
Complex Interplay of Metabolic Pathways in Grafting of Ziziphus Species: Transcriptomic Insights into Regulatory Networks of Carbohydrates and Secondary Metabolite Biosynthesis



 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimentation with Planting Materials and Grafting Techniques
2.2. Determination and Analysis of Phenotypic Indexes of Grafted Seedlings
2.3. Transcriptome Sequencing and Data Analysis
2.4. Gene Validation and Expression Analysis
2.5. Screening of Key Structural Genes in Multiple Pathways
2.6. Construction of Gene Co-Expression Networks via WGCNA
3. Results
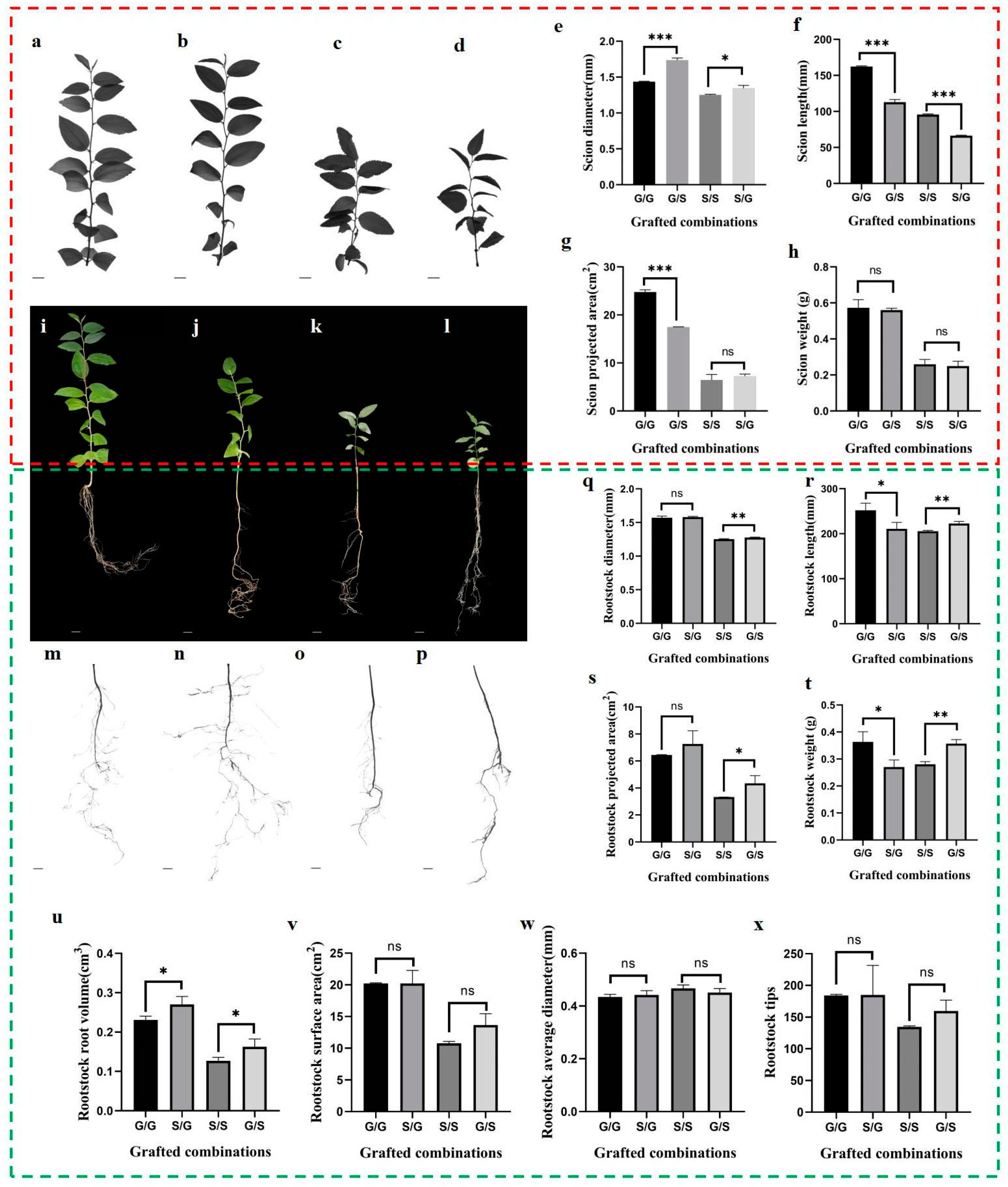
3.1. Phenotypic Variability and Influences across Grafting Combinations
3.2. Transcriptomic Analysis of Grafted Jujube Combinations
3.3. Comprehensive Differential Expression Analysis Illuminates Scion-Rootstock Interactions in Grafted Jujube Plants
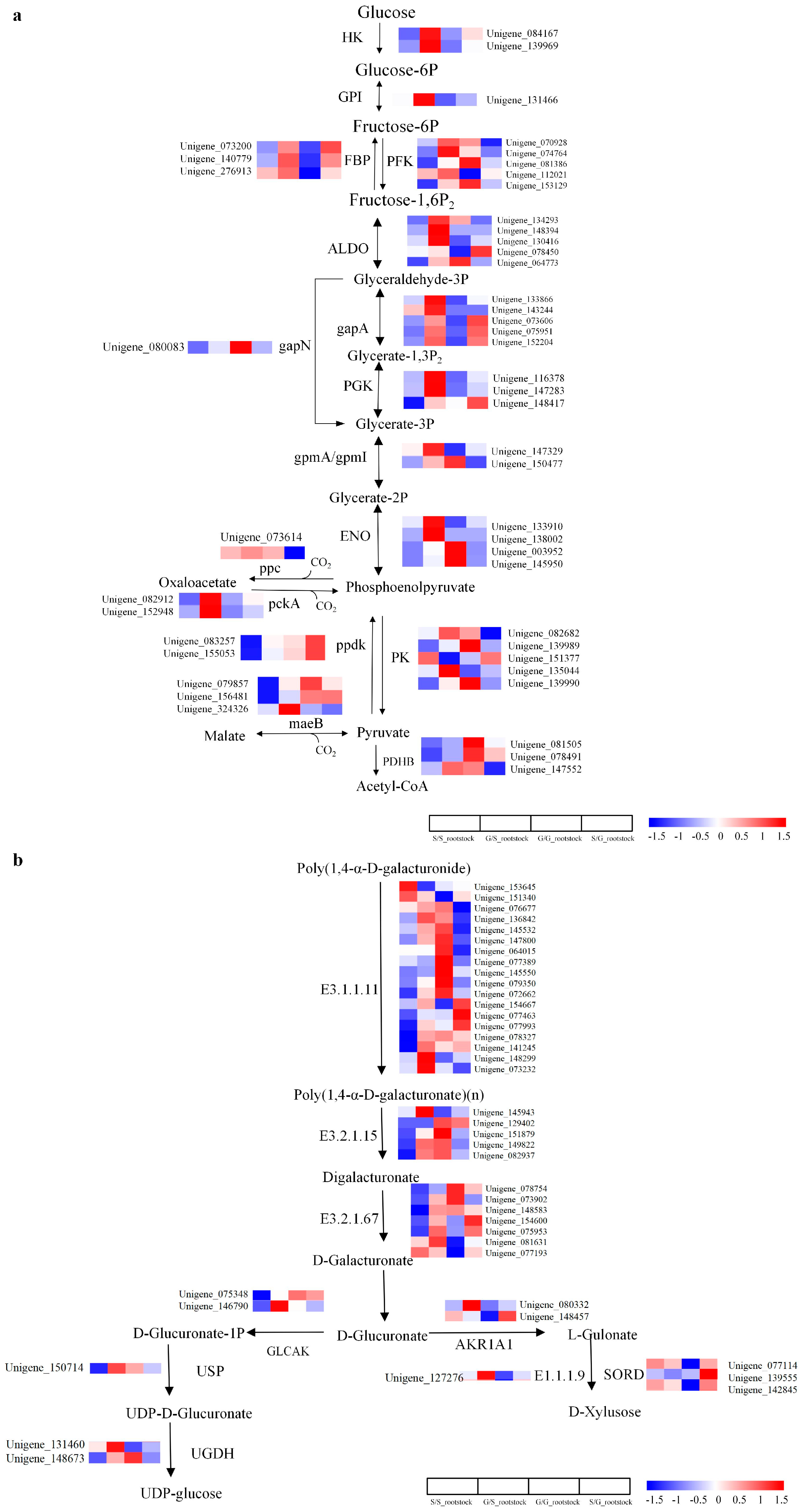
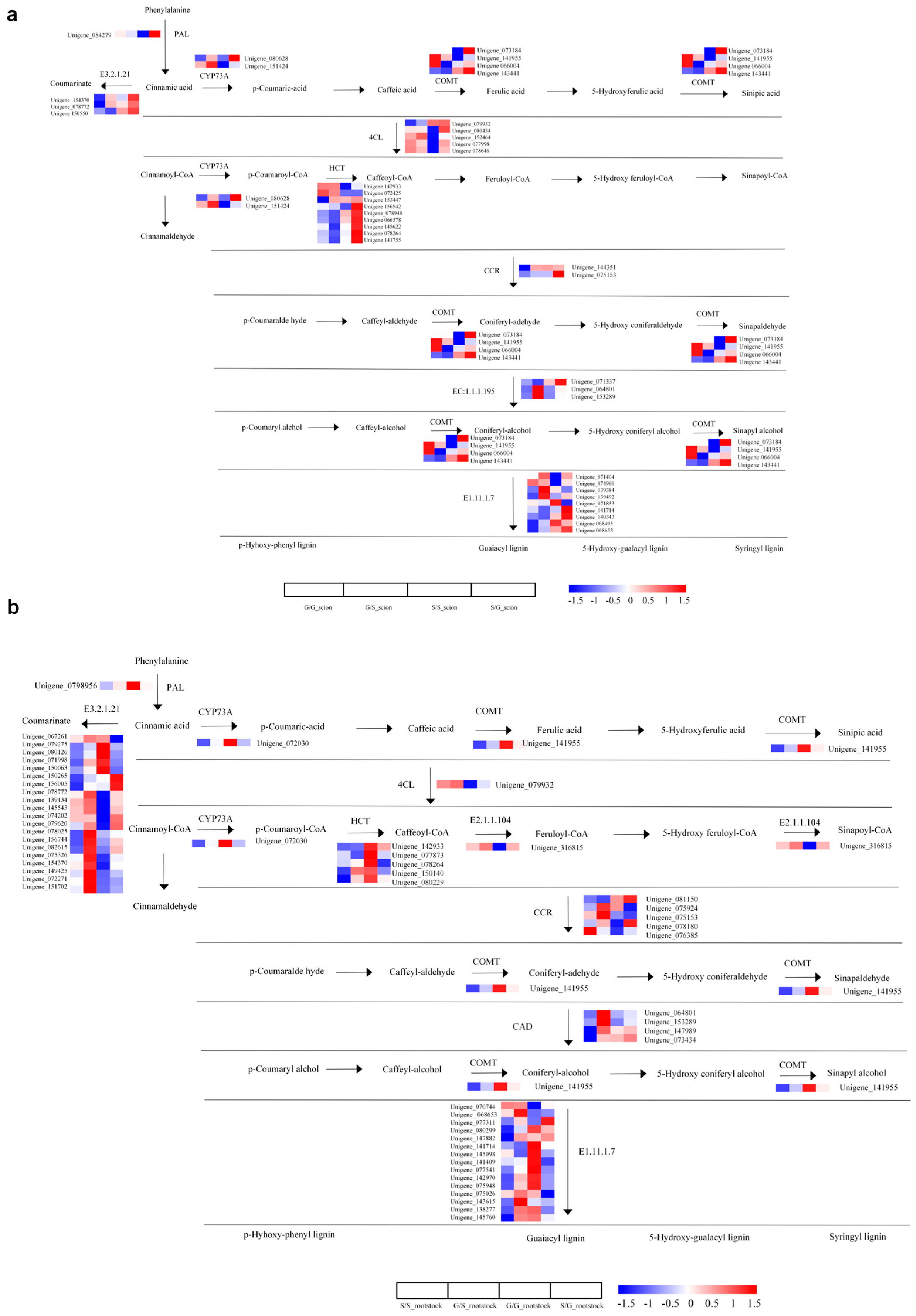
3.4. Multi-Pathway Analysis of Differentially Expressed Genes in Plant Graft Combinations
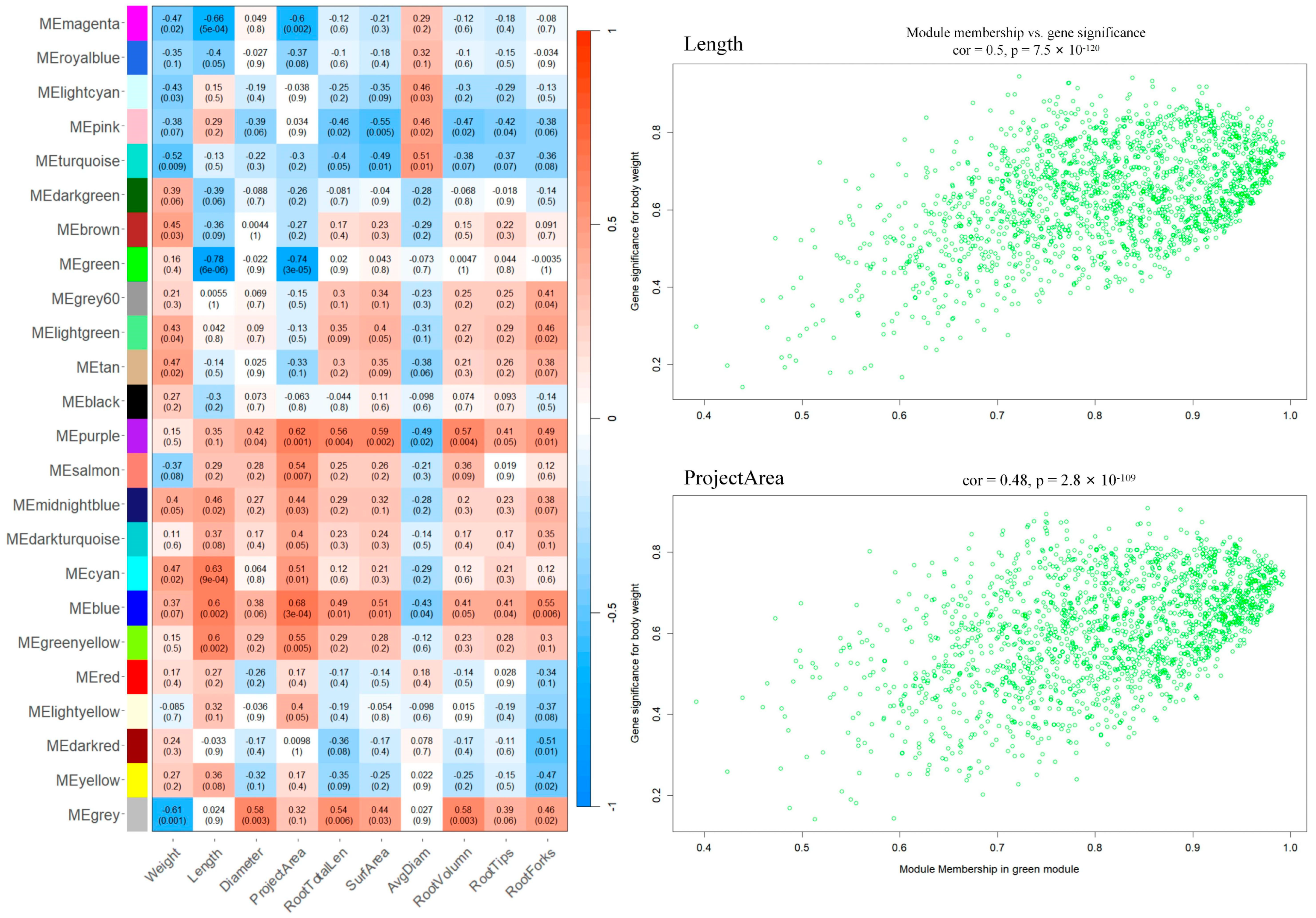
3.5. Exploring Functional Modules and Hub Genes in Post-Grafting Processes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liu, M.J.; Zhao, J.; Cai, Q.L.; Liu, G.C.; Wang, J.R.; Zhao, Z.H.; Liu, P.; Dai, L.; Yan, G.J.; Wang, W.J.; et al. The complex jujube genome provides insights into fruit tree biology. Nat. Commun. 2014, 5, 12. [Google Scholar] [CrossRef]
- Liu, M.-J.; Wang, J.-R.; Liu, P.; Zhao, J.; Zhao, Z.-H.; Dai, L.; Li, X.-S.; Liu, Z.-G. Historical Achievements and Frontier Advances in the Production and Research of Chinese Jujube (Ziziphus jujuba) in China. Acta Hortic. Sin. 2015, 42, 1683–1698. [Google Scholar]
- Lyu, R.H.; Wang, R.; Wu, C.Y.; Bao, Y.J.; Guo, P. Comparative transcriptome analysis of leaves of sour jujube seedlings under salt stress. Acta Physiol. Plant. 2022, 44, 14. [Google Scholar] [CrossRef]
- Hua, Y.; Xu, X.X.; Guo, S.; Xie, H.; Yan, H.; Ma, X.F.; Niu, Y.; Duan, J.A. Wild Jujube (Ziziphus jujuba var. spinosa): A Review of Its Phytonutrients, Health Benefits, Metabolism, and Applications. J. Agric. Food Chem. 2022, 70, 7871–7886. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Han, L.; Zhang, S.; Dai, L.; Li, B.; Han, S.; Zhao, J.; Liu, P.; Zhao, Z.; Liu, M. Insights into the evolution and spatial chromosome architecture of jujube from an updated gapless genome assembly. Plant Commun. 2023, 4, 100662. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wang, J.; Wang, L.; Liu, P.; Zhao, J.; Zhao, Z.; Yao, S.; Stanica, F.; Liu, Z.; Wang, L.; et al. The historical and current research progress on jujube-a superfruit for the future. Hortic. Res. 2020, 7, 119. [Google Scholar] [CrossRef] [PubMed]
- Koepke, T.; Dhingra, A. Rootstock scion somatogenetic interactions in perennial composite plants. Plant Cell Rep. 2013, 32, 1321–1337. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Maryam; Ercisli, S.; Anjum, M.A.; Ahmad, R. Progress in the Methods of Jujube Breeding. Erwerbs-Obstbau 2023, 65, 1217–1225. [Google Scholar] [CrossRef]
- Zhong, H.X.; Liu, Z.J.; Zhang, F.C.; Zhou, X.M.; Sun, X.X.; Li, Y.Y.; Liu, W.W.; Xiao, H.; Wang, N.; Lu, H.; et al. Metabolomic and transcriptomic analyses reveal the effects of self- and hetero-grafting on anthocyanin biosynthesis in grapevine. Hortic. Res. 2022, 9, 11. [Google Scholar] [CrossRef]
- Jang, Y.; Yang, E.; Cho, M.; Um, Y.; Ko, K.; Chun, C. Effect of grafting on growth and incidence of phytophthora blight and bacterial wilt of pepper (Capsicum annuum L.). Hortic. Environ. Biotechnol. 2012, 53, 9–19. [Google Scholar] [CrossRef]
- Zhou, Z.; Yuan, Y.Q.; Wang, K.T.; Wang, H.J.; Huang, J.Q.; Yu, H.; Cui, X. Rootstock-scion interactions affect fruit flavor in grafted tomato. Hortic. Plant J. 2022, 8, 499–510. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, P.; Zhu, W.Y.; Wang, F. De novo Comparative Transcriptome Analysis of Genes Differentially Expressed in the Scion of Homografted and Heterografted Tomato Seedlings. Sci. Rep. 2019, 9, 12. [Google Scholar] [CrossRef]
- Rasool, A.; Mansoor, S.; Bhat, K.M.; Hassan, G.I.; Baba, T.R.; Alyemeni, M.N.; Alsahli, A.A.; El-Serehy, H.A.; Paray, B.A.; Ahmad, P. Mechanisms Underlying Graft Union Formation and Rootstock Scion Interaction in Horticultural Plants. Front. Plant Sci. 2020, 11, 590847. [Google Scholar] [CrossRef]
- Tedesco, S.; Erban, A.; Gupta, S.; Kopka, J.; Fevereiro, P.; Kragler, F.; Pina, A.J.M. The Impact of Metabolic Scion-Rootstock Interactions in Different Grapevine Tissues and Phloem Exudates. Metabolites 2021, 11, 349. [Google Scholar] [CrossRef]
- Tandonnet, J.P.; Cookson, S.; Vivin, P.; Ollat, N. Scion genotype controls biomass allocation and root development in grafted grapevine. Aust. J. Grape Wine Res. 2010, 16, 290–300. [Google Scholar] [CrossRef]
- Zhang, F.C.; Zhong, H.X.; Zhou, X.M.; Pan, M.Q.; Xu, J.; Liu, M.B.; Wang, M.; Liu, G.T.; Xu, T.F.; Wang, Y.J.; et al. Grafting with rootstocks promotes phenolic compound accumulation in grape berry skin during development based on integrative multi-omics analysis. Hortic. Res. 2022, 9, uhac055. [Google Scholar] [CrossRef]
- Yin, W.C.; Wang, X.H.; Liu, H.; Wang, Y.; van Nocker, S.; Tu, M.X.; Fang, J.H.; Guo, J.Q.; Li, Z.; Wang, X.P. Overexpression of VqWRKY31 enhances powdery mildew resistance in grapevine by promoting salicylic acid signaling and specific metabolite synthesis. Hortic. Res. 2022, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Dong, D.; Shi, Y.N.; Mou, Z.M.; Chen, S.Y.; Zhao, D.K. Grafting: A potential method to reveal the differential accumulation mechanism of secondary metabolites. Hortic. Res. 2022, 9, 9. [Google Scholar] [CrossRef]
- Habibi, F.; Liu, T.; Folta, K.; Sarkhosh, A. Physiological, biochemical, and molecular aspects of grafting in fruit trees. Hortic. Res. 2022, 9, 18. [Google Scholar] [CrossRef]
- Jiao, S.Z.; Zeng, F.W.; Huang, Y.P.; Zhang, L.B.; Mao, J.; Chen, B.H. Physiological, biochemical and molecular responses associated with drought tolerance in grafted grapevine. BMC Plant Biol. 2023, 23, 18. [Google Scholar] [CrossRef]
- Li, Q.S.; Gao, Y.; Wang, K.; Feng, J.R.; Sun, S.M.; Lu, X.; Liu, Z.; Zhao, D.Y.; Li, L.W.; Wang, D.J. Transcriptome Analysis of the Effects of Grafting Interstocks on Apple Rootstocks and Scions. Int. J. Mol. Sci. 2023, 24, 807. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Machin, F.; Wang, S.F.; Saplaoura, E.; Kragler, F. Heritable transgene-free genome editing in plants by grafting of wild-type shoots to transgenic donor rootstocks. Nat. Biotechnol. 2023, 41, 958–967. [Google Scholar] [CrossRef] [PubMed]
- Thieme, C.J.; Rojas-Triana, M.; Stecyk, E.; Schudoma, C.; Zhang, W.N.; Yang, L.; Minambres, M.; Walther, D.; Schulze, W.X.; Paz-Ares, J.; et al. Endogenous Arabidopsis messenger RNAs transported to distant tissues. Nat. Plants 2015, 1, 7. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, X.T.; Xu, W.Y.; Liu, T.; Cai, C.M.; Chen, L.Y.; Clark, C.B.; Ma, J.X. Unidirectional movement of small RNAs from shoots to roots in interspecific heterografts. Nat. Plants 2021, 7, 19. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.Q.; Xiang, C.G.; Li, X.J.; Wang, T.; Lu, X.H.; Liu, Z.X.; Gao, L.H.; Zhang, W.N. Identification of Long-Distance Transmissible mRNA between Scion and Rootstock in Cucurbit Seedling Heterografts. Int. J. Mol. Sci. 2020, 21, 5253. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.Y.; Luo, H.; Wang, X.L.; Wang, X.M.; Qiu, X.J.; Liu, H.; Zhou, S.S.; Jia, K.H.; Nie, S.; Bao, Y.T.; et al. Chromosome-Scale Genome Assembly for Chinese Sour Jujube and Insights Into Its Genome Evolution and Domestication Signature. Front. Plant Sci. 2021, 12, 773090. [Google Scholar] [CrossRef] [PubMed]
- Saied, A.S.; Gebauer, J.; Hammer, K.; Buerkert, A.J.G.R.; Evolution, C. Ziziphus spina-christi (L.) Willd.: A multipurpose fruit tree. Genet. Resour. Crop Evol. 2008, 55, 929–937. [Google Scholar] [CrossRef]
- Sohail, M.; Saied, A.S.; Gebauer, J.; Buerkert, A. Effect of NaCl Salinity on Growth and Mineral Composition of Ziziphus spina-christi (L.) Willd. J. Agric. Rural Dev. Trop. Subtrop. 2009, 110, 107–114. [Google Scholar]
- Gorai, M.; Ben Romdhane, R.; Maraghni, M.; Neffati, M. Relationship between leaf gas-exchange characteristics and the performance of Ziziphus spina-christi (L.) Willd. seedlings subjected to salt stress. Photosynthetica 2019, 57, 897–903. [Google Scholar] [CrossRef]
- Wiehle, M.; Prinz, K.; Kehlenbeck, K.; Goenster, S.; Mohamed, S.A.; Buerkert, A.; Gebauer, J. The role of homegardens and forest ecosystems for domestication and conservation of Ziziphus spina-christi (L.) Willd. in the Nuba Mountains, Sudan. Genet. Resour. Crop. Evol. 2014, 61, 1491–1506. [Google Scholar] [CrossRef]
- Pawlowska, A.M.; Camangi, F.; Bader, A.; Braca, A. Flavonoids of Zizyphus jujuba L. and Zizyphus spina-christi (L.) Willd (Rhamnaceae) fruits. Food Chem. 2009, 112, 858–862. [Google Scholar] [CrossRef]
- Singh, A.; Kumar, A.; Kumar, R.; Sheoran, P.; Yadav, R.K.; Sharma, P.C. Multivariate analyses discern shared and contrasting eco-physiological responses to salinity stress of Ziziphus rootstocks and budded trees. S. Afr. J. Bot. 2022, 146, 573–584. [Google Scholar] [CrossRef]
- Zandiehvakili, G.; Khadivi, A. Identification of the promising Ziziphus spina-christi (L.) Willd.genotypes using pomological and chemical proprieties. Food Sci. Nutr. 2021, 9, 5698–5711. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, L.B.; Wu, R.L. Plant grafting: How genetic exchange promotes vascular reconnection. New Phytol. 2017, 214, 56–65. [Google Scholar] [CrossRef]
- Belmonte-Urena, L.J.; Garrido-Cardenas, J.A.; Camacho-Ferre, F. Analysis of World Research on Grafting in Horticultural Plants. Hortscience 2020, 55, 112–120. [Google Scholar] [CrossRef]
- Pal, M.D.; Mitre, I.; Asanica, A.C.; Sestras, A.F.; Peticila, A.G.; Mitre, V. The Influence of Rootstock on the Growth and Fructification of Cherry Cultivars in a High Density Cultivation System. Not. Bot. Hortic. Agrobot. Cluj-Napoca 2017, 45, 451–457. [Google Scholar] [CrossRef]
- Gautier, A.T.; Chambaud, C.; Brocard, L.; Ollat, N.; Gambetta, G.A.; Delrot, S.; Cookson, S.J. Merging genotypes: Graft union formation and scion-rootstock interactions. J. Exp. Bot. 2019, 70, 747–755. [Google Scholar] [CrossRef]
- Banerjee, A.K.; Chatterjee, M.; Yu, Y.; Suh, S.-G.; Miller, W.A.; Hannapel, D.J. Dynamics of a mobile RNA of potato involved in a long-distance signaling pathway. Plant Cell 2006, 18, 3443–3457. [Google Scholar] [CrossRef]
- Warschefsky, E.J.; Klein, L.L.; Frank, M.H.; Chitwood, D.H.; Londo, J.P.; von Wettberg, E.J.B.; Miller, A.J. Rootstocks: Diversity, Domestication, and Impacts on Shoot Phenotypes. Trends Plant Sci. 2016, 21, 418–437. [Google Scholar] [CrossRef]
- Ferlito, F.; Distefano, G.; Gentile, A.; Allegra, M.; Lakso, A.N.; Nicolosi, E. Scion-rootstock interactions influence the growth and behaviour of the grapevine root system in a heavy clay soil. Aust. J. Grape Wine Res. 2020, 26, 68–78. [Google Scholar] [CrossRef]
- Dahniya, M.; Oputa, C.; Hahn, S. Investigating source-sink relations in cassava by reciprocal grafts. Exp. Agric. 1982, 18, 399–402. [Google Scholar] [CrossRef]
- Mudge, K.; Janick, J.; Scofield, S.; Goldschmidt, E.E. A History of Grafting; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2009; Volume 35. [Google Scholar]
- Ahit, O.P.; Abit, S.E.; Posas, M.B. Growth and development of cassava under the traditional and the Mukibat systems of planting [in the Philippines]. Ann. Trop. Res. 1981, 21, 187–198. [Google Scholar]
- Abelenda, J.A.; Navarro, C.; Prat, S. From the model to the crop: Genes controlling tuber formation in potato. Curr. Opin. Biotechnol. 2011, 22, 287–292. [Google Scholar] [CrossRef]
- Li, G.; Ma, J.; Tan, M.; Mao, J.; An, N.; Sha, G.; Zhang, D.; Zhao, C.; Han, M. Transcriptome analysis reveals the effects of sugar metabolism and auxin and cytokinin signaling pathways on root growth and development of grafted apple. BMC Genom. 2016, 17, 150. [Google Scholar] [CrossRef]
- Liao, H.-Y.; Wang, S.; Zhou, C.-Y. Effects of Intergeneric Grafting of Schisandraceae on Root Morphology, Anatomy and Physiology of Rootstocks. Forests 2023, 14, 1183. [Google Scholar] [CrossRef]
- Sun, X.; Tian, L.; Xu, W.; Feng, L.; Jia, W.; Liu, Y.; Chen, Z.; Zhang, S.; Zhang, X.; Ru, G. Integrative Analysis of the Identified Transcriptome and Proteome Major Metabolism Pathways Involved in the Development of Grafted Apricot Hybrids. Forests 2023, 14, 417. [Google Scholar] [CrossRef]
- Amiri, M.E.; Fallahi, E. Influence of Rootstocks on Scion Growth and Leaf Mineral Content of Apple (Malus pumila ‘Golden Delicious’ and ‘Red Gala’) Trees. Acta Hortic 2012, 932, 433–440. [Google Scholar] [CrossRef]
- Clearwater, M.J.; Lowe, R.G.; Hofstee, B.J.; Barclay, C.; Mandemaker, A.J.; Blattmann, P. Hydraulic conductance and rootstock effects in grafted vines of kiwifruit. J. Exp. Bot. 2004, 55, 1371–1382. [Google Scholar] [CrossRef]
- Deng, W.W.; Han, J.Y.; Fan, Y.B.; Tai, Y.L.; Zhu, B.Y.; Lu, M.Q.; Wang, R.J.; Wan, X.C.; Zhang, Z.Z. Uncovering tea-specific secondary metabolism using transcriptomic and metabolomic analyses in grafts of Camellia sinensis and C. oleifera. Tree Genet. Genomes 2018, 14, 16. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Rouphael, Y.; Colla, G.; Zrenner, R.; Schwarz, D. Vegetable Grafting: The Implications of a Growing Agronomic Imperative for Vegetable Fruit Quality and Nutritive Value. Front. Plant Sci. 2017, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Shitan, N.; Dalmas, F.; Dan, K.; Kato, N.; Ueda, K.; Sato, F.; Forestier, C.; Yazaki, K. Characterization of Coptis japonica CjABCB2, an ATP-binding cassette protein involved in alkaloid transport. Phytochemistry 2013, 91, 109–116. [Google Scholar] [CrossRef]
- Xu, D.B.; Yuan, H.W.; Tong, Y.F.; Zhao, L.; Qiu, L.L.; Guo, W.B.; Shen, C.J.; Liu, H.J.; Yan, D.L.; Zheng, B.S. Comparative Proteomic Analysis of the Graft Unions in Hickory (Carya cathayensis) Provides Insights into Response Mechanisms to Grafting Process. Front. Plant Sci. 2017, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Migicovsky, Z.; Harris, Z.N.; Klein, L.L.; Li, M.; McDermaid, A.; Chitwood, D.H.; Fennell, A.; Kovacs, L.G.; Kwasniewski, M.; Londo, J.P.; et al. Rootstock effects on scion phenotypes in a ‘Chambourcin’ experimental vineyard. Hortic. Res. 2019, 6, 13. [Google Scholar] [CrossRef]
- Lu, J.; Cheng, F.; Huang, Y.; Bie, Z. Grafting Watermelon Onto Pumpkin Increases Chilling Tolerance by Up Regulating Arginine Decarboxylase to Increase Putrescine Biosynthesis. Front. Plant Sci. 2021, 12, 812396. [Google Scholar] [CrossRef]
- Liu, W.; Wang, Q.; Zhang, R.; Liu, M.; Wang, C.; Liu, Z.; Xiang, C.; Lu, X.; Zhang, X.; Li, X.; et al. Rootstock-scion exchanging mRNAs participate in the pathways of amino acids and fatty acid metabolism in cucumber under early chilling stress. Hortic. Res. 2022, 9, uhac031. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhan, R.; Wang, L.; Li, J.; Ma, B.; Ma, F.; Li, M. Overexpression of MdFRK2 enhances apple drought resistance by promoting carbohydrate metabolism and root growth under drought stress. Hortic. Plant J. 2023, 9, 884–897. [Google Scholar] [CrossRef]
- Zhang, Z.; Cao, B.; Li, N.; Chen, Z.; Xu, K. Comparative transcriptome analysis of the regulation of ABA signaling genes in different rootstock grafted tomato seedlings under drought stress. Environ. Exp. Bot. 2019, 166, 103814. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, L.; Wang, Z.; Zhang, R.; Liu, P.; Liu, M.; Liu, Z.; Zhao, Z.; Wang, L.; Chen, X.; et al. The regulation of cell wall lignification and lignin biosynthesis during pigmentation of winter jujube. Hortic. Res. 2021, 8, 238. [Google Scholar] [CrossRef]
- Liu, N.; Yang, J.; Fu, X.; Zhang, L.; Tang, K.; Guy, K.M.; Hu, Z.; Guo, S.; Xu, Y.; Zhang, M. Genome-wide identification and comparative analysis of grafting-responsive mRNA in watermelon grafted onto bottle gourd and squash rootstocks by high-throughput sequencing. Mol. Genet. Genom. MGG 2016, 291, 621–633. [Google Scholar] [CrossRef] [PubMed]
- Asahina, M.; Satoh, S. Molecular and physiological mechanisms regulating tissue reunion in incised plant tissues. J. Plant Res. 2015, 128, 381–388. [Google Scholar] [CrossRef]
- Harada, T. Grafting and RNA transport via phloem tissue in horticultural plants. Sci. Hortic. 2010, 125, 545–550. [Google Scholar] [CrossRef]
- Cookson, S.J.; Clemente Moreno, M.J.; Hevin, C.; Nyamba Mendome, L.Z.; Delrot, S.; Trossat-Magnin, C.; Ollat, N. Graft union formation in grapevine induces transcriptional changes related to cell wall modification, wounding, hormone signalling, and secondary metabolism. J. Exp. Bot. 2013, 64, 2997–3008. [Google Scholar] [CrossRef] [PubMed]
- Melnyk, C.W.; Schuster, C.; Leyser, O.; Meyerowitz, E.M. A Developmental Framework for Graft Formation and Vascular Reconnection in Arabidopsis thaliana. Curr. Biol. CB 2015, 25, 1306–1318. [Google Scholar] [CrossRef]
- Melnyk, C.W.; Gabel, A.; Hardcastle, T.J.; Robinson, S.; Miyashima, S.; Grosse, I.; Meyerowitz, E.M. Transcriptome dynamics at Arabidopsis graft junctions reveal an intertissue recognition mechanism that activates vascular regeneration. Proc. Natl. Acad. Sci. USA. 2018, 115, E2447–E2456. [Google Scholar] [CrossRef]
- Vibhuti, M.; Kumar, A.; Sheoran, N.; Nadakkakath, A.V.; Eapen, S.J. Molecular Basis of Endophytic Bacillus megaterium-induced Growth Promotion in Arabidopsis thaliana: Revelation by Microarray-based Gene Expression Analysis. J. Plant Growth Regul. 2017, 36, 118–130. [Google Scholar] [CrossRef]
- Martinoia, E.; Klein, M.; Geisler, M.; Bovet, L.; Forestier, C.; Kolukisaoglu, U.; Muller-Rober, B.; Schulz, B. Multifunctionality of plant ABC transporters—More than just detoxifiers. Planta 2002, 214, 345–355. [Google Scholar] [CrossRef]
- Hwang, J.U.; Song, W.Y.; Hong, D.; Ko, D.; Yamaoka, Y.; Jang, S.; Yim, S.; Lee, E.; Khare, D.; Kim, K.; et al. Plant ABC Transporters Enable Many Unique Aspects of a Terrestrial Plant’s Lifestyle. Mol. Plant. 2016, 9, 338–355. [Google Scholar] [CrossRef]
- Lefevre, F.; Boutry, M. Towards Identification of the Substrates of ATP-Binding Cassette Transporters. Plant Physiol. 2018, 178, 18–39. [Google Scholar] [CrossRef]
- Barzana, G.; Rios, J.J.; Lopez-Zaplana, A.; Nicolas-Espinosa, J.; Yepes-Molina, L.; Garcia-Ibanez, P.; Carvajal, M. Interrelations of nutrient and water transporters in plants under abiotic stress. Physiol. Plant. 2021, 171, 595–619. [Google Scholar] [CrossRef]
- Olmstead, M.A.; Lang, N.S.; Ewers, F.W.; Owens, S.A. Xylem vessel anatomy of sweet cherries grafted onto dwarfing and nondwarfing rootstocks. J. Am. Soc. Hortic. Sci. 2006, 131, 577–585. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
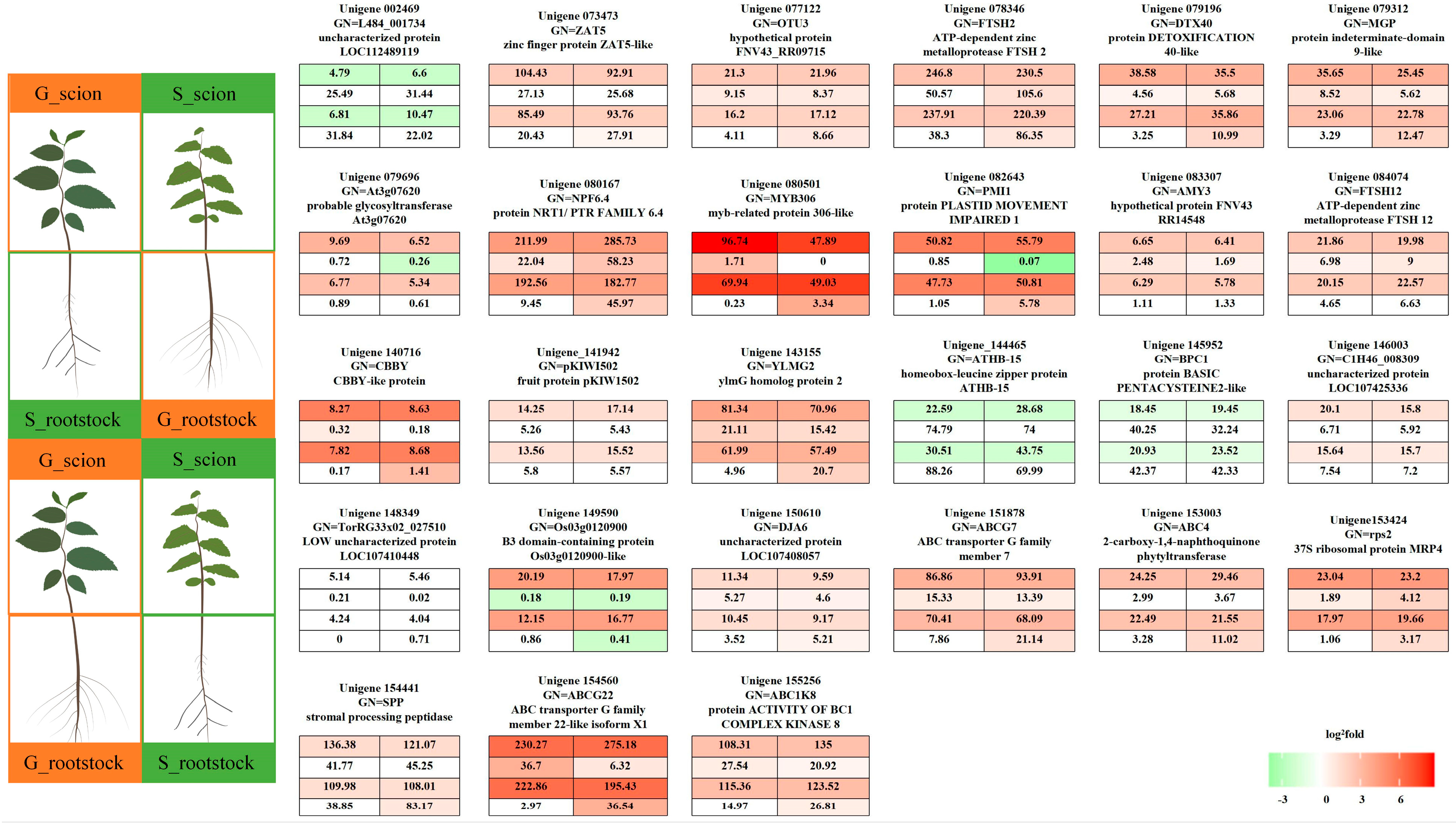{kind=link}
| Treatment | Scion | Rootstock | Grafted Combination |
|---|---|---|---|
| 1 | S | G | S/G |
| 2 | G | S | G/S |
| 3 | S | S | S/S |
| 4 | G | G | G/G |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Sheng, S.; Peng, J.; Liu, Z.; Shao, F.; Wang, S. Complex Interplay of Metabolic Pathways in Grafting of Ziziphus Species: Transcriptomic Insights into Regulatory Networks of Carbohydrates and Secondary Metabolite Biosynthesis. Forests 2024, 15, 618. https://doi.org/10.3390/f15040618
Zhang S, Sheng S, Peng J, Liu Z, Shao F, Wang S. Complex Interplay of Metabolic Pathways in Grafting of Ziziphus Species: Transcriptomic Insights into Regulatory Networks of Carbohydrates and Secondary Metabolite Biosynthesis. Forests. 2024; 15(4):618. https://doi.org/10.3390/f15040618
Chicago/Turabian StyleZhang, Saiyang, Song Sheng, Jiqing Peng, Zhiming Liu, Fengxia Shao, and Sen Wang. 2024. "Complex Interplay of Metabolic Pathways in Grafting of Ziziphus Species: Transcriptomic Insights into Regulatory Networks of Carbohydrates and Secondary Metabolite Biosynthesis" Forests 15, no. 4: 618. https://doi.org/10.3390/f15040618
APA StyleZhang, S., Sheng, S., Peng, J., Liu, Z., Shao, F., & Wang, S. (2024). Complex Interplay of Metabolic Pathways in Grafting of Ziziphus Species: Transcriptomic Insights into Regulatory Networks of Carbohydrates and Secondary Metabolite Biosynthesis. Forests, 15(4), 618. https://doi.org/10.3390/f15040618