
Improving the Simulation Accuracy of the Net Ecosystem Productivity of Subtropical Forests in China: Sensitivity Analysis and Parameter Calibration Based on the BIOME-BGC Model


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Subtropical Forest Flux Data and Meteorological Factors
2.3. Soil and Topographic Data
2.4. BIOME-BGC Model Description

2.5. Sensitivity Analysis
2.5.1. Morris Method
2.5.2. EFAST
2.5.3. Parameter Value Establishment
2.5.4. Sensitivity Analysis Steps
2.6. Model Accuracy Validation
3. Results
3.1. Uncertainty Analysis
3.2. Morris Sensitivity Analysis
3.3. EFAST Sensitivity Analysis
3.4. Comparative Analysis of Morris and EFAST Results
3.5. BIOME-BGC Model Validation
4. Discussion
4.1. Uncertainty Analysis
4.2. Sensitive Parameters
5. Conclusions
- Both the Morris method and EFAST can effectively screen out the important parameters affecting the output of the model. The Morris method has a significant advantage when the sample size is small, the parameters are numerous, and the computational effort is high; however, it is only a qualitative method of parameter sensitivity analysis. The EFAST method allows further quantitative analysis of the contribution of each input parameter and the interaction between parameters to the simulation results; however, its computational efficiency is lower than that of the Morris method. For parameter sensitivity analyses of complex process models, the Morris method is used for qualitative studies and the EFAST method is used for quantitative studies;
- The parameters k, SC:LC, SLA, FRC:LC, Gsmax, MRpern, Ko25, Ract25, Q10Ract, and SCRages (only for the BF) significantly affected the simulated subtropical forest NEP. Priority should be given to these parameters in model parameter optimization and correction to reduce computation and improve model accuracy;
- Compared with the flux observation data, the parameter-optimized BIOME-BGC model significantly improved the simulation ability of the original model for the NEP of subtropical forest ecosystems in China; the average R of the NEP increased by 25.19% and the average NRMSE reduced by 21.74%.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Buotte, P.C.; Law, B.E.; Ripple, W.J.; Berner, L.T. Carbon sequestration and biodiversity co-benefits of preserving forests in the western United States. Ecol. Appl. 2020, 30, e02039. [Google Scholar] [CrossRef]
- Mcveigh, P.; Sottocornola, M.; Foley, N.; Leahy, P.; Kiely, G. Meteorological and functional response partitioning to explain interannual variability of CO2 exchange at an Irish Atlantic blanket bog. Agric. For. Meteorol. 2014, 194, 8–19. [Google Scholar] [CrossRef]
- Mao, F.; Du, H.; Zhou, G.; Zheng, J.; Li, X.; Xu, Y.; Huang, Z.; Yin, S. Simulated net ecosystem productivity of subtropical forests and its response to climate change in Zhejiang Province, China. Sci. Total Environ. 2022, 838, 155993. [Google Scholar] [CrossRef]
- Mitchard, E.T.A. The tropical forest carbon cycle and climate change. Nature 2018, 559, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Fennel, K.; Losch, M.; Schröter, J.; Wenzel, M. Testing a marine ecosystem model: Sensitivity analysis and parameter optimization. J. Mar. Syst. 2001, 28, 45–63. [Google Scholar] [CrossRef]
- Raj, R.; Tol, C.V.D.; Hamm, N.A.S.; Stein, A. Bayesian integration of flux tower data into a process-based simulator for quantifying uncertainty in simulated output. Geosci. Model Dev. 2016, 11, 83–101. [Google Scholar] [CrossRef]
- He, L.H.; Wang, H.Y.; Lei, X.D. Parameter sensitivity of simulating net primary productivity of Larix olgensis forest based on BIOME-BGC model. Chin. J. Appl. Ecol. 2016, 27, 412–420. [Google Scholar]
- Du, L.; Zeng, Y.; Ma, L.; Qiao, C.; Wu, H.; Su, Z.; Bao, G. Effects of anthropogenic revegetation on the water and carbon cycles of a desert steppe ecosystem. Agric. For. Meteorol. 2021, 300, 108339. [Google Scholar] [CrossRef]
- Tian, X.; Yan, M.; Van Der Tol, C.; Li, Z.; Su, Z.; Chen, E.; Li, X.; Li, L.; Wang, X.; Pan, X.; et al. Modeling forest above-ground biomass dynamics using multi-source data and incorporated models: A case study over the qilian mountains. Agric. For. Meteorol. 2017, 246, 1–14. [Google Scholar] [CrossRef]
- Keller, E.D.; Baisden, W.T.; Timar, L.; Mullan, B.; Clark, A. Grassland production under global change scenarios for New Zealand pastoral agriculture. Geosci. Model Dev. 2014, 7, 2359–2391. [Google Scholar] [CrossRef]
- Saltelli, A.; Annoni, P. How to avoid a perfunctory sensitivity analysis. Environ. Modell. Softw. 2010, 25, 1508–1517. [Google Scholar] [CrossRef]
- Cukier, R.; Levine, H.; Shuler, K. Nonlinear sensitivity analysis of multiparameter model systems. J. Comput. Phys. 1978, 26, 1–42. [Google Scholar] [CrossRef]
- Morris, M.D. Factorial Sampling Plans for Preliminary Computational Experiments. Technometrics 1991, 33, 161–174. [Google Scholar] [CrossRef]
- Sobol, I.M. Global sensitivity indices for nonlinear mathematical models and their Monte Carlo estimates. Math. Comput. Simul. 2001, 55, 271–280. [Google Scholar] [CrossRef]
- Saltelli, A.; Tarantola, S.; Chan, K.P.S. A Quantitative Model-Independent Method for Global Sensitivity Analysis of Model Output. Technometrics 1999, 41, 39–56. [Google Scholar] [CrossRef]
- Wang, S.; Flipo, N.; Romary, T. Time-dependent global sensitivity analysis of the C-RIVE biogeochemical model in contrasted hydrological and trophic contexts. Water Res. 2018, 144, 341–355. [Google Scholar] [CrossRef] [PubMed]
- Dejonge, K.C.; Ascough Ii, J.C.; Ahmadi, M.; Andales, A.A.; Arabi, M. Global sensitivity and uncertainty analysis of a dynamic agroecosystem model under different irrigation treatments. Ecol. Model. 2012, 231, 113–125. [Google Scholar] [CrossRef]
- Peng, X.; Adamowski, J.; Inam, A.; Alizadeh, M.R.; Albano, R. Development of a behaviour-pattern based global sensitivity analysis procedure for coupled socioeconomic and environmental models. J. Hydrol. 2020, 585, 124745. [Google Scholar] [CrossRef]
- Li, Z.; Jin, X.; Liu, H.; Xu, X.; Wang, J. Global sensitivity analysis of wheat grain yield and quality and the related process variables from the DSSAT-CERES model based on the extended Fourier Amplitude Sensitivity Test method. J. Integr. Agric. 2019, 18, 1547–1561. [Google Scholar] [CrossRef]
- Gilardelli, C.; Confalonieri, R.; Cappelli, G.A.; Bellocchi, G. Sensitivity of WOFOST-based modelling solutions to crop parameters under climate change. Ecol. Model. 2018, 368, 1–14. [Google Scholar] [CrossRef]
- Li, L.; Wei, S.; Lian, J.; Cao, H. Distributional regularity of species diversity in plant community at different latitudes in subtropics. Chin. J. Ecol. 2020, 40, 1249–1257. [Google Scholar]
- Yuen, J.Q.; Fung, T.; Ziegler, A.D. Carbon stocks in bamboo ecosystems worldwide: Estimates and uncertainties. For. Ecol. Manag. 2017, 393, 113–138. [Google Scholar] [CrossRef]
- Yan, M.; Tian, X.; Li, Z.; Chen, E.; Wang, X.; Han, Z.; Sun, H. Simulation of Forest Carbon Fluxes Using Model Incorporation and Data Assimilation. Remote Sens. 2016, 8, 567. [Google Scholar] [CrossRef]
- Tatarinov, F.A.; Cienciala, E. Application of BIOME-BGC model to managed forests: 1. Sensitivity analysis. For. Ecol. Manag. 2006, 237, 267–279. [Google Scholar] [CrossRef]
- Raj, R.; Hamm, N.a.S.; Van Der Tol, C.; Stein, A. Variance-based sensitivity analysis of BIOME-BGC for gross and net primary production. Ecol. Model. 2014, 292, 26–36. [Google Scholar] [CrossRef]
- Liu, J.; Wu, Z.; Yang, S.; Yang, C. Sensitivity Analysis of Biome-BGC for Gross Primary Production of a Rubber Plantation Ecosystem: A Case Study of Hainan Island, China. Int. J. Environ. Res. Public Health 2022, 19, 14068. [Google Scholar] [CrossRef]
- Kumar, M.; Raghubanshi, A.S. Sensitivity analysis of BIOME-BGC model for dry tropical forests of Vindhyan highlands, India. Remote Sens. Spat. Inf. Sci. 2012, 38, 129–133. [Google Scholar] [CrossRef]
- Mao, F.; Du, H.; Zhou, G.; Li, X.; Xu, X.; Li, P.; Sun, S. Coupled LAI assimilation and BEPS model for analyzing the spatiotemporal pattern and heterogeneity of carbon fluxes of the bamboo forest in Zhejiang Province, China. Agric. For. Meteorol. 2017, 242, 96–108. [Google Scholar] [CrossRef]
- Papale, D.; Reichstein, M.; Aubinet, M.; Canfora, E.; Bernhofer, C.; Kutsch, W.; Longdoz, B.; Rambal, S.; Valentini, R.; Vesala, T. Towards a standardized processing of Net Ecosystem Exchange measured with eddy covariance technique: Algorithms and uncertainty estimation. Biogeosciences 2006, 3, 571–583. [Google Scholar] [CrossRef]
- Lasslop, G.; Reichstein, M.; Papale, D.; Richardson, A.D.; Arneth, A.; Barr, A.; Stoy, P.; Wohlfahrt, G. Separation of net ecosystem exchange into assimilation and respiration using a light response curve approach: Critical issues and global evaluation. Glob. Change Biol. 2010, 16, 187–208. [Google Scholar] [CrossRef]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; Granier, A. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: Review and improved algorithm. Glob. Change Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- Fujisada, H.; Bailey, G.B.; Kelly, G.G.; Hara, S.; Abrams, M.J. Aster dem performance. IEEE Trans. Geosci. Remote Sens. 2005, 43, 2707–2714. [Google Scholar] [CrossRef]
- Running, S.W.; Coughlan, J.C. A general model of forest ecosystem processes for regional applications I. Hydrologic balance, canopy gas exchange and primary production processes. Ecol. Model. 1988, 42, 125–154. [Google Scholar] [CrossRef]
- Running, S.W.; Hunt, E.R., Jr. Generalization of a Forest Ecosystem Process Model for other Biomes, BIOME-BCG, and an Application for Global-Scale Models; Academic Press: Cambridge, MA, USA, 1993. [Google Scholar]
- Du, H.; Mao, F.; Zhou, G.; Li, X.; Xu, X.; Ge, H.; Cui, L.; Liu, Y.; Zhu, D.E.; Li, Y. Estimating and Analyzing the Spatiotemporal Pattern of Aboveground Carbon in Bamboo Forest by Combining Remote Sensing Data and Improved BIOME-BGC Model. IEEE J. Sel. Top. App. Earth Observ. Remote Sens. 2018, 11, 2282–2295. [Google Scholar] [CrossRef]
- Mao, F.; Li, P.; Zhou, G.; Du, H.; Xu, X.; Shi, Y.; Mo, L.; Zhou, Y.; Tu, G. Development of the BIOME-BGC model for the simulation of managed Moso bamboo forest ecosystems. J. Environ. Manag. 2016, 172, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Thornton, P.E.; Rosenbloom, N.A. Ecosystem model spin-up: Estimating steady state conditions in a coupled terrestrial carbon and nitrogen cycle model. Ecol. Model. 2005, 189, 25–48. [Google Scholar] [CrossRef]
- Thornton, P.E.; Law, B.E.; Gholz, H.L.; Clark, K.L.; Falge, E.; Ellsworth, D.S.; Goldstein, A.H.; Monson, R.K.; Hollinger, D.; Falk, M. Modeling and measuring the effects of disturbance history and climate on carbon and water budgets in evergreen needleleaf forests. Agric. For. Meteorol. 2002, 113, 185–222. [Google Scholar] [CrossRef]
- Campolongo, F.; Cariboni, J.; Saltelli, A. An effective screening design for sensitivity analysis of large models. Environ. Modell. Softw. 2007, 22, 1509–1518. [Google Scholar] [CrossRef]
- Saltelli, A.; Ratto, M.; Andres, T.; Campolongo, F.; Cariboni, J.; Gatelli, D.; Saisana, M.; Tarantola, S. Global Sensitivity Analysis: The Primer; John Wiley & Sons: Chichester, UK, 2008. [Google Scholar]
- White, M.A.; Thornton, P.E.; Running, S.W.; Nemani, R.R. Parameterization and Sensitivity Analysis of the BIOME–BGC Terrestrial Ecosystem Model: Net Primary Production Controls. Earth Interact. 2000, 4, 1–85. [Google Scholar] [CrossRef]
- Waring, R.H.; Running, S.W. Forest Ecosystems: Analysis at Multiple Scales; Elsevier: Amsterdam, The Netherlands, 2010. [Google Scholar]
- Woodrow, I.E.; Berry, J.A. Enzymatic Regulation of Photosynthetic CO2, Fixation in C3 Plants. Annu. Rev. Plant Physiol. Plant Molec. Biol. 1988, 39, 533–594. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, X.; Ouyang, S.; Xu, K.; Hawkins, B.A.; Sun, O.J. A test of BIOME-BGC with dendrochronology for forests along the altitudinal gradient of Mt. Changbai in northeast China. J. Plant Ecol. 2016, 123, 439–452. [Google Scholar] [CrossRef]
- Zhou, C.-H.; Hao, Z.-Q.; He, H.-S.; Zhou, D.-H. Sensitivity of parameters in net primary productivity model of broadleaf-Korean pine mixed forest. Chin. J. Appl. Ecol. 2008, 19, 929–935. [Google Scholar]
- Gao, Q.; Peng, S.; Zhao, P.; Zeng, X.; Cai, X.; Yu, M.; Shen, W.; Liu, Y. Explanation of vegetation succession in subtropical southern China based on ecophysiological characteristics of plant species. Tree Physiol. 2003, 23, 641–648. [Google Scholar] [CrossRef]
- Lu, W.; Fan, W.Y.; Tian, T. Parameter optimization of BEPS model based on the flux data of the temperate deciduous broad-leaved forest in Northeast China. Chin. J. Appl. Ecol. 2016, 27, 1353–1358. [Google Scholar]
- Liu, G.; Fan, S.; Su, W.; Xiao, F.; Huang, Y. Distribution Characteristics and Coupling Relationship of Organic Carbon and Total Nitrogen in Phyllostachys pubescens Forests with Different Operations and Management Modes. J. Soil Water Conserv. 2010, 24, 218–222. [Google Scholar]
- Zhang, J.; Zhang, D.; Jian, Z.; Zhou, H.; Zhao, Y.; Wei, D. Litter decomposition and the degradation of recalcitrant components in Pinus massoniana plantations with various canopy densities. J. For. Res. 2019, 30, 1395–1405. [Google Scholar] [CrossRef]
- Liu, Z.; Fei, B. Characteristics of Moso Bamboo with Chemical Pretreatment; IntechOpen: London, UK, 2013. [Google Scholar]
- Holling, C.S.; Schindler, D.W.; Walker, B.H.; Roughgarden, J. Biodiversity in the Functioning of Ecosystems: An Ecological Synthesis; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Jin, A. High Profit Management and Participatory Development of Phyllostachys Pubescens Plantation in Zhejiang and Fujian Mountainous Areas. Ph.D. Dissertation, Nanjing Forestry University, Nanjing, China, 2004. [Google Scholar]
- Ren, H.; Zhang, L.; Yan, M.; Tian, X.; Zheng, X. Sensitivity analysis of Biome-BGCMuSo for gross and net primary productivity of typical forests in China. For. Ecosyst. 2022, 9, 100011. [Google Scholar] [CrossRef]
- Li, X.; Sun, J. Testing parameter sensitivities and uncertainty analysis of Biome-BGC model in simulating carbon and water fluxes in broadleaved-Korean pine forests. Chin. J. Plant Ecol. 2018, 42, 1131–1144. [Google Scholar] [CrossRef]
- Kang, M. Energy Partitioning and Modelling of Carbon and Water fluxes of a Poplar Plantation Ecosystem in Northern China. Ph.D. Dissertation, Beijing Forestry University, Beijing, China, 2016. [Google Scholar]
- Zheng, L.; Song, S.; Yuan, X.; Dong, J.; Li, L. Simulation of water and carbon fluxes in a broad-leaved Korean pine forest in Changbai Mountains based on Biome-BGC model and Ensemble Kalman Filter method. Chin. J. Ecol. 2017, 36, 1752–1760. [Google Scholar]
- Jung, M.; Vetter, M.; Herold, M.; Churkina, G.; Reichstein, M.; Zaehle, S.; Ciais, P.; Viovy, N.; Bondeau, A.; Chen, Y. Uncertainties of modeling gross primary productivity over Europe: A systematic study on the effects of using different drivers and terrestrial biosphere models. Glob. Biogeochem. Cycle. 2007, 21. [Google Scholar] [CrossRef]
- Eastaugh, C.S.; Pötzelsberger, E.; Hasenauer, H. Assessing the impacts of climate change and nitrogen deposition on Norway spruce (Picea abies L. Karst) growth in Austria with BIOME-BGC. Tree Physiol. 2011, 31, 262–274. [Google Scholar]
- Ma, H.; Ma, C.; Li, X.; Yuan, W.; Liu, Z.; Zhu, G. Sensitivity and uncertainty analyses of flux-based ecosystem model towards improvement of forest GPP simulation. Sustainability 2020, 12, 2584. [Google Scholar] [CrossRef]
- Saltelli, A.; Annoni, P.; Azzini, I.; Campolongo, F.; Ratto, M.; Tarantola, S. Variance based sensitivity analysis of model output. Design and estimator for the total sensitivity index. Comput. Phys. Commun. 2010, 181, 259–270. [Google Scholar] [CrossRef]
- Feng, K.; Lu, Z.; Yang, C. Enhanced Morris method for global sensitivity analysis: Good proxy of Sobol’ index. Struct. Multidiscip. Optim. 2018, 59, 373–387. [Google Scholar] [CrossRef]
- Sobol, I.; Kucherenko, S. Derivative based global sensitivity measures. Procedia. Soc. Behav. Sci. 2010, 2, 7745–7746. [Google Scholar] [CrossRef]
- Ciffroy, P.; Benedetti, M. A comprehensive probabilistic approach for integrating natural variability and parametric uncertainty in the prediction of trace metals speciation in surface waters. Environ. Pollut. 2018, 242, 1087–1097. [Google Scholar] [CrossRef]
- Xue, H.; Tian, X.; Wang, B.; Sun, S.; Cao, T. Comparison of global sensitivity analysis techniques based on a process-based model CROBAS. Chin. J. Appl. Ecol. 2021, 32, 134–144. [Google Scholar]
- Srinet, R.; Nandy, S.; Patel, N.R.; Padalia, H.; Watham, T.; Singh, S.K.; Chauhan, P. Simulation of forest carbon fluxes by integrating remote sensing data into biome-BGC model. Ecol. Model. 2023, 475, 110185. [Google Scholar] [CrossRef]
- Ryan, M.G. Effects of climate change on plant respiration. Ecol. Appl. 1991, 1, 157–167. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Escalona, J.M.; Sampol, B.; Medrano, H. Effects of drought on photosynthesis in grapevines under field conditions: An evaluation of stomatal and mesophyll limitations. Funct. Plant Biol. 2002, 29, 461–471. [Google Scholar] [CrossRef]
- Aber, J.D.; Federer, C.A. A generalized, lumped-parameter model of photosynthesis, evapotranspiration and net primary production in temperate and boreal forest ecosystems. Oecologia 1992, 92, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ma, L.; Jia, Z.; Jia, L. Root inclusion net method: Novel approach to determine fine root production and turnover in Larix principis-rupprechtii Mayr plantation in North China. Turk. J. Agric. For. 2014, 38, 388–398. [Google Scholar] [CrossRef]
- Zhu, J.; Huang, M.; Chen, Y.; Huang, R.; Li, X. The structure of a culm and shoot producing stand of Phyllostachys pubescens. Chin. J. Plant Ecol. 2000, 24, 483. [Google Scholar]
- Zhang, C.; Ding, X. Analysis and Research on the Factors Affecting the Growth of Phyllostachys edulis Forest. J. Bamboo Res. 1997, 16, 31–36. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Name | Lat (°N) | Lon (°E) | Plant Functional Type |
|---|---|---|---|
| Dinghushan (DHS) | 23.17 | 112.53 | EBF |
| Tianmushan (TMS) | 30.32 | 119.48 | EBF |
| Qianyanzhou (QYZ) | 26.74 | 115.06 | ENF |
| Anji (AJ) | 30.28 | 119.40 | BF |
| Taihuyuan (THY) | 30.26 | 119.59 | BF |
| Symbol | Description | Unit |
|---|---|---|
| Photosynthesis biophysics parameters | ||
| FLNR | Fraction of leaf N in Rubisco | kgNRub·kgNleaf−1 |
| Ko25 | Michaelis constant of oxidation reaction at 25 °C | - |
| Ract25 | Rubisco activity at 25 °C | μmol·mg·Rubisco−1·min−1 |
| Kc25 | Michaelis constant of carboxylation reaction at 25 °C | - |
| Q10kc | The Q10 temperature coefficient of kc | - |
| Q10ko | The Q10 temperature coefficient of ko | - |
| Q10Ract | The Q10 temperature coefficient of Rubisco | - |
| Allocation of carbon parameters | ||
| FRC:LC | New fine root C: leaf C | kgC·kgC−1 |
| LWC:TWC | New live wood C: total wood C | kgC·kgC−1 |
| SC:LC | New stem C: new leaf C | kgC·kgC−1 |
| CRS:SC | New coarse root C: stem C | kgC·kgC−1 |
| CGP | Current growth proportion | - |
| Canopy structure biophysics parameters | ||
| Wint | Water interception coefficient | LAI−1·d−1 |
| k | Light extinction coefficient | - |
| SLA | Average specific leaf area | m·kgC−1 |
| SLAshd:sun | Ratio of shade SLA: sunlit SLA | - |
| LAIall:proj | Ratio of all sides to projected leaf area | - |
| Stomatal conductance biophysics parameters | ||
| Gsmax | Maximum stomatal conductance | m·s−1 |
| Gbl | Boundary layer conductance | m·s−1 |
| LWPf | Leaf water potential: completion of gs reduction | MPa |
| LWPi | Leaf water potential: start of gs reduction | MPa |
| VPDf | Vapor pressure deficit: completion of gs reduction | Pa |
| VPDi | Vapor pressure deficit: start of gs reduction | Pa |
| Gcut | Cuticular conductance | m·s−1 |
| Symbol | Description | Unit |
|---|---|---|
| Heterotrophic respiration biophysics parameters | ||
| C:Nlitter | C: N of falling leaf litter | kgC·kgN−1 |
| LFG | Litterfall period | - |
| LWT | Annual live wood turnover fraction | a−1 |
| Maintenance respiration biophysics parameters | ||
| MRpern | Maintenance respiration in kg C/day per kg of tissue N | kgC·kgN−1·d−1 |
| C:Nleaf | C: N of leaves | kgC·kgN−1 |
| C:Nfr | C: N of fine roots | kgC·kgN−1 |
| C:Ndw | C: N of dead wood | kgC·kgN−1 |
| C:Nlw | C: N of live wood | kgC·kgN−1 |
| Vegetation chemical parameters | ||
| Tt | Transfer growth period | - |
| Llab | Leaf litter labile proportion | - |
| Llig | Leaf litter lignin proportion | - |
| FRlab | Fine root labile proportion | - |
| FRlig | Fine root lignin proportion | - |
| DWlig | Dead wood lignin proportion | - |
| Management measure parameters | ||
| Pdsw | Excavation percentage of winter bamboo shoots | % |
| Pobtr_total | The ratio of hook tip carbon storage to total leaf carbon storage | % |
| SCRages | Selective cutting ratio of each age | - |
| Fer | Apply fertilizer | kgN·hm−2 |
| Pdss | Percentage of bamboo shoots harvested | % |
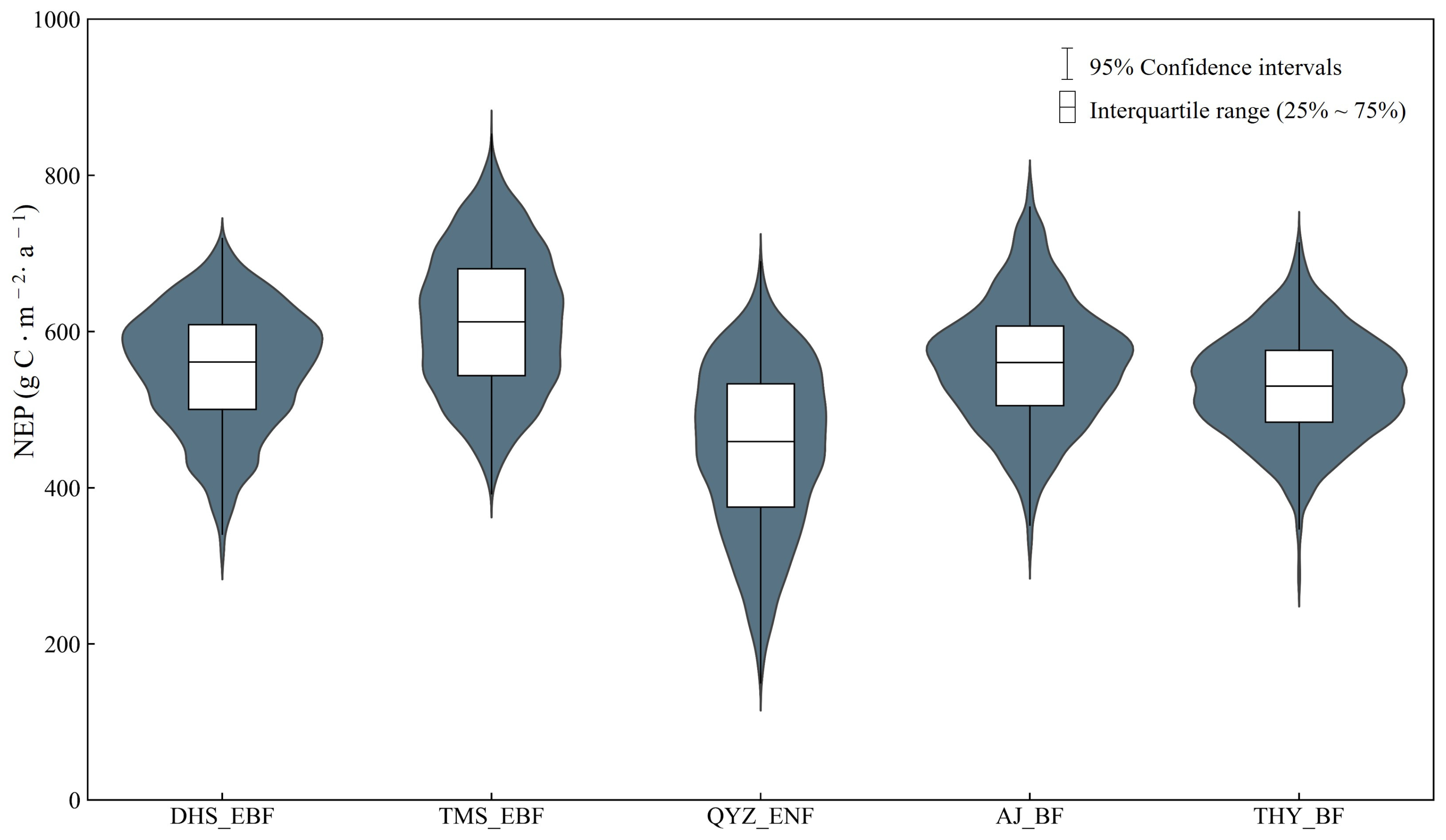
| Statistics | DHS-EBF | TMS-EBF | QYZ-ENF | AJ-BF | THY-BF |
|---|---|---|---|---|---|
| Mean | 551.90 | 612.79 | 449.89 | 558.19 | 529.23 |
| Standard deviation | 76.47 | 89.15 | 104.46 | 79.47 | 67.65 |
| Coefficient of variation (%) | 13.86 | 14.55 | 23.22 | 14.24 | 12.78 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, J.; Mao, F.; Du, H.; Li, X.; Xu, C.; Zheng, Z.; Teng, X.; Ye, F.; Yang, N.; Huang, Z. Improving the Simulation Accuracy of the Net Ecosystem Productivity of Subtropical Forests in China: Sensitivity Analysis and Parameter Calibration Based on the BIOME-BGC Model. Forests 2024, 15, 552. https://doi.org/10.3390/f15030552
Sun J, Mao F, Du H, Li X, Xu C, Zheng Z, Teng X, Ye F, Yang N, Huang Z. Improving the Simulation Accuracy of the Net Ecosystem Productivity of Subtropical Forests in China: Sensitivity Analysis and Parameter Calibration Based on the BIOME-BGC Model. Forests. 2024; 15(3):552. https://doi.org/10.3390/f15030552
Chicago/Turabian StyleSun, Jiaqian, Fangjie Mao, Huaqiang Du, Xuejian Li, Cenheng Xu, Zhaodong Zheng, Xianfeng Teng, Fengfeng Ye, Ningxin Yang, and Zihao Huang. 2024. "Improving the Simulation Accuracy of the Net Ecosystem Productivity of Subtropical Forests in China: Sensitivity Analysis and Parameter Calibration Based on the BIOME-BGC Model" Forests 15, no. 3: 552. https://doi.org/10.3390/f15030552
APA StyleSun, J., Mao, F., Du, H., Li, X., Xu, C., Zheng, Z., Teng, X., Ye, F., Yang, N., & Huang, Z. (2024). Improving the Simulation Accuracy of the Net Ecosystem Productivity of Subtropical Forests in China: Sensitivity Analysis and Parameter Calibration Based on the BIOME-BGC Model. Forests, 15(3), 552. https://doi.org/10.3390/f15030552