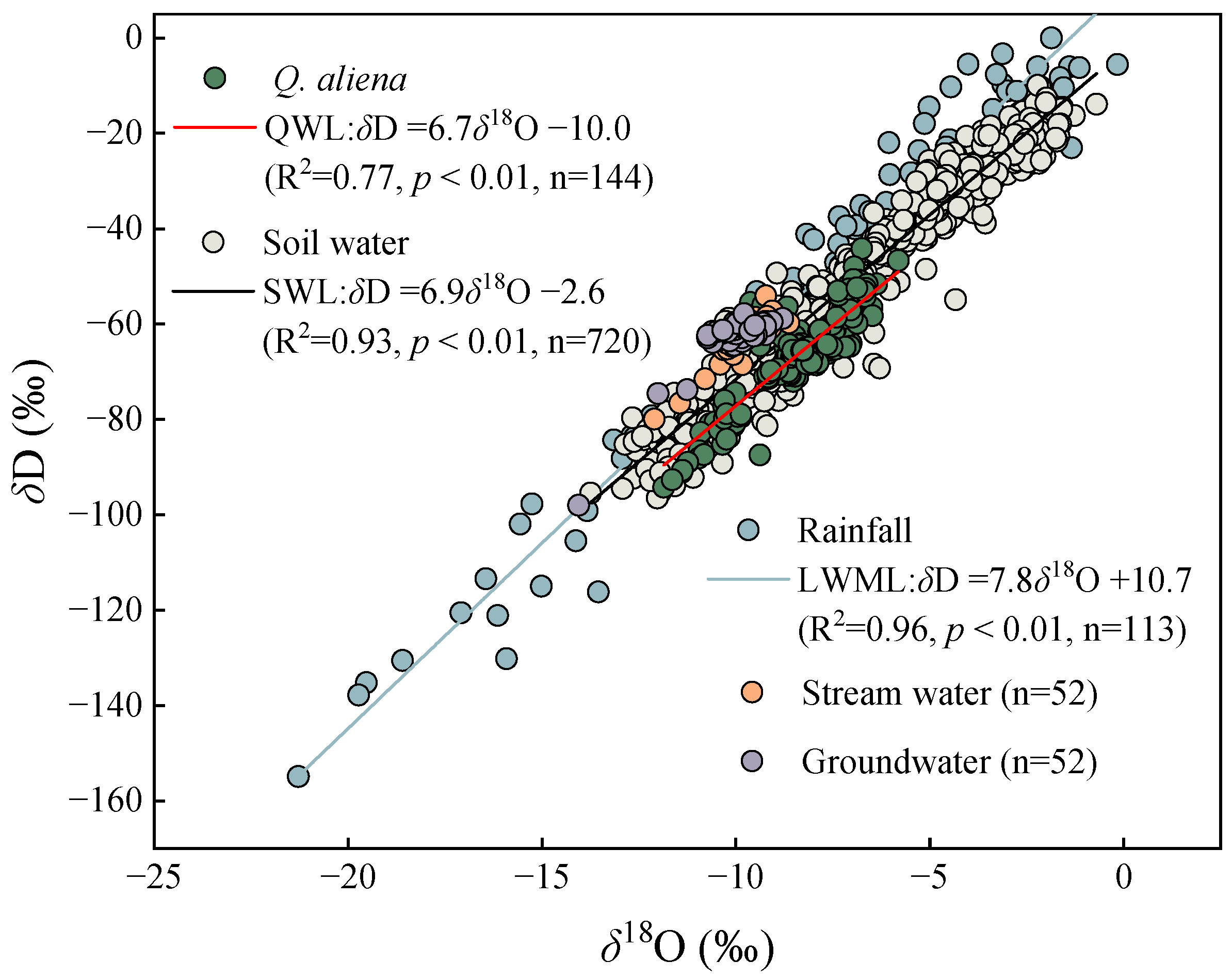
3.2. Relationship between Tree Xylem Water and Potential Water Sources
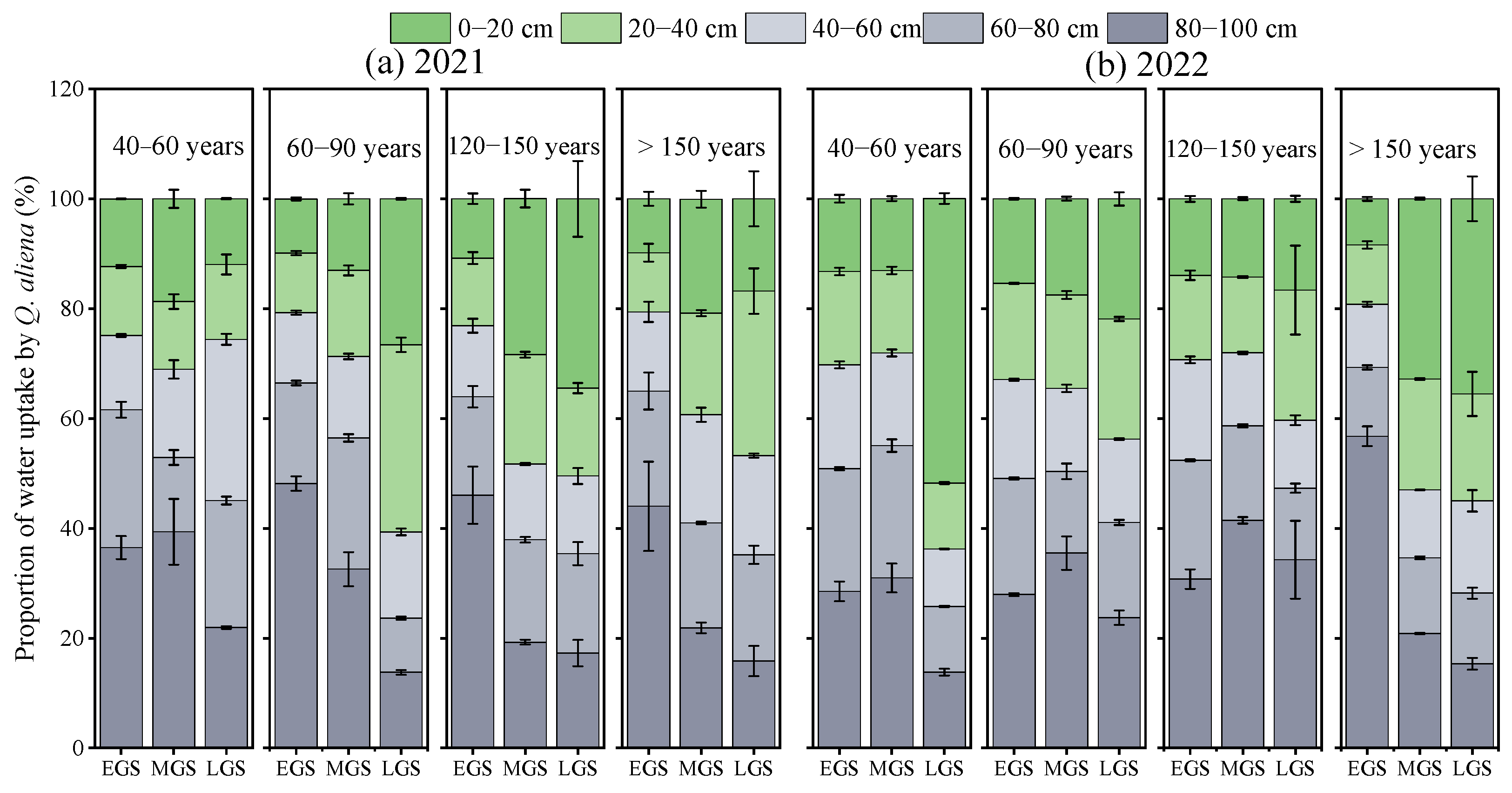
3.3. Water Uptake Patterns of Q. aliena Trees
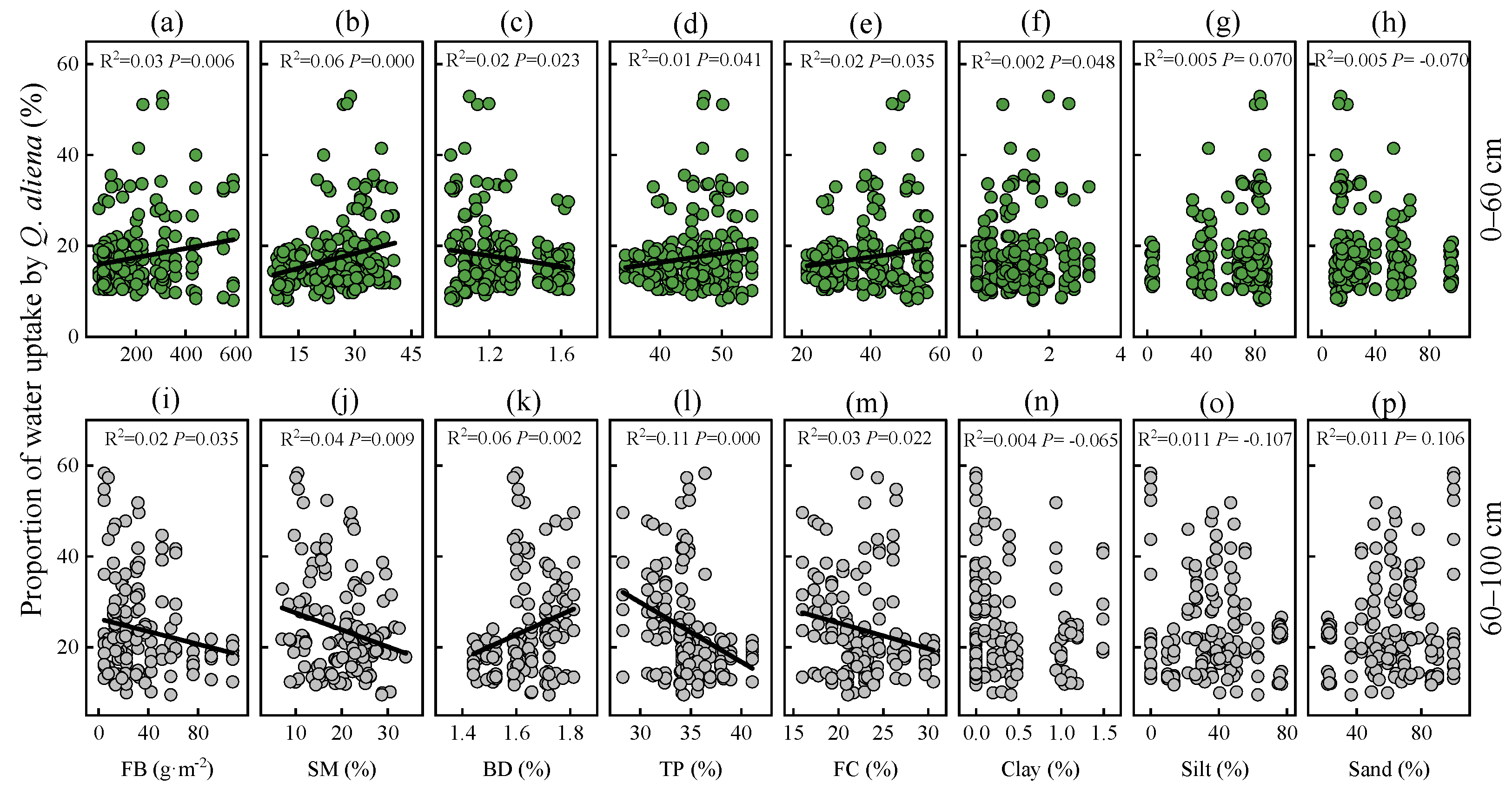
3.4. Relationship between Water Uptake Pattern of Q. aliena Trees and Plant or Soil Properties in the Growing Season
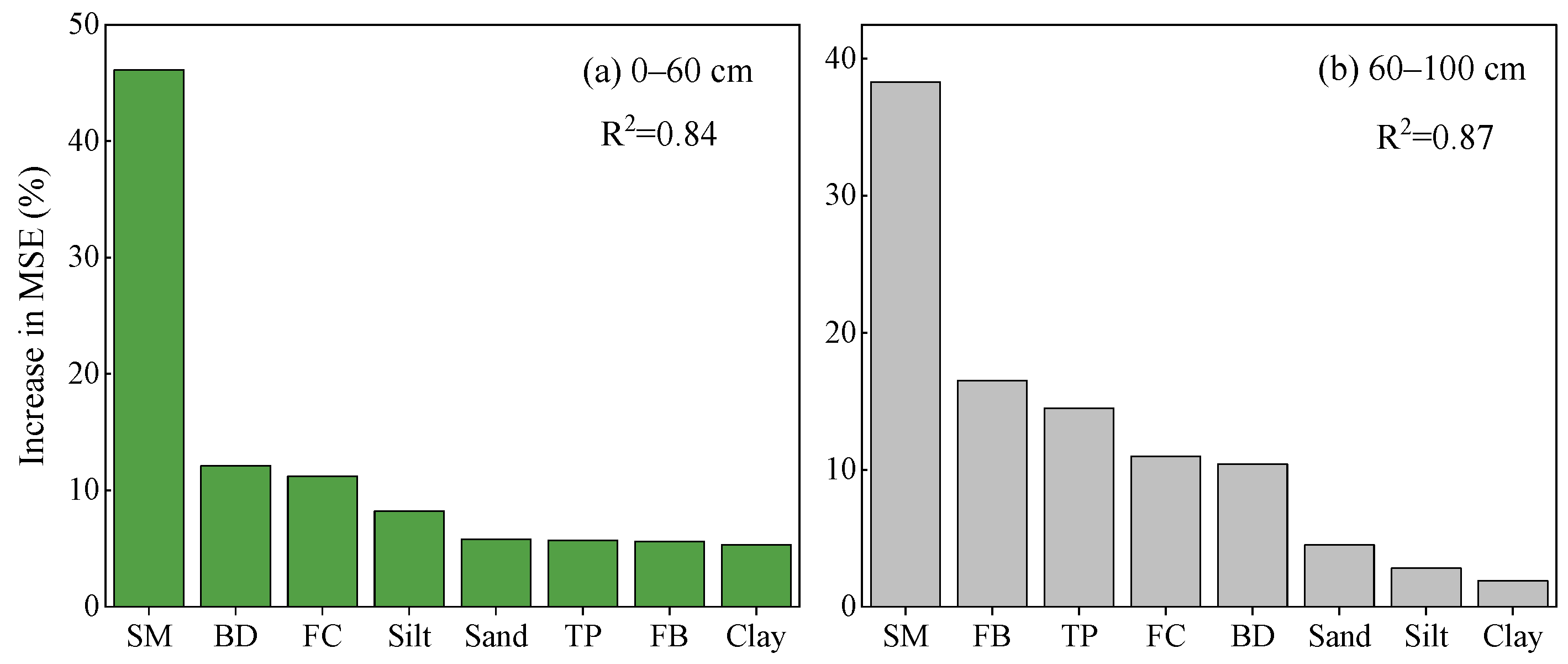
3.5. Dominant Driver of Water Use Pattern
4. Discussion
4.1. Water Use Strategies of Mature Q. aliena Trees
4.2. Key Drivers of Water Use Strategies for Mature Q. aliena Trees
4.3. Implications and Uncertainties
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pappas, C.; Peters, R.L.; Fonti, P. Linking Variability of Tree Water Use and Growth with Species Resilience to Environmental Changes. Ecography 2020, 43, 1386–1399. [Google Scholar] [CrossRef]
- Stocker, B.D.; Tumber-Dávila, S.J.; Konings, A.G.; Anderson, M.C.; Hain, C.; Jackson, R.B. Global Patterns of Water Storage in the Rooting Zones of Vegetation. Nat. Geosci. 2023, 16, 250–256. [Google Scholar] [CrossRef]
- Bayala, J.; Prieto, I. Water Acquisition, Sharing and Redistribution by Roots: Applications to Agroforestry Systems. Plant Soil 2020, 453, 17–28. [Google Scholar] [CrossRef]
- Zheng, P.; Wang, D.; Jia, G.; Yu, X.; Liu, Z.; Wang, Y.; Zhang, Y. Variation in Water Supply Leads to Different Responses of Tree Growth to Warming. For. Ecosyst. 2022, 9, 100003. [Google Scholar] [CrossRef]
- Ouyang, L.; Wu, J.; Zhao, P.; Zhu, L.; Ni, G. Stand Age Rather than Soil Moisture Gradient Mainly Regulates the Compromise between Plant Growth and Water Use of Eucalyptus Urophylla in Hilly South China. Land Degrad. Dev. 2021, 32, 2423–2436. [Google Scholar] [CrossRef]
- McDowell, N.G.; Phillips, N.; Lunch, C.; Bond, B.J.; Ryan, M.G. An Investigation of Hydraulic Limitation and Compensation in Large, Old Douglas-Fir Trees. Tree Physiol. 2002, 22, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Delzon, S.; Loustau, D. Age-Related Decline in Stand Water Use: Sap Flow and Transpiration in a Pine Forest Chronosequence. Agric. For. Meteorol. 2005, 129, 105–119. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A Global Overview of Drought and Heat-Induced Tree Mortality Reveals Emerging Climate Change Risks for Forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Meir, P.; Mencuccini, M.; Dewar, R.C. Drought-related Tree Mortality: Addressing the Gaps in Understanding and Prediction. New Phytol. 2015, 207, 28–33. [Google Scholar] [CrossRef]
- Johnson, D.M.; Domec, J.-C.; Carter Berry, Z.; Schwantes, A.M.; McCulloh, K.A.; Woodruff, D.R.; Wayne Polley, H.; Wortemann, R.; Swenson, J.J.; Scott Mackay, D. Co-occurring Woody Species Have Diverse Hydraulic Strategies and Mortality Rates during an Extreme Drought. Plant Cell Environ. 2018, 41, 576–588. [Google Scholar] [CrossRef]
- Reich, P.B.; Sendall, K.M.; Stefanski, A.; Rich, R.L.; Hobbie, S.E.; Montgomery, R.A. Effects of Climate Warming on Photosynthesis in Boreal Tree Species Depend on Soil Moisture. Nature 2018, 562, 263–267. [Google Scholar] [CrossRef]
- Dawson, T.E.; Ehleringer, J.R. Streamside Trees That Do Not Use Stream Water. Nature 1991, 350, 335–337. [Google Scholar] [CrossRef]
- Huo, G.; Zhao, X.; Gao, X.; Wang, S.; Pan, Y. Seasonal Water Use Patterns of Rainfed Jujube Trees in Stands of Different Ages under Semiarid Plantations in China. Agric. Ecosyst. Environ. 2018, 265, 392–401. [Google Scholar] [CrossRef]
- Martin-Benito, D.; Pederson, N.; Férriz, M.; Gea-Izquierdo, G. Old Forests and Old Carbon: A Case Study on the Stand Dynamics and Longevity of Aboveground Carbon. Sci. Total Environ. 2021, 765, 142737. [Google Scholar] [CrossRef]
- Pyttel, P.L.; Fischer, U.F.; Suchomel, C.; Gärtner, S.M.; Bauhus, J. The Effect of Harvesting on Stump Mortality and Re-Sprouting in Aged Oak Coppice Forests. For. Ecol. Manag. 2013, 289, 18–27. [Google Scholar] [CrossRef]
- Song, L.; Zhu, J.; Li, M.; Zhang, J. Water Use Patterns of Pinus Sylvestris Var. Mongolica Trees of Different Ages in a Semiarid Sandy Lands of Northeast China. Environ. Exp. Bot. 2016, 129, 94–107. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, Y.; Qiu, K.; López-Vicente, M.; Shen, W.; Wu, G.-L. Soil-Water Deficit in Deep Soil Layers Results from the Planted Forest in a Semi-Arid Sandy Land: Implications for Sustainable Agroforestry Water Management. Agric. Water Manag. 2021, 254, 106985. [Google Scholar] [CrossRef]
- Wang, S.; An, J.; Zhao, X.; Gao, X.; Wu, P.; Huo, G.; Robinson, B.H. Age-and Climate-Related Water Use Patterns of Apple Trees on China’s Loess Plateau. J. Hydrol. 2020, 582, 124462. [Google Scholar] [CrossRef]
- Song, L.; Yang, B.; Liu, L.-L.; Mo, Y.-X.; Liu, W.-J.; Meng, X.-J.; Lu, H.-Z.; Li, Y.; Zakari, S.; Tan, Z.-H. Spatial-Temporal Differentiations in Water Use of Coexisting Trees from a Subtropical Evergreen Broadleaved Forest in Southwest China. Agric. For. Meteorol. 2022, 316, 108862. [Google Scholar] [CrossRef]
- Kuster, T.M.; Arend, M.; Günthardt-Goerg, M.S.; Schulin, R. Root Growth of Different Oak Provenances in Two Soils under Drought Stress and Air Warming Conditions. Plant Soil 2013, 369, 61–71. [Google Scholar] [CrossRef]
- Skubel, R.; Arain, M.A.; Peichl, M.; Brodeur, J.J.; Khomik, M.; Thorne, R.; Trant, J.; Kula, M. Age Effects on the Water-Use Efficiency and Water-Use Dynamics of Temperate Pine Plantation Forests. Hydrol. Process. 2015, 29, 4100–4113. [Google Scholar] [CrossRef]
- Langs, L.E.; Petrone, R.M.; Pomeroy, J.W. A δ18O and δ2H Stable Water Isotope Analysis of Subalpine Forest Water Sources under Seasonal and Hydrological Stress in the Canadian Rocky Mountains. Hydrol. Process. 2020, 34, 5642–5658. [Google Scholar] [CrossRef]
- Liu, S.; Chen, Y.; Chen, Y.; Friedman, J.M.; Hati, J.H.A.; Fang, G. Use of 2 H and 18 O Stable Isotopes to Investigate Water Sources for Different Ages of Populus Euphratica along the Lower Heihe River. Ecol. Res. 2015, 30, 581–587. [Google Scholar] [CrossRef]
- Süßel, F.; Brüggemann, W. Tree Water Relations of Mature Oaks in Southwest Germany under Extreme Drought Stress in Summer 2018. Plant Stress 2021, 1, 100010. [Google Scholar] [CrossRef]
- Zhang, X.; Yu, P.; Wang, D.; Xu, Z. Density-and Age-Dependent Influences of Droughts and Intrinsic Water Use Efficiency on Growth in Temperate Plantations. Agric. For. Meteorol. 2022, 325, 109134. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, M.; Qu, D.; Wang, S.; Argiriou, A.A.; Wang, J.; Yang, Y. Water Use Characteristics of Different Pioneer Shrubs at Different Ages in Western Chinese Loess Plateau: Evidence from δ2H Offset Correction. J. Arid Land 2022, 14, 653–672. [Google Scholar] [CrossRef]
- Song, L.; Zhu, J.; Li, M.; Zhang, J.; Li, D. Water Use Strategies of Natural Pinus Sylvestris Var. Mongolica Trees of Different Ages in Hulunbuir Sandy Land of Inner Mongolia, China, Based on Stable Isotope Analysis. Trees 2018, 32, 1001–1011. [Google Scholar] [CrossRef]
- Pei, Y.; Huang, L.; Zhang, Y.; Pan, Y. Water Use Pattern and Transpiration of Mongolian Pine Plantations in Relation to Stand Age on Northern Loess Plateau of China. Agric. For. Meteorol. 2023, 330, 109320. [Google Scholar] [CrossRef]
- Bargués Tobella, A.; Hasselquist, N.; Bazié, H.; Nyberg, G.; Laudon, H.; Bayala, J.; Ilstedt, U. Strategies Trees Use to Overcome Seasonal Water Limitation in an Agroforestry System in Semiarid West Africa. Ecohydrology 2017, 10, e1808. [Google Scholar] [CrossRef]
- Nadezhdina, N.; Ferreira, M.I.; Silva, R.; Pacheco, C.A. Seasonal Variation of Water Uptake of a Quercus Suber Tree in Central Portugal. Plant Soil 2008, 305, 105–119. [Google Scholar] [CrossRef]
- Ellsworth, P.Z.; Sternberg, L.S.L. Seasonal Water Use by Deciduous and Evergreen Woody Species in a Scrub Community Is Based on Water Availability and Root Distribution. Ecohydrology 2015, 8, 538–551. [Google Scholar] [CrossRef]
- Baldocchi, D.D.; Xu, L.; Kiang, N. How Plant Functional-Type, Weather, Seasonal Drought, and Soil Physical Properties Alter Water and Energy Fluxes of an Oak–Grass Savanna and an Annual Grassland. Agric. For. Meteorol. 2004, 123, 13–39. [Google Scholar] [CrossRef]
- Cavalcanti, R.Q.; Rolim, M.M.; de Lima, R.P.; Tavares, U.E.; Pedrosa, E.M.R.; Cherubin, M.R. Soil Physical Changes Induced by Sugarcane Cultivation in the Atlantic Forest Biome, Northeastern Brazil. Geoderma 2020, 370, 114353. [Google Scholar] [CrossRef]
- Voltas, J.; Lucabaugh, D.; Chambel, M.R.; Ferrio, J.P. Intraspecific Variation in the Use of Water Sources by the Circum-Mediterranean Conifer Pinus halepensis. New Phytol. 2015, 208, 1031–1041. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, Q.; Zhang, B.; Gao, D.; Wang, T.; Xu, W.; Ren, R.; Wang, S. Contrasting Water-Use Patterns of Chinese Fir among Different Plantation Types in a Subtropical Region of China. Front. Plant Sci. 2022, 13, 946508. [Google Scholar] [CrossRef]
- Wang, D.; Fu, B.; Zhao, W.; Hu, H.; Wang, Y. Multifractal Characteristics of Soil Particle Size Distribution under Different Land-Use Types on the Loess Plateau, China. CATENA 2008, 72, 29–36. [Google Scholar] [CrossRef]
- Zhang, B.; Xu, Q.; Gao, D.; Jiang, C.; Liu, F.; Jiang, J.; Wang, T. Altered Water Uptake Patterns of Populus Deltoides in Mixed Riparian Forest Stands. Sci. Total Environ. 2020, 706, 135956. [Google Scholar] [CrossRef]
- Imada, S.; Yamanaka, N.; Tamai, S. Water Table Depth Affects Populus Alba Fine Root Growth and Whole Plant Biomass. Funct. Ecol. 2008, 22, 1018–1026. [Google Scholar] [CrossRef]
- Fan, Y.; Miguez-Macho, G.; Jobbágy, E.G.; Jackson, R.B.; Otero-Casal, C. Hydrologic Regulation of Plant Rooting Depth. Proc. Natl. Acad. Sci. USA 2017, 114, 10572–10577. [Google Scholar] [CrossRef]
- Jia, H.-R.; Chen, Y.; Yuan, Z.-L.; Ye, Y.-Z.; Huang, Q.-C. Effects of Environmental and Spatial Heterogeneity on Tree Community Assembly in Baotianman National Nature Reserve, Henan, China. Pol. J. Ecol. 2015, 63, 175–183. [Google Scholar] [CrossRef]
- Qi, G.; Peng, S.L.; Chen, C.D.; Zhao, G.Q.; Liang, Y.H.; Li, P.; Yang, F.L.; Wen, Z.Z. Biomass Carbon Storage of Three Forests at Different Altitudes in Baotianman Mountain, Central China. Adv. Mater. Res. 2014, 864, 2463–2468. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, S.; Lu, H.; Wan, X. Interaction of Stomatal Behaviour and Vulnerability to Xylem Cavitation Determines the Drought Response of Three Temperate Tree Species. AoB Plants 2019, 11, plz058. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Helliker, B.R.; Tang, X.; Li, F.; Zhou, Y.; Song, X. Stem Water Cryogenic Extraction Biases Estimation in Deuterium Isotope Composition of Plant Source Water. Proc. Natl. Acad. Sci. USA 2020, 117, 33345–33350. [Google Scholar] [CrossRef] [PubMed]
- Lyu, S.; Wang, J.; Song, X.; Wen, X. The Relationship of δD and δ18O in Surface Soil Water and Its Implications for Soil Evaporation along Grass Transects of Tibet, Loess, and Inner Mongolia Plateau. J. Hydrol. 2021, 600, 126533. [Google Scholar] [CrossRef]
- Ehleringer, J.R.; Dawson, T.E. Water Uptake by Plants: Perspectives from Stable Isotope Composition. Plant Cell Environ. 1992, 15, 1073–1082. [Google Scholar] [CrossRef]
- Stock, B.C.; Semmens, B.X. MixSIAR GUI User Manual: Version 1.0. 2013, Version 1.0; Scripps Institution of Oceanography: San Diego, CA, USA, 2018.
- Wang, J.; Fu, B.; Lu, N.; Zhang, L. Seasonal Variation in Water Uptake Patterns of Three Plant Species Based on Stable Isotopes in the Semi-Arid Loess Plateau. Sci. Total Environ. 2017, 609, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Özcan, M.; Gökbulak, F.; Hizal, A. Exclosure Effects on Recovery of Selected Soil Properties in a Mixed Broadleaf Forest Recreation Site. Land Degrad. Dev. 2013, 24, 266–276. [Google Scholar] [CrossRef]
- Cui, H.; Bai, J.; Du, S.; Wang, J.; Keculah, G.N.; Wang, W.; Zhang, G.; Jia, J. Interactive Effects of Groundwater Level and Salinity on Soil Respiration in Coastal Wetlands of a Chinese Delta. Environ. Pollut. 2021, 286, 117400. [Google Scholar] [CrossRef]
- Werner, C.; Meredith, L.K.; Ladd, S.N.; Ingrisch, J.; Kübert, A.; van Haren, J.; Bahn, M.; Bailey, K.; Bamberger, I.; Beyer, M. Ecosystem Fluxes during Drought and Recovery in an Experimental Forest. Science 2021, 374, 1514–1518. [Google Scholar] [CrossRef]
- Grossiord, C.; Sevanto, S.; Dawson, T.E.; Adams, H.D.; Collins, A.D.; Dickman, L.T.; Newman, B.D.; Stockton, E.A.; McDowell, N.G. Warming Combined with More Extreme Precipitation Regimes Modifies the Water Sources Used by Trees. New Phytol. 2017, 213, 584–596. [Google Scholar] [CrossRef]
- Wang, B.; Chen, T.; Li, C.; Xu, G.; Wu, G.; Liu, G. Discrepancy in Growth Resilience to Drought among Different Stand-Aged Forests Declines Going from a Semi-Humid Region to an Arid Region. For. Ecol. Manag. 2022, 511, 120135. [Google Scholar] [CrossRef]
- Manoli, G.; Meijide, A.; Huth, N.; Knohl, A.; Kosugi, Y.; Burlando, P.; Ghazoul, J.; Fatichi, S. Ecohydrological Changes after Tropical Forest Conversion to Oil Palm. Environ. Res. Lett. 2018, 13, 064035. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, Y.; He, M.; Tong, Y.; Zhou, J.; Guo, X.; Liu, J.; Zhang, X. Transference of Robinia Pseudoacacia Water-Use Patterns from Deep to Shallow Soil Layers during the Transition Period between the Dry and Rainy Seasons in a Water-Limited Region. For. Ecol. Manag. 2020, 457, 117727. [Google Scholar] [CrossRef]
- Jagodzinski, A.M.; Ziółkowski, J.; Warnkowska, A.; Prais, H. Tree Age Effects on Fine Root Biomass and Morphology over Chronosequences of Fagus Sylvatica, Quercus Robur and Alnus Glutinosa Stands. PLoS ONE 2016, 11, e0148668. [Google Scholar] [CrossRef]
- Gwak, Y.; Kim, S. Factors Affecting Soil Moisture Spatial Variability for a Humid Forest Hillslope. Hydrol. Process. 2017, 31, 431–445. [Google Scholar] [CrossRef]
- Coners, H.; Leuschner, C. In Situ Measurement of Fine Root Water Absorption in Three Temperate Tree Species—Temporal Variability and Control by Soil and Atmospheric Factors. Basic. Appl. Ecol. 2005, 6, 395–405. [Google Scholar] [CrossRef]
- Volkmann, T.H.; Haberer, K.; Gessler, A.; Weiler, M. High-resolution Isotope Measurements Resolve Rapid Ecohydrological Dynamics at the Soil–Plant Interface. New Phytol. 2016, 210, 839–849. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Sites | Stand Age (Years) | Altitude (m) | Slope (°) | Stand Density (tree/hm2) | BDH (cm) |
|---|---|---|---|---|---|
| 1 | 40–60 | 1386.0 | 17 | 1075 | 23.8 |
| 2 | 90–120 | 1369.6 | 12 | 692 | 27.2 |
| 3 | 120–150 | 1411.2 | 20 | 275 | 51.9 |
| 4 | >150 | 1365.9 | 15 | 62 | 87.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, R.; Zhang, B.; Xu, Q.; Gao, D.; Xu, W.; Zuo, H.; Zhang, Y.; Diao, K. Do Mature Quercus aliena Primarily Use Deep Soil Water? Forests 2024, 15, 402. https://doi.org/10.3390/f15030402
Ren R, Zhang B, Xu Q, Gao D, Xu W, Zuo H, Zhang Y, Diao K. Do Mature Quercus aliena Primarily Use Deep Soil Water? Forests. 2024; 15(3):402. https://doi.org/10.3390/f15030402
Chicago/Turabian StyleRen, Ranran, Beibei Zhang, Qing Xu, Deqiang Gao, Wenbin Xu, Haijun Zuo, Ying Zhang, and Ke Diao. 2024. "Do Mature Quercus aliena Primarily Use Deep Soil Water?" Forests 15, no. 3: 402. https://doi.org/10.3390/f15030402
APA StyleRen, R., Zhang, B., Xu, Q., Gao, D., Xu, W., Zuo, H., Zhang, Y., & Diao, K. (2024). Do Mature Quercus aliena Primarily Use Deep Soil Water? Forests, 15(3), 402. https://doi.org/10.3390/f15030402